Идеальное свободное распределение в модели «хищник–жертва» с эффектом Олли

Автор: Зеленчук П.А.
Журнал: Российский журнал биомеханики @journal-biomech
Статья в выпуске: 1 т.30, 2026 года.
Бесплатный доступ
Рассмотрено влияние сильного эффекта Олли в популяции жертвы на динамику модели «хищник–жертва» с идеальным свободным распределением (ИСР). Приводится сравнение стационарных решений для двух систем: с эффектом Олли и без него. Показано, что жертва всегда сохраняет ИСР, в то время как хищник имеет сложные ИСР–подобные распределения, зависящие от функции ресурса и параметра Олли. Наличие сильного эффекта Олли у жертвы приводит к смене устойчивости тривиального нулевого состояния равновесия и появлению нового неустойчивого решения. При этом диапазон устойчивости стационарного состояния, отвечающего сосуществованию двух видов, уменьшается на величину пропорциональну параметру Олли и обратно пропорциональную ресурсу. Найдено, что изменение параметра Олли напрямую влияет на численность популяции хищника (чем больше параметр, тем меньше численность), в то время как популяция жертвы остается неизменной. Установлено, что с увеличением параметра Олли для некоторых точек ареала (в силу зависимости функции ресурса от пространственной координаты) стационарное решение с двумя видами становится неустойчивым для малых возмущений системы. В результате распределение популяций содержит области, в которых их численность равна нулю. Приведены вычислительные эксперименты, демонстрирующие наличие осциллирующего режима для двух видов. Система без эффекта Олли сохраняет колебания и общий вид пространственного распределения при малых возмущениях начальных условий. При сильном эффекте Олли возможны локальные колебания, вызванные тем, что условие их появления зависит от изменяющейся по пространству функции ресурса.
Идеальное свободное распределение, эффект Олли, «хищник–жертва», стационарное состояние, неоднородный ареал
Короткий адрес: https://sciup.org/146283285
IDR: 146283285 | УДК: 57.05:519.62:517.9 | DOI: 10.15593/RZhBiomeh/2026.1.14
Ideal free distribution in the predatorprey model with the allee effect
The influence of a strong Allee effect in a prey population on the dynamics of a predator–prey model with ideal free distribution (IFD) is considered. A comparison of stationary solutions is given for two systems: with and without the Allee effect. It is shown that the prey always maintains its IFD, while the predator has complex IFD-like distributions depending on the resource function and the Allee parameter. The presence of a strong Allee effect in the prey leads to a change in the stability of the triv-ial zero equilibrium and the emergence of a new unstable solution. In this case, the range of stability of the stationary state corresponding to the coexistence of two species decreases by an amount proportional to the Allee parameter and inversely proportional to the resource. It was found that changing the Allee parameter directly affects the population size of the predator (the larger the parameter, the smaller the population), while the population of the prey remains un-changed. It was found that with increasing Allee's parameter, for some points in the range (due to the dependence of the resource function on the spatial coordinate), the stationary solution with two species becomes unstable for small perturbations of the system. As a result, the population dis-tribution contains regions where their abundance is zero. Computational experiments demonstrating the presence of an oscillatory regime for two species are presented. A system without the Allee effect maintains oscillations and the general form of its spatial distribution under small perturbations of the initial conditions. With a strong Allee effect, local oscillations are possible, caused by the fact that the condition for their occur-rence depends on a spatially varying resource function.
