Характеристика генетического разнообразия пород и кроссов индеек (Meleagris gallopavo), разводимых в России
, разводимых в России")
Автор: Денискова Т.Е., Доцев А.В., Волкова В.В., Фисинин В.И., Кошкина О.А., Шинкаренко Л.А., Лебедев А.С., Харзинова В.Р., Зиновьева Н.А.
Журнал: Сельскохозяйственная биология @agrobiology
Рубрика: Генетика, геномика
Статья в выпуске: 2 т.61, 2026 года.
Бесплатный доступ
Мясо индейки (Meleagris gallopavo) характеризуется постным составом и высоким содержанием белка и приобретает все большую популярность. В индейководстве практикуется получение межпородных и межлинейных кроссов, поскольку кроссы, как правило, имеют более высокие продуктивные показатели по сравнению с породами. С разработкой платформ для высокопроизводительного SNP-генотипирования начался этап поиска геномных вариантов, лежащих в основе формирования мясных и откормочных качеств индеек. До настоящего времени не было создано стандартизированных панелей микросателлитных маркеров для генетической характеристики пород и популяций индеек. В связи с этим продолжается выявление новых и апробация ранее известных локусов у различных пород и кроссах индеек. В настоящей работе мы впервые установили, что современная популяция бронзовой северокавказской породы (2025 год) индеек Meleagris gallopavo имеет более высокие значения генетического и аллельного разнообразия по сравнению с архивной популяцией этой породы в 2011 году, в которой, в свою очередь, были выявлены достоверные значения инбридинга. Отечественные кроссы Виктория и Универсал характеризовались сопоставимыми значениями генетического разнообразия и более высокими значениями аллельного разнообразия по сравнению с иностранными промышленными кроссами. Генетическая дифференциация между исследуемыми породами и кроссами индеек варьировала от незначительной до высокой. Цель работы — оценить генетическое разнообразие и дифференциацию пород и кроссов индеек, разводимых в России, а также изменения популяционно-генетических показателей у индеек бронзовой северокавказской и черной тихорецкой пород в процессе разведения. Работу проводили в 2025 году. Образцы цельной крови индеек (Meleagris gallopavo) бронзовой северокавказской (популяция 2025 года, n= 48) и черной тихорецкой (популяция 2025 года, n= 34) пород были отобраны в Селекционно-генетическом центре Северо-Кавказская зональная опытная станция по птицеводству» — филиал ФНЦ ВНИТИП (Ставропольский край, Георгиевский р-н, с. Обильное). Образцы цельной крови популяций бронзовой северокавказской (популяция 2011 года, n= 13), черной тихорецкой (популяция 2011 года, n= 15), узбекской палевой (n= 13), белой московской (n= 15), белой широкогрудой (n= 29), серебристой северокавказской (n= 9), белой северокавказской (n= 14), а также кроссов B.U.T.6 (n= 12), Big-6 (n= 20), Big -10 (n= 18), Виктория (n= 14) и Универсал (n= 9) были получены из биоколлекции Банк генетического материала домашних и диких видов животных и птицы ФГБНУ ФИЦ ВИЖ им. академика Л.К. Эрнста (УНУ № 498808). Анализ по шести микросателлитам MNT036, MNT0115, MNT0247, MNT0348, MNT0370 и WT54 выполняли на генетическом анализаторе ABI3130xl («Applied Biosystems, Inc.», США). В R пакете diveRsity были рассчитаны показатели генетического разнообразия и значения генетической дифференциации, а также выполнена оценка степени миграции генов между популяциями. Генетическая сеть была построена в программе SplitsTree 4.14.5. Кластерный анализ проводили в программе Structure 2.3.4. Значения наблюдаемой гетерозиготности варьировали от 0,319 в популяции бронзовой северокавказской породы 2011 года до 0,600 в узбекской палевой породе среди пород и от 0,444 в кроссе B.U.T.6 до 0,620 в кроссе Big-10. Значения аллельного разнообразия варьировали от 3,335 (B.U.T.6) до 4,284 (Универсал) среди кроссов и от 3,000 (серебристая северокавказская) до 4,215 (бронзовая северокавказская популяция 2025 года) среди пород. Популяция черной тихорецкой породы 2025 года характеризовалась более высоким аллельным разнообразием по сравнению с архивной (3,612 и 3,026), но практически не отличалась по наблюдаемой гетерозиготности. Обе популяции имели схожую генетическую структуру и кластеризовались совместно (FST = 0,09, Jost`s D = 0,07). Популяция бронзовой северокавказской породы 2025 года характеризовалась более высоким генетическим (HO 0,507 и 0,319) и аллельным разнообразием (AR 4,215 и 3,290) по сравнению с популяцией 2011 года. Популяции 2011 и 2025 годов различались по генетической структуре и кластеризовались отдельно друг от друга. Это указывает на генетические изменения, произошедшие в бронзовой северокавказской породе, причины которых будут изучены более подробно в наших следующих работах с увеличенным числом микросателлитных локусов и анализом последовательностей полного генома.
