Характеристика популяции С-клеток щитовидной железы потомства самок крыс с хроническим экспериментальным поражением печени различного генеза

Автор: Солянникова Д.Р., Брюхин Г.В.
Журнал: Человек. Спорт. Медицина @hsm-susu
Рубрика: Проблемы здравоохранения
Статья в выпуске: 39 (172), 2009 года.
Бесплатный доступ
Изучено влияние хронического экспериментального поражения печени матери различного генеза на морфофункциональное состояние популяции С-клеток щитовидной железы потомства. При этом использовались модели поражения печени с помощью E.coli и D-галактозамина. Анализ полученных в ходе проведенного исследования данных позволяет сделать вывод о том, что у потомства матерей с хроническим поражением печени различного генеза наблюдается увеличение количества С-клеток щитовидной железы, а также увеличение доли секреторно активных форм эндокриноцитов.
Хронические заболевания печени, щитовидная железа, парафоллику-лярные с-клетки, крыса
Короткий адрес: https://sciup.org/147152624
IDR: 147152624 | УДК: 616.36-002.2-092.9-06:616.441-053.31
Characteristics of the thyroid C-cell population of the posterity of female rats with long-lasting liver injury of various genesis
Influence of long-lasting experimental liver injury of the mother to morpho-functional state of thyroid gland C-cell population of its posterity was studied. The models of liver injury with E.coli and D-galactosamine were considered. From the analysis of experimental data it is concluded that in posterity from female rats with long-lasting liver injury of various genesis the increase of quantity of thyroid gland C-cell population and endo-crynocytes with high secretory activity was established.
Текст научной статьи Характеристика популяции С-клеток щитовидной железы потомства самок крыс с хроническим экспериментальным поражением печени различного генеза
В настоящее время в ходе реализации государственного проекта «Здоровье» особое внимание уделяется проблеме воспроизводства здорового потомства. При этом особая роль должна отводиться здоровью будущих матерей. Одним из очевидных препятствий для безопасного и эффективного материнства является экстрагенитальная патология, частота которой среди беременных женщин, несмотря на их молодой возраст, очень высока и составляет 35-50 %, а по некоторым данным даже 60-80 % [8]. Особое место в экстрагениталь-ной патологии занимают болезни печени, самыми распространенными формами которых являются хронические гепатиты [7, 12]. Экспериментальным путем было доказано, что у потомства лабораторных животных, имеющих поражение печени различного генеза, отмечаются нарушения структурно-функциональной организации различных органов и систем: селезенки [2], тимуса [4], яичников [3], семенников [5] и другие. Особое место в координации и регуляции деятельности организма отводится эндокринной системе. Самой крупной эндокринной железой является щитовидная железа, которая во многом определяет нормальное функционирование практически всех органов и систем организма, в том числе и организма развивающегося плода. При этом особое влияние железа оказывает на развитие и функционирование нервной системы в антенатальном и раннем постнатальном периодах. В связи с этим нарушение становления и функционирования щитовидной железы может привести к нарушению развития плода и ребенка.
Основную массу щитовидной железы образует собственно тиреоидная паренхима, в значительно меньшем количестве в органе представлена система парафолликулярных С-клеток, являющая ся, однако, ее неотъемлемым компонентом. Основной функцией парафолликулярных клеток является выработка гипокальциемического фактора -тиреокальцитонина. Помимо этого, парафолликулярные клетки синтезируют и выделяют целый ряд других биологически активных веществ, таких как соматостатин, вещество Р, катакальцин I и II, гаст-рин-релизинг пептид, тиролиберин и др., являющиеся, наряду с кальцитонином, факторами регуляции интраорганного гомеостаза. В связи с этим целью нашего исследования явился анализ морфофункционального состояния популяции парафолликулярных С-клеток щитовидной железы потомства самок крыс с хроническим экспериментальным поражением печени различного генеза.