Текст научной статьи Идеальное свободное распределение в модели «хищник–жертва» с эффектом Олли
Теоретическое описание пространственных распределений живых организмов в неоднородной среде обитания – одна из важнейших проблем популяцион- ной динамики и эволюционной экологии [1; 2]. Идеальное свободное распределение (ИСР) – одна из фундаментальных теоретических концепций, описывающих механизмы распределения особей, направленные на максимизацию их приспособленности к условиям


Эта статья доступна в соответствии с условиями лицензии Creative Commons Attribution-NonCommercial 4.0 International
License (CC BY-NC 4.0)
This work is licensed under a Creative Commons Attribution-NonCommercial 4.0 International License (CC BY-NC 4.0)
среды с учётом биотических, абиотических и антропогенных факторов [3]. В 1969 году Стивен Фретуэлл и Генри Лукас-младший опубликовали ставшую классической работу [4], в которой представили теоретические основы для понимания механизмов распределения птиц в зависимости от качества местообитаний. В своем исследовании они описали первую теоретическую модель ИСР и ее зависимость от эффекта Олли. Статья Фретуэлла и Лукаса является одной из самых цитируемых в экологии (более 4000 цитирований), а предложенные в ней идеи нашли применение далеко за пределами орнитологии: от управления промысловыми популяциями рыб [5] до изучения процессов колонизации человеком различных регионов в археологии [6].
Несмотря на первоначальную критику, основанную на результатах полевых наблюдений [7; 8], свидетельствовавших в некоторых случаях об отклонении от ИСР, справедливость теоретических прогнозов теории была подтверждена многими исследователями на примере вертикального распределения зоопланктона [9; 10], выбора мест обитания северной щуки ( Esox lucius ) [11] и морских рыб [12]. Кроме того, чтобы не сложилось впечатление, что ИСР реализуется только в водной среде, приведем ссылку на работу [13], в которой было показано, что некоторые виды растений распределяют свои корни в почве пропорционально количеству доступных ресурсов (удобрений), а также исследование [14] с участием групп людей, распределявшихся между двумя источниками вознаграждения, которое также продемонстрировало соответствие прогнозам ИСР.
Небольшие нестыковки с теорией можно объяснить тем, что в основе концепции ИСР лежат несколько простых допущений, согласно которым особи популяции имеют «идеальное» представление о неоднородном ареале и могут «свободно» перемещаться в любую его точку [4; 15]. Процесс перемещения часто рассматривают как динамическую эволюционную игру в результате которой достигается равновесие, так, что никто не сможет улучшить свою приспособленность, просто перейдя на другое место (аналог равновесия Нэша) [16]. В работе [17] было показано, что для одного вида ИСР является эволюционно стабильной стратегией (ЭСС). Понятие ЭСС широко используется в эволюционной теории игр [18] для решения различных биологических проблем [19; 20] уже более пятидесяти лет с момента своего появления [21]. Это одно из актуальных направлений исследований, позволяющее описывать широкий диапазон биологических явлений - от агрессии и территориальности у животных [22] до выбора пола у растений и кооперации микроорганизмов [23]. Обобщение ИСР в качестве ЭСС для нескольких взаимодействующих видов представлено в работах [24-26].
По-видимому, первые исследования, распространившие концепцию ИСР на систему «хищник-жертва», - это работы [27-30]. В основе этих моделей лежали классические уравнения Лотки-Вольтерры [31; 32], которые дополнялись либо функциональным откликом Холлинга II типа [33], либо логистическим уравнением [34] для роста популяции жертвы. Такого рода системы являются «точечными» и не учитывают форму, размер и пространственную неоднородность среды обитания, а также миграцию в явном виде [35], при этом для перемещения особей между двумя разными по качеству участками ареала авторы используют различные предположения. Несколько позже начали интенсивно развиваться исследования ИСР для нескольких конкурирующих за общий ресурс видов, динамика которых описывается уравнениями диффузии-реакции [36-39]. В работах [40; 41] были рассмотрены модели «хищник-жертва» на основе системы уравнений диффузии-адвекции-реакции с мультифак-торным таксисом, где ИСР рассматривается как пропорциональность распределения обоих видов ресурсу жертвы в стационарном или осциллирующем режиме.