Индейка, породы, кроссы, генетическое разнообразие, микросателлиты, полиморфизм
Короткий адрес: https://sciup.org/142247688
IDR: 142247688 | УДК: 636.592:575.174 | DOI: 10.15389/agrobiology.2026.2.273rus
Characterization of genetic diversity of turkey (Meleagris gallopavo) breeds and crosses raised in Russia
Turkey meat (Meleagris gallopavo) is characterized by its lean composition and high protein content that contributes to its growing popularity. In turkey farming, crossbreeding is widely practiced, as crosses generally exhibit superior productivity compared to pure breeds. The development of high-throughput SNP genotyping arrays has enabled the search for genomic variants that influence meat and fattening traits in turkeys. Nevertheless, no standardized panels of microsatellite markers have been created for genetic characterization of turkey breeds and populations. In this regard, the search for novel loci and the validation of previously described loci in various turkey breeds and crosses is ongoing. In this study, we demonstrated for the first time that the contemporary population of the Bronze North Caucasian turkey breed Meleagris gallopavo (2025) exhibited higher genetic and allelic diversity compared to the archival population of this breed (2011), which, in turn, showed significant levels of inbreeding. The domestic Victoria and Universal crosses exhibited comparable levels of genetic diversity, but higher allelic diversity compared to foreign commercial crosses. Genetic differentiation among the studied turkey breeds and crosses ranged from negligible to high. The aim of this study was to assess the genetic diversity and differentiation of turkey breeds and crosses raised in Russia, as well as to analyze changes in population genetic parameters in the Bronze North Caucasian and Black Tikhoretsk turkey populations during the breeding process. The study was conducted in 2025. Whole blood samples were collected from turkeys (Meleagris gallopavo) of the Bronze North Caucasian (2025 population, n = 48) and Black Tikhoretsk (2025 population, n =) breeds at the Selection and Genetic Center North Caucasian Zonal Experimental Station for Poultry Farming — a branch of the Federal Scientific Center VNITIP (Stavropol Territory, Georgievsky District, Obilnoye village). Whole blood samples of the populations of the Bronze North Caucasian (population of 2011, n = 13), Black Tikhoretsk (population of 2011, n = 15), Uzbek fawn (n = 13), White Moscow (n = 15), White Broad-Breasted (n = 29), Silver North Caucasian (n = 9), White North Caucasian (n = 14), as well as the crosses B.U.T.6 (n = 12), Big-6 (n = 20), Big-10 (n = 18), Victoria (n = 14) and Universal (n = 9) were obtained from the biocollection Bank of Genetic Material of Domestic and Wild Species of Animals and Poultry, Ernst Federal Research Center for Animal Husbandry (UNU No. 498808). Six microsatellite markers MNT036, MNT0115, MNT0247, MNT0348, MNT0370, and WT54 were genotyped using an ABI3130xl Genetic Analyzer (Applied Biosystems, USA). Genetic diversity indices, genetic differentiation metrics, and gene flow between populations were calculated using the R package diversity. The genetic network was constructed using SplitsTree 4.14.5. Cluster analysis was performed using Structure 2.3.4. The observed heterozygosity values ranged from 0.319 in the 2011 Bronze North Caucasian population to 0.600 in the Uzbek Fawn breed among breeds and from 0.444 in the B.U.T.6 cross to 0.620 in the Big-10 cross. The allelic diversity values varied from 3.335 (B.U.T.6) to 4.284 (Universal) among crosses and from 3.000 (Silver North Caucasian) to 4.215 (Bronze North Caucasian population of 2025) among breeds. The 2025 Black Tikhoretsk population exhibited higher allelic diversity compared to the archived population (3.612 vs 3.026), but similar observed heterozygosity. Both populations had a similar genetic structure and clustered together (Fst = 0.09, Jost's D = 0.07). Compared to the 2011 population, the 2025 North Caucasian Bronze group showed higher genetic diversity (HO = 0.507 vs 0.319) and allelic richness (AR = 4.215 vs 3.290). The2011 and 2025 populations differed genetically, forming separate clusters. This indicates that genetic changes have occurred in the North Caucasian Bronze breed. The underlying causes will be investigated in future studies using an expanded panel of microsatellite loci and whole-genome sequencing.
Текст научной статьи Характеристика генетического разнообразия пород и кроссов индеек (Meleagris gallopavo), разводимых в России
В мире отмечается устойчивый рост спроса на птицеводческую продукцию, в частности на мясо индейки ( Meleagris gallopavo ), которое характеризуется постным составом и высоким содержанием белка (1). Мясо индейки реализуется в виде целых тушек и разделанных сырых частей, а также переработанных продуктов — различных мясных деликатесов (1).
В истории одомашнивания индейки в Новом Свете до сих пор имеется большое число пробелов. Так, на основе анализа археологической митохондриальной ДНК C.F. Speller с соавт. (2) указали на возможность существования двух отдельных процессов одомашнивания — в северной или Центральной Мексике и на юго-западе Северной Америки. Эти предположения согласуются с анализом нуклеотидного разнообразия митогеномов современных представителей домашней индейки из различных географических популяций (3). Кроме того, E.K. Thornton с соавт. (4) обнаружили археологические доказательства существования мексиканской индейки у древних майя (300 год до н.э.-100 год н.э.).
Известно, что все современные породы домашней индейки — это потомки индейки из Центральной Мексики, завезенной в Европу во время испанской колонизации юга США и Мексики в XVI веке (2). В России для производства мяса индейки используются как отечественные породы и кроссы, так и кроссы иностранного происхождения.
Первыми породами индеек, созданными в СССР, были бронзовая северокавказская и белая северокавказская (5). Бронзовая северокавказская порода (официально зарегистрирована в 1956 году) была получена посредством скрещивания аборигенных индеек с индюками бронзовой и бронзовой широкогрудой пород в 1950-1960-х годах. Порода характеризуется высокой жизнеспособностью и отличной приспособленностью к пастбищному разведению (5, 6). Белая северокавказская порода была выведена в результате улучшения аборигенных бронзовых северокавказских индеек самцами белой широкогрудой породы английского происхождения в 1970-1980-е годы и отличается скороспелостью, высокой яичной продуктивностью и хорошей приспособленностью к разведению в условиях юга России (5, 6). Черная тихорецкая порода (ранее известная как черная кубанская, черная краснодарская) была получена при длительной селекции местных черных индеек в Тихорецком районе Краснодарского края. Индейки этой породы отличаются активностью, приспособляемостью к условиям содержания и хорошими мясными качествами (6). На первом этапе создания белой московской породы использовались местные белые и белтсвилские индейки, после чего был привлечен генетический материал белых голландских индеек (5). В целом, порода характеризуется относительно высокой плодовитостью и выводимостью яиц, хорошим выходом мяса (6). В 2008 году официально утверждена серебристая северокавказская порода, созданная посредством улучшения узбекских палевых индеек самцами белой широкогрудой породы с целью совершенствования репродуктивных качеств и мясной продуктивности (5, 7). Узбекская палевая порода была выведена на основе узбекских бронзовых индеек, улучшенных белой широкогрудой породой (5). Белая широкогрудая порода — это трансграничная порода, выведенная на основе голландских белых индеек в США в 1960-х годах (5).