Материалы и методы исследования. В эксперименте были использованы белые лабораторные, половозрелые крысы (самки) «Вистар» и их потомство на 1, 15, 30, 45 и 60-е сутки. Животные были разделены на 3 группы: 1) потомство от интактных матерей (контрольная группа); 2) потомство от матерей с хроническим экспериментальным поражением печени с помощью E.coli (группа «E.coli») и 3) потомство от матерей с хроническим поражением печени с помощью D-галактозамина (группа «D-галактозамин»). Модель хронического поражения печени с помощью E.coli создавали путем внутрипеченочного введения 0,2 мл фильтрата 6-дневной культуры E.coli и последующим введением через сутки в хвостовую вену того же фильтрата E.coli в количестве 0,3 мл/кг массы тела. Возникающие морфологические и функциональные изменения, согласно данным литературы, обнаруживают сходство с таковыми при гепатите А [10]. Вторую модель поражения печени создавали путем внутрибрюшинного введения D-галактозамина гидрохлорида в количестве 250 мг/кг массы тела. Экспериментальный гепатит, вызываемый введением D-галактозамина гидрохлорида по своим морфологическим, гистологическим и биохимическим характеристикам рассматривается как адекватная модель вирусного гепатита В у человека [6, 11, 13]. Поражение гепатобилиарной системы экспериментальных животных верифицировали с помощью морфологических, биохимических и иммунологических методов исследования. Серийные гистологические срезы щитовидной железы толщиной 5-6 мкм окрашивали нитратом серебра по Гримелиусу в модификации Никонова [1]. Подсчитывали абсолютное количество парафолликулярных клеток на условную единицу площади -1251,6 мкм2, а также оценивали субпопуляционный состав С-клеток по степени и характеру их насыщения аргирофильными гранулами [9]. Статистическая обработка проводилась с помощью непараметрического критерия Манна-Уитни.
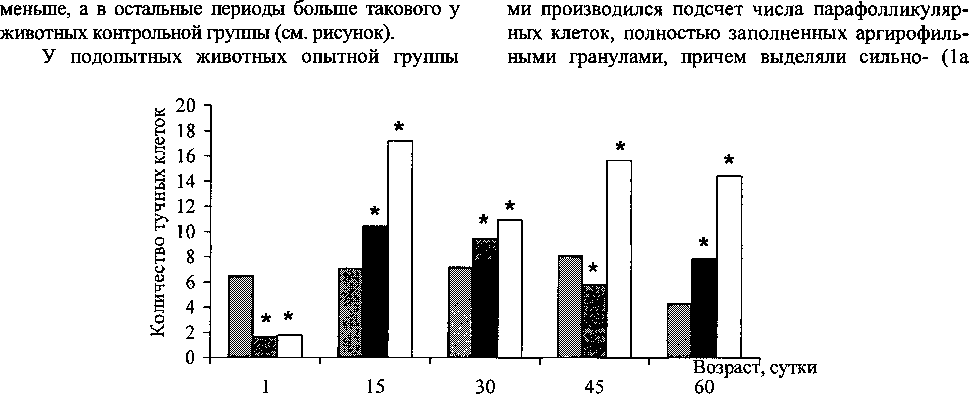
Результаты исследования и их обсуждение. Установлено, что у интактных крысят количество С-клеток в единице условной площади после рождения увеличивается с 6,53 ± 0,469 в период новорожденности до 8,10 ± 0,628 к концу периода полового созревания, а затем снижается до 4,30 ±0,159 в период половой зрелости (см. рисунок).
У экспериментальных животных группы «Е.соП» после рождения наблюдается резкое увеличение количества парафолликулярных клеток с 1,67 ± 0,412 в период новорожденности до 10,39 ± 1,224 в подсосный период, после чего количество С-клеток в течение периода полового созревания несколько снижается, а уже к периоду половой зрелости число данных клеток вновь возрастает. При этом в период новорожденности и в конце периода полового созревания количество парафолликулярных клеток щитовидной железы статистически достоверно
«D-галактозамин» количество парафолликулярных клеток после рождения, так же как и в первой подопытной группе, резко увеличивается с 1,80 ± 0,501 в период новорожденное™ до 17,22 ± 0,719 к 15-м суткам, но уже к началу периода полового созревания исследуемый показатель начинает снижаться, а к концу данного периода - вновь увеличиваться, после чего показатель стабилизируется. При этом важно отметать, что во все возрастные периоды, за исключением периода новорожденно-сти, количество С-клеток в щитовидной железе животных группы «D-галактозамин» больше количества данных клеток у контрольных животных (см. рисунок).
Как видно из рисунка, в период новорожден-ности у подопытных крысят количество парафолликулярных клеток щитовидной железы в единице условной площади значительно снижено по сравнению с контрольным значением. На 15-е и 30-е сутки исследования содержание С-клеток у подопытных животных превышает таковое в контроле, причем к 15-м суткам наблюдается значительный рост количества клеток, особенно в группе «D-галактозамин». К концу периода полового созревания число парафолликулярных клеток в щитовидной железе крысят группы «E.coli» снижается до уровня более низкого, а в группе «D-галактозамин» наоборот растет до более высокого, чем у интактных животных. В период половой зрелости количество С-клеток у животных обеих подопытных групп превышает число клеток в контроле, причем вновь данный показатель значительно выше у животных группы «D-галактозамин».