Эффект Олли представляет собой зависимость скорости роста популяции от ее плотности [42; 43]. Это явление впервые было описано Уордером Клай дом Олли, изучавшим динамику популяции золотых рыбок [44; 45]. Эффект Олли может проявляться вследствие таких факторов, как трудности нахождения партнеров для размножения, инбридинг, защита от хищников и коллективная добыча пищи [46; 47]. Выделяют слабый и сильный эффекты Олли: в первом случае скорость роста популяции снижается с уменьшением плотности, но остается положительной [48]; во втором - популяция имеет критический порог численности [49], ниже которого ее прирост становится отрицательным, что ведет к вымиранию. Из-за угрозы неблагоприятных последствий для экологии и биоразнообразия в настоящее время эффект Олли активно исследуется на уровне математического моделирования [50-52].
В данной работе рассматривается влияние сильного эффекта Олли в популяции жертвы на ИСР в системе хищник-жертва. Исследование проводится для неоднородного ареала.
Математическая модель
Основываясь на работе [53] (и полагая n = 1, m = 0, qt = 0 ) рассмотрим модель хищник-жертва с ИСР, описываемую системой обыкновенных дифференциальных уравнений:
ди д t
fi и У ^ 1
q U I 1--II p J
дv п и .
— = v —— Z. д t [ p '
где и (x, t) - популяция жертвы, а v(x, t) - хищника, p (x) - не равный нулю, неравномерно распределенный на ареале ресурс жертвы. Положительные коэф- фициенты μ характеризуют прирост, а λ – убыль популяций. Система (1) дополняется начальными условиями
u ( x , 0) = u 0 ( x ), v ( x , 0) = v 0 ( x ). (2)
Заменой u на ( u - a ) в системе (1) введем сильный эффект Олли для популяции жертвы:
ди
д t
/ \ I 1 и I
-! (и - a )l 1
I P J
д v - и — = v —— д t [ p
- X 2
где a – параметр Олли (пороговое значение) для численности популяции и ( x, t ), ниже которого она обре-
вое стационарное решение (3.2). Для стационарного состояния, отвечающего сосуществованию двух видов (выражения (1.4) и (3.5) соответственно), видно, что эффект Олли приводит к уменьшению численности популяции хищника. Заметим, что распределение жертвы в обоих случаях удовлетворяет ИСР, в то время как распределение популяции хищника пропорциональна квадрату ресурса без эффекта Олли и более сложной зависимости при его наличии. Кроме того, диапазон устойчивости стационарного решения становится меньше: левая часть неравенства (3.6) на μ a 2 p становится ближе к μ . Отметим также, что если параметр Олли равен нулю ( a = 0), то обе колонки таблицы 1 совпадут.
чена на вымирание.
Рассмотрим влияние параметра a на стационарные состояния системы (3) в сравнении с системой (1). Ниже приведена таблица 1, в которой слева представлены стационарные решения и условия их устойчивости для системы (1), без эффекта Олли, а справа – для системы (3), с сильным эффектом Олли у жертвы. Для удобства все формулы снабжены номерами с двойным индексом, где первая цифра соотносится с той системой, относительно которой получено решение.
Для поиска и исследования устойчивости стационарных решений систем (1) и (3) использовались стандартные методы теории динамических систем на фазовой плоскости [54].
Детальный анализ таблицы 1 показывает, что при наличии сильного эффекта Олли в популяции жертвы тривиальное состояние равновесия меняет свой характер от неустойчивого (без эффекта Олли) к устойчивому (с эффектом Олли). Более того, при наличии эффекта Олли, в системе (3) возникает новое, неустойчи-
Результаты математического моделирования
В данном разделе представлены результаты вычислительных экспериментов дающих представление о влиянии параметра Олли на ИСР и устойчивость стационарного решения для двух видов (3.5).
Численное решение задачи (3) с начальными условиями (2) осуществлялось с помощью комплекса программ SPECIES-25, защищенного свидетельством о государственной регистрации программ для ЭВМ [55]. Данный комплекс позволяет проводить вычислительные эксперименты для различных задач популяционной динамики (в том числе и на основе нелинейных систем диффузии-адвекции-реакции), в условиях неоднородного ареала.