В индейководстве практикуется получение межпородных и межлинейных кроссов, поскольку кроссы, как правило, характеризуются более высокими продуктивными показателями по сравнению с породами. Универсал и Виктория — это два известных отечественных кросса. Кросс
Универсал, утвержденный в 2004 году, был создан в процессе длительной селекционной работы с использованием разных подходов (вводное скрещивание, разведение «в себе» при индивидуально-гнездовой и групповой селекции, искусственное осеменение и естественное спаривание) и с привлечением генетического материала белой широкогрудой (линии К, А-7 и О2) и белой северокавказской пород (линии Б1 и Б6). Кросс включает материнскую линию У1 и отцовскую линию У2. В генеалогической структуре линии У1 выделяют микролинии, лучшие по яйценоскости, выводимости, сохранности, плодовитости и мясным качествам, линии У2 — микролинии, лучшие по живой массе в убойном возрасте, обмускуленности груди, сохранности молодняка, качеству спермопродукции (8, 9). Кросс Виктория был создан на основе линий О2 и О4 белой широкогрудой породы и состоит из двух линий: материнская линия КА связана с высокой яйценоскостью и отцовская линия ВИ ответственна за наследование мясной и откормочной продуктивности (10). Следует отметить, что ведутся селекционные работы по созданию кросса индеек тяжелого типа на основе линий К1 (новая линия), РЛ11 (резервная линия 11), РЛ22 (резервная линия 22) и 1124 (гетерогенная популяция) (11). В промышленном птицеводстве популярны тяжелые кроссы индеек, например B.U.T.6 («Aviagen Turkeys, Ltd.», Великобритания).
С разработкой платформ для высокопроизводительного SNP-генотипирования начался этап поиска геномных вариантов, лежащих в основе формирования мясных и откормочных качеств индеек. Например, были выявлены 37 генов, ассоциированные с ростом и метаболизмом (12), и 14 функциональных кандидатов, связанных с белковым обменом и иммунитетом (13). Полногеномный поиск ассоциаций позволил идентифицировать 95 позиционных генов-кандидатов, влияющих на качество мяса, в частности появление аномальных миопатий, у кроссов белых индеек (14).
Создание и массовое использование кроссов, помимо положительных аспектов в виде повышенной продуктивности, имеет и негативные последствия. Прежде всего, утрачивается генетическое разнообразие многих локальных пород, которые не могут конкурировать с кроссами в рамках промышленного производства, что ставит их под угрозу деградации и исчезновения (15). В связи с этим, несмотря на внедрение новых технологий (ДНК-чипы и NGS секвенирование), микросателлиты по-прежнему остаются полезным типом ДНК-маркеров для проведения популяционно-генетических исследований сельскохозяйственных животных и птицы (16). Их использование дает возможность быстро оценить процессы, происходящие в алле-лофонде местных пород (17, 18).
До настоящего времени не было создано стандартизированных панелей микросателлитных маркеров для генетической характеристики пород и популяций индеек (15). В связи с этим продолжается открытие новых и апробация ранее известных локусов в различных породах и кроссах индеек.
A.M. Canales Vergara с соавт. (15) протестировали 39 локусов для изучения генетической изменчивости местных пород индеек из Мексики, США, Италии, Бразилии, Египта и Испании. Показано, что микросателлиты были полиморфными и имели число аллелей от 4 до 19 со средним числом аллелей на маркер 10,28.
Охарактеризовано генетическое разнообразие 10 популяций домашних индеек из Европы, Америки и с Ближнего Востока с использованием 34 микросателлитов. Установлено, что мексиканские популяции индеек наиболее разнообразны. Испанские популяции также характеризовались довольно высоким генетическим разнообразием, поскольку Испания импортировала большое число индеек из Нового Света (19).
Создание мультиплексных панелей микросателлитных маркеров требует предварительного тестирования локусов с целью определения степени их полиморфизма в популяциях локальных пород индеек. Так, предыдущие генетические исследования отечественных пород индеек выявили низкую степень полиморфизма (1-1,83 аллелей на локус) (5, 20). В связи с этим актуален отбор оптимальных микросателлитных локусов для создания панелей для генетического мониторинга индеек, разводимых в нашей стране.
В настоящей работе мы впервые установили, что современная популяция бронзовой северокавказской породы (2025 года) индеек Meleagris gallopavo характеризировалась более высокими значениями генетического и аллельного разнообразия по сравнению с архивной популяцией этой породы (2011 года), в которой, в свою очередь, были выявлены достоверные значения инбридинга. Установлено более высокое аллельное разнообразие в популяциях кроссов, созданных в России, по сравнению с кроссами иностранного происхождения. Генетическая дифференциация между исследуемыми породами и кроссами индеек варьировала от незначительной до высокой.
Цель работы — оценить генетическое разнообразие и дифференциацию пород и кроссов индеек, разводимых в России, а также изменения популяционно-генетических показателей у индеек бронзовой северокавказской и черной тихорецкой пород в процессе разведения.
Ìåòîäèêà. Работу проводили в 2025 году. Образцы цельной крови были отобраны от индеек ( Meleagris gallopavo ) бронзовой северокавказской (популяция 2025 года, n = 48) и черной тихорецкой (популяция 2025 года, n = 34) пород в Селекционно-генетическом центре «Северо-Кавказская зональная опытная станция по птицеводству» — филиал ФНЦ ВНИТИП (Ставропольский край, Георгиевский р-н, с. Обильное). Образцы цельной крови популяций бронзовой северокавказской (популяция 2011 года, n = 13), черной тихорецкой (популяция 2011 года, n = 15), узбекской палевой ( n = 13), белой московской ( n = 15), белой широкогрудой ( n = 29), серебристой северокавказской ( n = 9), белой северокавказской ( n = 14), а также кроссов B.U.T.6 ( n = 12), Big-6 ( n = 20), Big -10 ( n = 18), Виктория ( n = 14) и Универсал ( n = 9) были получены из биоколлекции Банк генетического материала домашних и диких видов животных и птицы ФГБНУ ФИЦ ВИЖ им. Л.К. Эрнста (УНУ ¹ 498808).
ДНК выделяли из цельной крови с помощью наборов ДНК-Экстран-1 (ЗАО «Синтол», Россия).