Одним из показателей функциональной активности парафолликулярных клеток является уровень и особенности их гранулярного насыщения. При анализе субпопуляционного состава С-клеток по степени и характеру гранулярного насыщения на-
®контроль ^группа "E.coli" □ группа "D-галактозамин
Количество парафолликулярных клеток щитовидной железы экспериментальных животных. * - результаты статистически достоверны (Р<0,05)
Таблица 1
Субпопуляционный состав парафолликулярных клеток щитовидной железы интактных животных по степени и характеру гранулярного насыщения (%)
|
Тип клеток |
Возраст, сутки |
||||
|
1 (п=Ю) |
15 (п=10) |
30 (п=10) |
45 (п=10) |
60 (п=Ю) |
|
|
1а тип клеток |
19,23 ±4,040 |
21,92 ±2,073 |
17,10 ± 1,402 |
19,35 ± 1,209 |
24,95 ± 1,202 |
|
16 тип клеток |
44,38 ± 1,994 |
46,60 ±2,753 |
51,79 ±1,761 |
55,49 ± 1,620 |
49,68 ± 0,761 |
|
2-й тип клеток |
14,50 ±2,822 |
4,98 ± 1,137 |
7,02 ± 0,803 |
3,44 ±0,453 |
6,12 ±0,463 |
|
3-й тип клеток |
6,03 ± 1,095 |
8,73 ± 0,528 |
9,37 ±0,737 |
8,67 ±0,766 |
10,15 ± 0,420 |
|
4-й тип клеток |
15,87 ±4,087 |
17,78 ± 1,167 |
14,77 ± 1,049 |
12,80 ± 1,509 |
9,13 ±0,692 |
* - результаты статистически достоверны (Р < 0,05).
тип) и умеренногранулированные (16 тип) формы; клеток с преимущественной концентрацией гранул на тироцитарном (2-й тип), а также сосудистом (3-й тип) полюсах и клеток с единичными гранулами (4-й тип клеток) [9]. Полученные результаты отражены в табл. 1-3.
Как видно из табл. 1, у животных контрольной группы в популяции парафолликулярных клеток щитовидной железы на всех сроках исследования преобладают клетки 16 типа, количество которых незначительно увеличивается с периода ново-рожденности до конца периода полового созревания, после чего их содержание несколько снижается. Число клеток 1а типа с возрастом постепенно увеличивается с 19,23 ± 4,040 % в период новорожденности до 24,95 ±1,202 % в период половой зрелости, за исключением начала периода полового созревания, когда наблюдается снижение данного показателя. Изменение содержания С-клеток 2-го типа носит зигзагообразный характер. С возрастом количество данных клеток снижается, за исключением начала периода полового созревания и периода половой зрелости, когда отмечается увеличение количества парафолликулярных клеток. Количество С-клеток 3-го типа увеличивается после рождения с 6,03 ± 1,095 % в период новорожденное™ до 10,15 ±0,420 % в период половой зрелости, исключение составляет конец периода полового созревания. Содержание С-клеток 4-го типа после рождения растет до 15-х суток, когда и достигает максимального значения (17,78 ± 1,167 %), после чего показатель неуклонно снижается вплоть до периода половой зрелости (см. табл. 1).
У подопытных животных экспериментальной группы «Е.соИ» количество С-клеток типа 1а с возрастом увеличивается с 4,22 ± 2,356 % в период новорожденное™ до 9,97 ± 2,249 % в период половой зрелости, за исключением конца периода полового созревания, когда данный показатель снижается. Число С-клеток типа 16 после рождения до 15-х суток резко снижается, затем несколько возрастает и стабилизируется. Изменение содержания парафолликулярных клеток 2-го тапа носит зеркальных характер относительно динамики данных клеток в контроле. Количество С-клеток 3-го типа увеличивается с возрастом с 12,00 ± 1,310 % в период ново- рожденности до 17,40 ± 1,389 % в конце периода полового созревания, после чего исследуемый показатель снижается. Число С-клеток 4-го типа с возрастом увеличивается с 24,04 ± 5,239 % в период новорожденности до 36,63 ± 2,109 % в период половой зрелости, исключение составило начало периода полового созревания, когда наблюдается резкое снижение количества данного типа клеток.