Выбор микросателлитных локусов для включения в состав модернизированной тест-системы для популяционно-генетических исследований индеек был выполнен на основе анализа источников литературы. На первом этапе были выбраны шесть микросателлитных локусов: MNT036 (21), MNT0115 (22), MNT0247 (23), MNT0348 (24), MNT0370 (23), WT54 (15). Последовательности праймеров также выбирали согласно соответствующим источникам: прямой 3´-TAACTGTCTGCCAGGTGGTG-5´ и обратный 3´-GATCTCGGGTGGTGATTGC-5´ для локуса MNT20, прямой 3´-ACC-AACAGTCAGGTGGTGTG-5´ и обратный 3´-GGAGTATATATATGGTG-GATTTGAGG-5´ для MNT036, прямой 3´-CCTTGCCAGGCTTTTCTTC-5´ и обратный 3´-GCTTGCATAGGCTTCGTAGG-5´ для MNT0115, прямой 3´-GAACAACTTGAGGCCATTCC-5´ и обратный 3´-TTCCAGCCTTCCC-TAATGTG-5´ для MNT0247, прямой 3´-AACACAGTGACCCTGCCAAC-5´ и обратный 3´-TTTTCTTCCCCTCTTTGTTTTC-5´ для MNT0370, прямой 3´-TGGGACCAAAGGTATTTTCC-5´ и обратный 3´-CAAGATAGAGCAC-276
AAACAAGTGC-5´ для MNT0348, прямой 3´-CATCCATCCTGTGTCTCCCA-5´ и обратный 3´-ACAGTGTCACGATTCCCCTT-5´ для WT54. Флуоресцентная метка FAM была выбрана для MNT0247 и MNT0348, флуоресцентная метка R6G — для MNT036, MNT0115, MNT0370, WT54. Праймеры синтезировали в ЗАО «Евроген» (Россия). Анализ микросателлитов выполняли на генетическом анализаторе ABI3130xl («Applied Biosystems, Inc.», США). Размеры аллелей были соотнесены с размерным стандартом с использованием программного обеспечения GeneMapper 4 («Applied Biosystems, Inc.», США).
Показатели генетического разнообразия (средние значения и ошибки среднего) — несмещенная ожидаемая ( u H e ) и наблюдаемая (H o ) гетерозиготность, несмещенный коэффициент инбридинга ( u F is ), аллельное разнообразие, скорректированное на размер выборки (A r ) рассчитывали с использованием R пакета «diveRsity» (24). В этом же пакете были вычислены попарные значения F st (25) и Jost`s D (26), а также выполнена оценка степени миграции генов между популяциями (divMigrate). Анализ divMigrate был проведен с 1000 бутстрепами на основе оценки числа мигрантов на поколение (Nm) (27).
Генетическая сеть была построена по принципу ближайшего соседа (Neighbor Net) в программе SplitsTree 4.14.5 (28). Для установления межпопуляционных связей применялась функция максимального правдоподобия, реализованная в программе Structure 2.3.4 (29) с последующей визуализацией с помощью пакета R pophelper (30).
Результаты. Анализ наблюдаемой гетерозиготности (Н о ) выявил наибольшие значения в узбекской палевой породе (H o = 0,600) и кроссе Big-10 (H o = 0,620), наименьшие — в популяции 2011 года бронзовой северокавказской породы (H o = 0,319) и в кроссе B.U.T.6 (H o = 0,444) (табл. 1).
1. Показатели генетического разнообразия в исследуемых кроссах и породах индеек ( Meleagris gallopavo ) ( M ±SEM, ЦКП «Биоресурсы и биоинженерия сельскохозяйственных животных» ФГБНУ ФИЦ ВИЖ им. Л.К. Эрнста, 2025 год)
|
Группа |
n |
H O |
u H e |
u F is |
A r |
|
Породы |
|||||
|
Бронзовая северокавказская (популяция 2011 года) |
13 |
0,319±0,092 |
0,493±0,099 |
0,331 [0,063; 0,599] |
3,29±0,408 |
|
Бронзовая северокавказская (популяция 2025 года) |
48 |
0,507±0,083 |
0,62±0,088 |
0,143 [ - 0,056; 0,342] |
4,215±0,541 |
|
Черная тихорецкая (популяция 2011 года) |
15 |
0,567±0,075 |
0,523±0,07 |
- 0,101 [ - 0,257; 0,055] |
3,026±0,284 |
|
Черная тихорецкая (популяция 2025 года) |
34 |
0,569±0,068 |
0,605±0,056 |
0,056 [ - 0,137; 0,249] |
3,612±0,425 |
|
Узбекская палевая |
15 |
0,600±0,071 |
0,600±0,047 |
0,011 [ - 0,125; 0,147] |
3,24±0,311 |
|
Серебристая северокавказская |
9 |
0,556±0,091 |
0,523±0,043 |
- 0,045 [ - 0,293; 0,203] |
3±0,258 |
|
Белая широкогрудая |
29 |
0,500±0,11 |
0,593±0,091 |
0,15 [ - 0,067; 0,367] |
3,874±0,476 |
|
Белая северокавказская |
15 |
0,526±0,104 |
0,515±0,104 |
- 0,034 [ - 0,157; 0,089] |
3,374±0,392 |
|
Белая московская |
15 |
0,456±0,09 0,562±0,098 Кроссы |
0,16 [ - 0,04; 0,36] |
3,715±0,473 |
|
|
Виктория |
14 |
0,577±0,12 |
0,597±0,1 |
0,028 [ - 0,163; 0,219] |
4,049±0,521 |
|
Универсал |
9 |
0,577±0,101 |
0,631±0,107 |
0,059 [ - 0,114; 0,232] |
4,284±0,659 |
|
Big-6 |
20 |
0,458±0,044 |
0,44±0,056 |
- 0,07 [ - 0,193; 0,053] |
3,383±0,373 |
|
Big -10 |
18 |
0,620±0,134 |
0,556±0,111 |
- 0,101 [ - 0,204; 0,002] |
3,651±0,504 |
Продолжение таблицы 1
B.U.T.6 12 0,444±0,09 0,420±0,086 - 0,057 3,335±0,424
[ - 0,114; 0]
Примечание. n — число образцов, Н о — наблюдаемая гетерозиготность, uH E — несмещенная ожидаемая гетерозиготность, A r — аллельное разнообразие, u F is — несмещенный коэффициент инбридинга (доверительный интервал 95 %).
Значения коэффициента инбридинга были недостоверны во всех исследуемых группах индеек, кроме популяции бронзовой северокавказской породы 2011 года. Тенденции к избытку гетерозигот наблюдались в серебристой северокавказской и белой северокавказской породах, а также в кроссах Big-6, Big-10 и B.U.T.6. Значения аллельного разнообразия варьировали от 3,335 (B.U.T.6) до 4,284 (Универсал) среди кроссов и от 3 (серебристая северокавказская) до 4,215 (бронзовая северокавказская популяция 2025 года) среди пород.
Наличие представителей популяций 2011 и 2025 годов для двух пород индеек позволило провести анализ изменений в параметрах генетического и аллельного разнообразия в динамике. Популяция 2025 года бронзовой северокавказской породы характеризовалась более высокой наблюдаемой гетерозиготностью: H o = 0,507 по сравнению с H o = 0,319, выявленной в популяции 2011 года. Следует отметить, что только популяция 2011 года бронзовой северокавказской породы среди всех исследуемых групп индеек характеризовалась достоверно значимыми значениями коэффициента инбридинга, а именно недостатком гетерозигот. У представителей этой породы в 2025 году было отмечено большее аллельное разнообразие по сравнению с популяцией 2011 года: 4,215 против 3,290 аллелей. У черной тихорецкой породы не было выявлено различий в наблюдаемой гетерозиготности. Значения коэффициента инбридинга оказались недостоверными в обеих исследуемых группах. Популяция 2025 года характеризовалась более высоким аллельным разнообразием по сравнению с популяцией 2011 года: соответственно 3,612 и 3,026 аллелей.
2. Генетическая дифференциация между исследуемыми кроссами и породами индеек ( Meleagris gallopavo ), оцененная на основе расчета попарных значений Fst и генетических дистанций Jost`s D (ЦКП «Биоресурсы и биоинженерия сельскохозяйственных животных» ФГБНУ ФИЦ ВИЖ им. Л.К. Эрнста, 2025 год)
|
Группа |
1 |
2 |
3 |
4 |
5 |
6 |
7 |
8 |
9 |
10 |
11 |
12 |
13 |
14 |
|
1 |
0,09 |
0,18 |
0,26 |
0,19 |
0,19 |
0,13 |
0,20 |
0,18 |
0,15 |
0,12 |
0,27 |
0,18 |
0,25 |
|
|
2 |
0,07 |
0,10 |
0,14 |
0,05 |
0,11 |
0,06 |
0,15 |
0,08 |
0,08 |
0,08 |
0,16 |
0,09 |
0,13 |
|
|
3 |
0,16 |
0,08 |
0,01 |
0,11 |
0,15 |
0,16 |
0,24 |
0,20 |
0,17 |
0,19 |
0,18 |
0,16 |
0,17 |
|
|
4 |
0,22 |
0,10 |
0 |
0,10 |
0,19 |
0,21 |
0,29 |
0,23 |
0,22 |
0,24 |
0,28 |
0,24 |
0,26 |
|
|
5 |
0,12 |
0,04 |
0,07 |
0,04 |
0,10 |
0,07 |
0,15 |
0,10 |
0,09 |
0,12 |
0,21 |
0,13 |
0,17 |
|
|
6 |
0,24 |
0,16 |
0,21 |
0,28 |
0,10 |
0,05 |
0,08 |
0,06 |
0,04 |
0,04 |
0,17 |
0,09 |
0,16 |
|
|
7 |
0,15 |
0,03 |
0,16 |
0,25 |
0,06 |
0,07 |
0,02 |
- 0,01 |
- 0,01 |
0,00 |
0,20 |
0,07 |
0,17 |
|
|
8 |
0,23 |
0,11 |
0,21 |
0,31 |
0,12 |
0,09 |
0 |
0,03 |
0,03 |
0,03 |
0,31 |
0,15 |
0,27 |
|
|
9 |
0,19 |
0,05 |
0,26 |
0,30 |
0,08 |
0,09 |
0 |
0,03 |
0,00 |
0 |
0,24 |
0,10 |
0,20 |
|
|
10 |
0,13 |
0,05 |
0,15 |
0,27 |
0,06 |
0,06 |
0 |
0,02 |
0 |
0,01 |
0,22 |
0,10 |
0,19 |
|
|
11 |
0,10 |
0,08 |
0,29 |
0,40 |
0,09 |
0,07 |
0 |
0,02 |
0 |
0 |
0,26 |
0,08 |
0,23 |
|
|
12 |
0,24 |
0,10 |
0,10 |
0,15 |
0,08 |
0,18 |
0,15 |
0,19 |
0,16 |
0,14 |
0,22 |
0,11 |
0,01 |
|
|
13 |
0,21 |
0,11 |
0,20 |
0,27 |
0,13 |
0,10 |
0,06 |
0,09 |
0,09 |
0,05 |
0,11 |
0,1 |
0,10 |
|
|
14 |
0,21 |
0,10 |
0,14 |
0,16 |
0,08 |
0,17 |
0,16 |
0,19 |
0,15 |
0,15 |
0,22 |
0 |
0,08 |
Примечание. Над диагональю представлены значения F st , под диагональю — генетические дистанции Jost`s D. Группы индеек: 1 — черная тихорецкая порода (популяция 2011); 2 — черная тихорецкая порода (популяция 2025); 3 — узбекская палевая порода; 4 — серебристая северокавказская порода; 5 — бронзовая северокавказская порода (популяция 2011); 6 — бронзовая северокавказская порода (популяция 2025); 7 — белая широкогрудая порода; 8 — белая северокавказская порода; 9 — белая московская порода; 10 — кросс Виктория; 11 — кросс Универсал; 12 — кросс Big-6; 13 — кросс Big-10; 14 — кросс B.U.T.6.
Генетическую дифференциацию между группами индеек оценивали на основании попарных значений F st и генетических дистанций Jost`s D 278
(табл. 2, рис. 1).