Таким образом, у экспериментальных животных подопытной группы «Е.соИ» наблюдается значительное снижение доли С-клеток 1а и 16 типа и увеличение клеток 3-го и особенно 4-го типов, которые в некоторые возрастные периоды (15-е и 60-е сутки) являются доминирующими клетками данной популяции. Содержание парафолликулярных клеток 2-го типа в период новорожденности и в начале периода полового созревания снижено, на 15-е сутки и в конце периода полового созревания повышено по сравнению с контролем, а в период половой зрелости практически равно ему (табл. 2). Полученные данные говорят о переходе значительного числа элементов популяции в фазу активной секреции. При этом биологически активные вещества наиболее активно выделяются в тироцитарном направлении, а не сосудистом, и, таким образом, осуществляют преимущественно паракриновое воздействие.
У экспериментальных животных группы «D-галактозамин» количество С-клеток 1а типа с возрастом постепенно снижается с 9,55 ±2,176 % в период новорожденности до 7,67 ± 0,854 % в период половой зрелости, исключение составили 30-е сутки, когда наблюдается увеличение значения исследуемого показателя. Содержание парафолликулярных клеток 16 типа после рождения, наоборот, увеличивается с 40,77 ± 7,480 % в период новорожденности до 47,93 ± 1,381 % в период половой зрелости, опять же за исключением начала периода полового созревания. Число С-клеток 2-го типа уменьшается с 7,63 ± 2,983 % у 1-дневных крысят до 2,39 ± 0,364 % у 60-дневных животных, исключение вновь составило начало периода полового созревания. Количество С-клеток 3-го типа возрастает после рождения с 7,63 ± 1,643 % в период новорожденности до 13,03 ± 2,097 % к началу периода полового созревания, после чего ис-
Таблица 2
Субпопуляционный состав парафолликулярных клеток щитовидной железы животных группы «Е.соК» по степени и характеру гранулярного насыщения (%)
|
Тип клеток |
Возраст, сутки |
||||
|
1 (п= 10) |
15 (п=10) |
30 (и = 10) |
45 (п = Ю) |
60 (п = 7) |
|
|
1а тип клеток |
4,22 ± 2,356* |
5,65 ± 1,050* |
10,96 ±2,078 |
7,85 ± 1,476* |
9,97 ± 2,249* |
|
16 тип клеток |
53,06 ±5,183 |
32,34 ±2,660* |
37,72 ± 1,414* |
34,30±2,460* |
36,20 ±3,164* |
|
2-й тип клеток |
3,33 ± 1,746* |
8,98 ± 1,239* |
6,45 ± 0,726 |
9,35 ± 0,762* |
6,10 ±0,671 |
|
3-й тип клеток |
12,00 ± 1,310* |
14,14 ±0,745* |
14,44 ± 0,539* |
17,40 ± 1,389* |
11,09 ±0,785 |
|
4-й тип клеток |
24,04 ± 5,239 |
38,89 ±3,009* |
30,64 ± 1,066* |
31,09 ±0,838* |
36,63 ±2,109* |
* - результаты статистически достоверны (Р<0,05).
Таблица 3
Субпопуляционный состав парафолликулярных клеток щитовидной железы животных группы «D-галактозамин» по степени и характеру гранулярного насыщения (%)
|
Тип клеток |
Возраст, сутки |
||||
|
1 (п=10) |
15 (п=10) |
30 (п = 6) |
45 (п= 10) |
60 (п-7) |
|
|
1а тип клеток |
9,55 ±2,176 |
6,55 ± 1,030* |
8,53 ± 1,739* |
8,44 ±0,821* |
7,67 ± 0,854* |
|
16 тип клеток |
40,77 ± 7,480 |
46,06 ± 1,614 |
35,60 ±5,011* |
42,79 ± 1,832* |
47,93 ± 1,381 |
|
2-й тип клеток |
7,63 ± 2,983 |
4,89 ± 0,784 |
12,25 ± 1,597* |
9,90 ± 0,924* |
2,39 ± 0,364* |
|
3-й тип клеток |
7,63 ± 1,643 |
9,44 ± 0,848 |
13,03 ± 2,097 |
8,95 ± 0,279 |
6,56 ±0,346* |
|
4-й тип клеток |
21,10 ±4,753 |
33,06 ±0,988* |
30,58 ± 1,115* |
29,87 ± 1,324* |
35,47 ± 1,071* |
* - результаты статистически достоверны (Р<0,05).