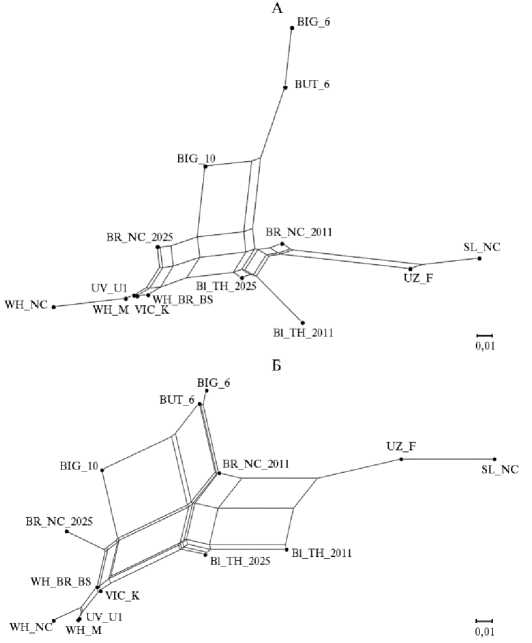
Рис. 1. Генетическая сеть, построенная по принципу ближайшего соседа на основе матрицы значений F st (А) и генетических дистанций Jost`s D (Б) , для исследуемых кроссов и пород индеек ( Meleagris gallopavo ): Bl_TH_2011 — черная тихорецкая порода (популяция 2011 года); Bl_TH_2025 — черная тихорецкая порода (популяция 2025 года); UZ_F — узбекская палевая порода; SL_NC — серебристая северокавказская порода; BR_NC_2011 — бронзовая северокавказская порода (популяция 2011 года); BR_NC_2025 — бронзовая северокавказская порода (популяция 2025 года); WH_BR_BS — белая широкогрудая порода; WH_NC — белая северокавказская порода; WH_M — белая московская порода; VIC_K — кросс Виктория; UV_U1 — кросс Универсал; BIG6 — кросс Big-6; BIG10 — кросс Big-10; BUT6 — кросс B.U.T.6 (ЦКП «Биоресурсы и биоинженерия сельскохозяйственных животных» ФГБНУ ФИЦ ВИЖ им. Л.К. Эрнста, 2025 год).
В структуре генетической сети, построенной на основе матрицы попарных значений Fst (см. рис. 1, А), выделялись три большие группы. Первая состояла из трех белых пород и двух кроссов (Виктория и Универсал). Белая северокавказская, белая московская и кросс Универсал соединялись с общей генетической сетью через узел кросса Виктория. Вторая группа была представлена кроссами B.U.T.6, Big-6 и Big-10. Кросс Big-10 был наиболее дифференцированным в этой группе, а ветвь, соответствующая кроссу Big-6, соединялась с общей генетической сетью через узел кросса B.U.T.6. Популяция бронзовой северокавказской породы 2025 года в виде короткой ветви располагалась между первой и второй группами. В состав третьей группы входили узбекская палевая, серебристая северокавказская породы, популяция бронзовой северокавказской породы 2011 года и отделенные в своей подгруппе обе популяции черной тихорецкой породы. Следует отметить, что большинство исследуемых групп индеек располагались на ребрах генетической сети, что указывает на их смешанное происхождение. Ветви формировали черная тихорецкая (популяция 2011 года), белая северокавказская и серебристая северокавказская породы, в также кроссы Универсал и Big-6.
В целом, генетическая дифференциация, визуализированная на основе матрицы попарных дистанций Jost`s D (см. рис. 1, Б), была сходна с полученной на основе значений F st. Тем не менее были выявлены и различия. Так, ветвь серебристой северокавказской породы соединялась с общей генетической сетью через ветвь узбекской палевой породы. Кроссы B.U.T.6 и Big-6 располагались на ребрах генетической сети, а кросс Big-10 был более сильно дифференцирован от них. Популяция 2025 года бронзовой северокавказской породы имела более длинную ветвь. Генетическая сеть, сформированная кроссом Универсал, белой московской и белой северокавказской породами, соединялась с другими группами посредством сети, образованной кроссом Виктория и белой широкогрудой породой.
Значительная генетическая дифференциация, выражающаяся значениями F st > 0,25, была выявлена между черной тихорецкой породой (популяция 2011 года) и кроссом B.U.T.6 (F st = 0,25, Jost`s D = 0,21), черной тихорецкой (популяция 2011 года) и серебристой северокавказской породами (F st = 0,26, Jost`s D = 0,23), кроссом B.U.T.6 и серебристой северокавказской породой (F st = 0,26, Jost`s D = 0,16), кроссами Универсал и Big-6 (F st = 0,26, Jost`s D = 0,22), кроссом B.U.T.6 и белой северокавказской породой (F st = 0,27, Jost`s D = 0,19), кроссом Big-6 и черной тихорецкой породой (популяция 2011 года) (F st = 0,27, Jost`s D = 0,24), кроссом BIG-6 и серебристой северокавказской породой (F st = 0,28, Jost`s D = 0,15), белой северокавказской и серебристой северокавказской породами (F st = 0,29, Jost`s D = 0,31), кроссом Big-6 и белой северокавказской породой (F st = 0,31, Jost`s D = 0,19) (см. табл. 2).
Тем не менее, согласно значениям генетических дистанций Jost`s D, существенная дифференциация также наблюдалась между черной тихорецкой (популяция 2011 года) и бронзовой северокавказской (популяция 2025 года) (Jost`s D = 0,24), черной тихорецкой (популяция 2011 года) и белой северокавказской породами (Jost`s D = 0,23), между узбекской палевой и белой московской породами (Jost`s D = 0,26), между узбекской палевой породой и кроссом Универсал (Jost`s D = 0,29). Кроме того, серебристая северокавказская порода была довольно удалена от большинства пород (Jost`s D = 0,25 с белой широкогрудой, Jost`s D = 0,27 с бронзовой северокавказской, популяция 2025 года, Jost`s D = 0,30 с белой московской) и кроссов (Jost`s D = 0,27 с кроссами Виктория и Big-6, Jost`s D = 0,40 с кроссом Универсал). Незначительная дифференциация (значения F st < 0,05), которая согласовывалась со значениями Jost`s D, наблюдалась между бронзовой северокавказской (популяция 2011 года) и черной тихорецкой (популяция 2025 года) породами, бронзовой северокавказской (популяция 2025 года) и белой широкогрудой породами, белой северокавказской и белой московской породами, бронзовой северокавказской породой (популяция 2025 года) и кроссом Виктория, бронзовой северокавказской (популяция 2025 года) и кроссом Универсал, белой северокавказской породой и кроссом Виктория, белой северокавказской породой и кроссом Универсал.
Анализ исследуемых выборок не выявил генетической дифференциации по обоим изучаемым показателям между серебристой северокавказской и узбекской палевой породами, белой северокавказской и белой широкогрудой породами, кроссами B.U.T.6 и Big-6, кроссом Виктория и белой широкогрудой породой, кроссом Универсал и белой широкогрудой породой, кроссом Универсал и белой московской породой, кроссами Универсал и Виктория.
Результаты кластерного анализа в целом согласовывались с особенностями структуры генетической сети (рис. 2).
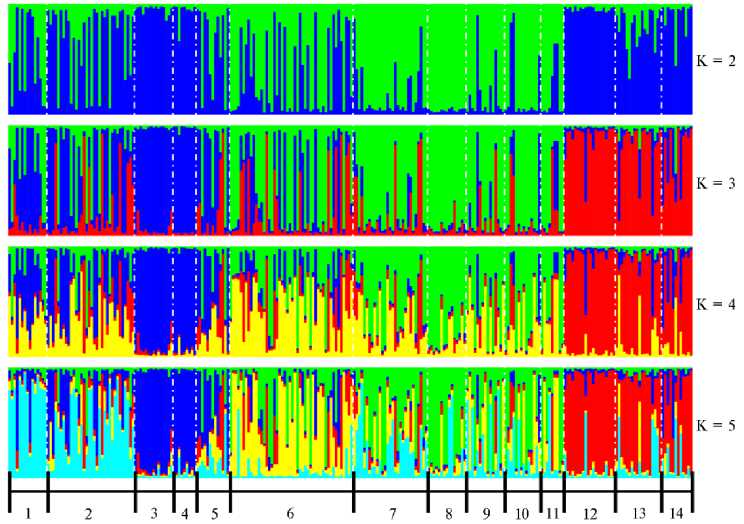
Рис. 2. Популяционная структура кроссов и пород индеек ( Meleagris gallopavo ), разводимых в России, при числе кластеров от K = 2 до K = 5: 1 — черная тихорецкая порода (популяция 2011 года); 2 — черная тихорецкая порода (популяция 2025 года); 3 — узбекская палевая порода; 4 — серебристая северокавказская порода; 5 — бронзовая северокавказская порода (популяция 2011 года); 6 — бронзовая северокавказская порода (популяция 2025 года); 7 — белая широкогрудая порода; 8 — белая северокавказская порода; 9 — белая московская порода; 10 — кросс Виктория; 11 — кросс Универсал; 12 — кросс Big-6; 13 — кросс Big-10; 14 — кросс B.U.T.6 (ЦКП «Биоресурсы и биоинженерия сельскохозяйственных животных» ФГБНУ ФИЦ ВИЖ им. Л.К. Эрнста, 2025 год).
При K = 2 были сформированы два кластера. Первый (преобладающий синий цвет) состоял из узбекской палевой и серебристой северокавказской пород, обеих популяций черной тихорецкой породы и популяции 2011 года бронзовой северокавказской породы, а также кроссов B.U.T.6, Big-6, Big-10; второй (преобладающий зеленый) — из белой широкогрудой, белой северокавказской, белой московской пород и популяции 2025 года бронзовой северокавказской породы, кроссов Универсал и Виктория. При K = 3 четко отделились кластеры, первый из которых включал кроссы B.U.T.6, Big-6, Big-10 (красный цвет), второй — узбекскую палевую и серебристую северокавказскую породы (синий цвет). Остальные породы и кроссы имели неоднородную популяционную структуры. Популяции черной тихорецкой породы и популяция 2011 года бронзовой северокавказской породы характеризовались наличием более выраженного предкового компонента, который соответствовал синему цвету. При K = 4, несмотря на появление нового предкового компонента, в целом наблюдались схожие тенденции, как и при K = 3.
Согласно значениям ошибки кросс-валидации, наиболее вероятное число кластеров в исследуемой выборке составило 5. При K = 5 популяции черной тихорецкой породы сформировали свой, хотя и не полностью однородный, но дифференцируемый кластер (преобладающий голубой цвет). Популяция 2025 года бронзовой северокавказской породы дифференциро- валась от группы, состоящей из белой широкогрудой, белой северокавказской, белой московской пород, кроссов Универсал и Виктория. Тем не менее популяции 2011 и 2025 годов бронзовой северокавказской породы в целом различались по генетической структуре.
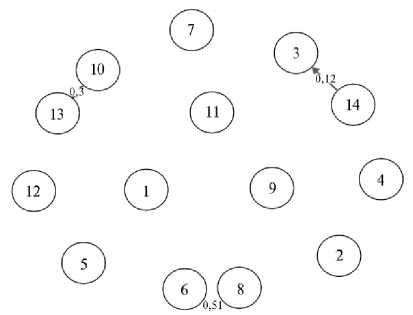
Рис. 3. Относительная миграция генов между исследуемыми породами и кроссами индеек ( Meleagris gallopavo ) на основе оценки числа мигрантов на поколение: 1 — черная тихорецкая порода (популяция 2011 года); 2 — черная тихорецкая порода (популяция 2025 года); 3 — узбекская палевая порода; 4 — серебристая северокавказская порода; 5 — бронзовая северокавказская порода (популяция 2011 года); 6 — бронзовая северокавказская порода (популяция 2025 года); 7 — белая широкогрудая порода; 8 — белая северокавказская порода; 9 — белая московская порода; 10 — кросс Виктория; 11 — кросс Универсал; 12 — кросс Big-6; 13 — кросс Big-10; 14 — кросс B.U.T.6. (ЦКП «Биоресурсы и биоинженерия сельскохозяйственных животных» ФГБНУ ФИЦ ВИЖ им. Л.К. Эрнста, 2025 год). Цифры вне кругов обозначают доли интрогрессии генов, стрелка — направление интрогрессии.
Была выявлена небольшая односторонняя интрогрессия генов от кросса B.U.T.6 в узбекскую палевую породу (0,12) (рис. 3). Кроме того, наблюдался интенсивный обмен генетическим материалом между популяцией 2025 года бронзовой северокавказской породы и белой северокавказской породой (0,51), а также между кроссами Big-10 и Виктория (0,30).
В России разводят семь пород и десять кроссов индеек. Несмотря на то, что популяционно-генетические исследования пород индеек были проведены ранее (5, 20, 31), отсутствовал мониторинг с учетом популяционной динамики, а также сравнительный анализ генетического разнообразия с кроссами. В рамках реализации новой программы по развитию био-ресурсных коллекций начаты активные научно-исследовательские работы по генетическому мониторингу ранее не охваченных генетических ресурсов животных, сохраняемых ex situ, с использованием различных молекулярногенетических подходов.