следуемый показатель снижается до 6,56 ± 0,346 % к периоду половой зрелости. Количество парафолликулярных клеток 4-го типа значительно увеличивается после рождения до 15-х суток с 21,10 ± 4,753 % до 33,06 ± 0,988 % соответственно. К 30-м суткам количество данных клеток несколько снижается и на всем протяжении периода полового созревания практически не изменяется, но уже к периоду половой зрелости число С-клеток 4-го типа вновь возрастает. Таким образом, у экспериментальных животных подопытной группы «D-галактозамин», так же как и у животных первой подопытной группы наблюдается значительное снижение доли С-клеток 1а и 16 типа и увеличение клеток 3-го (за исключение 60-х суток) и особенно 4 типов, которые, однако, не будут являться доминирующими формами данной популяции ни в один возрастной период. Количество С-клеток 2-го типа до 30-х суток и в период половой зрелости снижено, а на протяжении всего периода полового созревания повышено, по сравнению с контрольными значениями (табл. 3). Полученные данные, так же как и в первой подопытной группе, говорят о переходе популяции С-клеток в фазу активной секреции, причем вновь наблюдается преимущественный выброс гранул с биологически активными веществами в тироцитарном направлении, за исключением конца периода полового созревания.
Таким образом, в ходе проведенного исследования было выяснено, что у потомства самок крыс с хроническим экспериментальным поражением печени различного генеза наблюдается изменение количества парафолликулярных клеток щитовидной железы и их субпопуляционного состава по степени и характеру гранулярного насыщения. На большинстве сроков исследования отмечается увеличение числа С-клеток у животных обеих подопытных групп, особенно у крысят группы «D-галактозамин». Также выявлено изменение субпопуляционного состава данных клеток у животных обеих подопытных групп в сторону значительного увеличения числа секреторно активных клеточных форм. При этом наибольшие изменения в составе популяции парафолликулярных клеток щитовидной железы наблюдаются у животных группы «Е.соИ».
Список литературы Характеристика популяции С-клеток щитовидной железы потомства самок крыс с хроническим экспериментальным поражением печени различного генеза
- Автандилов Г.Г. Патогистологическая техника/Г.Г. Автандилов. -М.: Медицина, 1994. -303 с.
- Брюхин Г.В. Влияние хронической интоксикации матери на морфо-функционалъные особенности тучных клеток селезенки потомства/Г.В. Брюхин, Е.Н. Пашнина, Н.В. Леонов//Морфологические ведомости. -2004. -№ 1-2. -С. 16.
- Брюхин Г В. Особенности становления фолликулогенеза в яичниках у потомства матерей с хроническим поражением гепатобилиарной системы в условиях эксперимента/Г.В. Брюхин, Е.В. Вторушина//Проблемы репродукции. -2005. -№ 2. -С. 23-26.
- Брюхин Г.В. Характеристика пролиферативной активности тимоцитов и лимфоцитов периферической крови потомства самок с хроническим экспериментальным поражением печени различной этиологии/Г.В. Брюхин, А.А. Федосов//Морфология. -2006. -№1.-С. 57-59.
- Брюхин Г.В. Влияние хронической экспериментальной патологии печени матери на эндокринную функцию мужских половых желез потомства/Г.В. Брюхин, М.Л. Сизоненко//Проблемы репродукции. -2008. -№2.-С. 45-47.
- Венгеровский А.И. Метаболизм липидов и функциональное состояние печени при интоксикации D-галактозамином у крыс/А.И. Венгеровский, А.С. Сарактиков//Патологическая физиология и экспериментальная терапия. -1988. -№3.-С. 52-54.
- Игнатова Т.М. Хронические заболевания печени беременных/Т.М. Игнатова//Тер. Архив. -2002.-М 10. -С. 55-59.
- Медведь В.И. Введение в клинику экстрагениталъной патологии беременных/В. И. Медведь. -Киев: Авиценна. -2004. -168 с.
- Павлов А.В. Цитологический анализ популяции парафолликулярных клеток щитовидной железы/А. В. Павлов//Цитология. -1985. -Т. 27, №11.-С. 1300-1303.
- Моделирование воспалительного процесса в печени. Моделирование, методы изучения и экспериментальная терапия патологических процессов/Б.А. Саков, А.И. Поляк, В.Е. Рычнев и др. -М., 1967.-4.1.-251 с.
- Влияние убихинона-10 на развитие D-галактозаминового гепатита у крыс/Н.П. Сугробова, Р.С. Медведник, Л.В. Ефимова и др.//Бюллетень экспериментальной биологии и медицины. -1992. -№11. -С. 504-506.
- Шехтман М.М. Руководство по экстрагенитальной патологии у беременных/М.М. Шехтман.-М., 1999.-818 с.
- Mieke Joker A. Immunopathology of Acute Galactosamine Hepatitis in Rats/Mieke Joker, A. and other//Hepatology. -1990. -V. 11, №4.-P. 622-627.