В настоящей работе мы провели анализ полиморфизма шести локусов: MNT036, MNT0115, MNT0247, MNT0348, MNT0370, WT54. Все исследуемые локусы оказались полиморфными. Аллельное разнообразие варьировало от 3,000 до 4,284, что было выше степени полиморфизма, рассчитанной при использовании локусов MNT9-MNT20 в отечественных породах индеек (5). В работах других авторов средняя наблюдаемая гетерозиготность варьировала от 0,452±0,229 до 0,667±0,265 у местных американских и европейских индеек (16) и 0,286 в популяции генофондной фермы Университета Миннесоты (University of Minnesota) (21). В популяциях трех итальянских пород индеек Colli Euganei, Nero d’Italia и Brianzolo были отмечены низкие значения биологического разнообразия. Среднее число аллелей на локус варьировало от 1,68 в породе Colli Euganei до 2,18 у Brianzolo, ожидаемая гетерозиготность — от 0,203 до 0,250 в тех же породах (32). A.M. Canales с соавт. (19) продемонстрировали, что домашние индейки из Андалусии, Мексики, США и дикие индейки характеризовались высокими значениями наблюдаемой гетерозиготности (соответственно 0,422; 0,521; 0,429 и 0,627) и аллельного разнообразия (6,50; 6,76; 8,35 и 5,09). В нашей работе показатели наблюдаемой гетерозиготности составили 0,319-0,620, что было выше значений, выявленных в отечественных породах другими исследователями. Так, согласно В.И. Фисинину с соавт. (5), значения наблюдаемой гетерозиготности варьировали от 0,063 до 0,273 в группах бронзовой северокавказской и белой широкогрудой пород.
В целом следует отметить, что популяции домашних индеек, выведенных в различных странах, характеризуются сопоставимыми значениями генетического разнообразия по сравнению с другими видами одомашненной птицы: курицей ( Gallus gallus domesticus ), уткой ( Anas platyrhynchos ) и гусем ( Anser anser ). Например, анализ генетического разнообразия в популяциях местных средиземноморских кур показал, что среднее число аллелей на локус варьировало от 2,63 в породе Modenese до 4,96 в породах Sureсa и Albanian, а наблюдаемая гетерозиготность — от 0,35 в породах Combatiente Espaсol и Maltese Black до 0,61 в породе Banat Nacked Neck. Что касается коммерческих яичных и бройлерных линий, то среднее число аллелей на локус и наблюдаемая гетерозиготность составили 3,03 ± 0,38 и 0,42±0,09 (33). По данным D.A. Carvalho с соавт. (34), при генотипировании бразильских кур среднее число аллелей на локус варьировало от 4,960 для пород Canela-Preta и Peloco до 5,040 для Caneluda do Catol e . Наблюдаемая гетерозиготность составила 0,617, 0,646 и 0,642 для бразильских пород кур (34). Было показано, что у пекинских уток среднее число аллелей на локус составило 4,18 (35), а у индийских уток — 4,48 (36). В трех популяциях гусей из провинции Ляонин (Китай) число аллелей на локус и наблюдаемая гетерозиготность варьировали соответственно от 5,083 до 6,083 и от 0,149 до 0,184 (37).
При изучении генетических связей были выявлены закономерности, соответствующие происхождению групп индеек. Так, кроссы иностранного происхождения ожидаемо сформировали свой дифференцированный кластер (B.U.T.6, Big-6, Big-10), в то время как кроссы отечественной селекции кластеризовались с белыми породами индеек. Была отмечена удаленность серебристой северокавказской породы от большинства изучаемых групп. Серебристая северокавказская порода индеек была создана на основе узбекской палевой и белой широкогрудой пород (7). Анализируя исследуемую выборку, мы обнаружили выраженное генетическое сходство серебристой северокавказской и узбекской палевой пород, тогда как с белой широкогрудой породой была отмечена умеренная генетическая дифференциация. По мнению В.И. Фисинина с соавт (5), определенная генетическая дифференциация черной тихорецкой и узбекской палевой пород может быть связана с тем, что при их создании были использованы популяций местных индеек.
Результаты всех проведенных анализов продемонстрировали, что две популяции черной тихорецкой породы кластеризовались совместно, однако в отношении популяций бронзовой северокавказской породы наблюдалась некоторая удаленность и отличия в генетической структуре, что свидетельствует о генетических изменениях, произошедших в породе. Выявленный поток генов между популяцией 2025 года бронзовой северокавказской породы и белой северокавказской породой, вероятно, мог быть связан с тем, что обе эти породы были созданы на основе местных аборигенных индеек (5). В то же время в сочетании с большим генетическим и аллельным разнообразием можно предположить, что происходит скрещивание с другими породами (например, с белой кавказской). Вероятно, расширение количества микросателлитных локусов позволит более точно понять причины этого наблюдения.
Таким образом, генетическая дифференциация между исследуемыми породами и кроссами индеек ( Meleagris gallopavo ), оцененная на основе расчета значений F st и генетических дистанций Jost`s D, варьировала от незначительной до высокой. Большинство изученных пород индеек характеризовались высокой дифференциацией с иностранными кроссами. Выявлена тесная генетическая связь между узбекской палевой и серебристой северокавказской породами. Показана совместная кластеризация отечественных кроссов с породами с белым окрасом оперения, тогда как иностранные промышленные кроссы формировали свою обособленную группу. Выявлено, что отечественные кроссы Виктория и Универсал характеризовались сопоставимыми значениями генетического разнообразия и более высокими значениями аллельного разнообразия по сравнению с иностранными промышленными кроссами. В нашей работе для двух генофондных пород индеек — бронзовой северокавказской и черной тихорецкой были доступны образцы крови, датируемые 2011 годом (архивная популяция) и отобранные в 2025 году (современная популяция). Показано, что в современной популяции черной тихорецкой породы произошли меньшие изменения по сравнению архивной. Обе популяции этой породы сохраняли схожую генетическую структуру и характеризовались близкими значениями наблюдаемой гетерозиготности. Тем не менее в современной популяции черной тихорецкой породы были отмечены более высокие значения аллельного разнообразия. Современная популяция бронзовой северокавказской породы (2025 года) индеек характеризировалась более высокими значениями генетического и аллельного разнообразия по сравнению с архивной популяцией этой породы, в которой, в свою очередь, были выявлены достоверные значения инбридинга. Также отмечены различия в генетической структуре архивной и современной популяций этой породы, причины которых будут изучены более подробно в наших следующих работах посредством расширения спектра анализируемых микросателлитных локусов и анализа последовательностей полного генома.