На рис. 1 изображены распределения ресурса и популяций жертвы и хищника для стационарных решений (1.4) и (3.5) соответственно.
Таблица 1
Сравнение стационарных решений двух систем в отсутствии и при наличии сильного эффекта Олли у популяции жертвы
|
Система (1), без эффекта Олли |
Система (3), с сильным эффектом Олли у жертвы |
|
и = 0, v = 0 (1.1) неустойчивое стационарное решение |
и = 0, v = 0 (3.1) устойчивое стационарное решение |
|
--- |
и = a , v = 0 (3.2) неустойчивое стационарное решение |
|
и = p, v = 0 (1.2) стационарное решение устойчиво, если X 2 > р 2 (1.3) |
и = p , v = 0 (3.3) стационарное решение устойчиво, если X 2 > - 2 (3.4) |
|
и = ^ p , v = ^ f1 - ^I ^ p2 (1.4) - 2 X1 1 - 2 J - 2 стационарное решение устойчиво, если - 2 < Х2 < - 2 (1 .5) |
и = — p , v = —|1 - — || — p - a | p (3.5) - 2 X1 I - 2 Д - 2 J стационарное решение устойчиво, если - 2 ( p + a )^. < < X2 < -2 (3.6) 2 p |
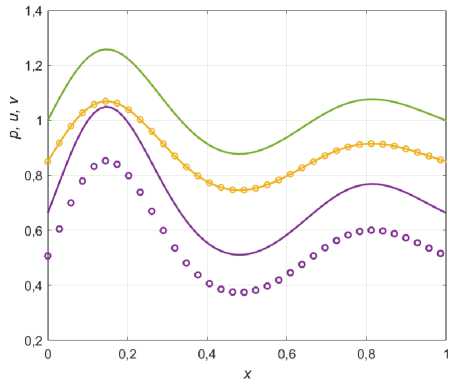
Рис. 1. Распределения ресурса (зеленый), жертвы (желтый) и хищника (фиолетовый). Сплошные линии решение (1.4), кружочки - решение (3.5) при a = 0,2 . Коэффициенты систем: Ц; = 5,2;^ = 1;ц2 = 2Д2 = 1,7 .
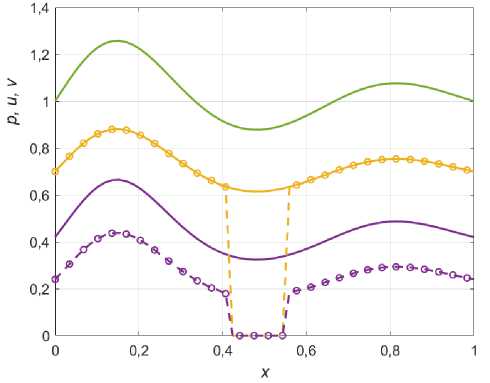
Рис. 2. Распределения ресурса (зеленый), жертвы (желтый) и хищника (фиолетовый). Сплошные линии решение (1.4), кружочки с пунктиром – решение (3.5) при значениях а = 0,3; ц = ц2 = 2; \ = 1; Х2 = 1,4 .
Как видно из графиков, распределение хищника носит ИСР-подобный характер: пропорционально квадрату ресурса для решения (1.4) и более сложной зависимости для решения (3.5). При этом, распределение жертвы пропорционально распределению ресурса (ИСР) и, что интересно, не изменяется с появлением эффекта Олли, когда а ^ 0 . Однако, при увеличении параметра Олли, для некоторых точек ареала (где ресурс p ( x ) минимален) решение (3.5) становится неустойчивым для малых возмущений системы (рис. 2).
Более детально процесс установки к равновесию для небольших (менее 10%) возмущений начальных распределений (2) от точного решения (3.5) показан на рис. 3. Видно, что совершая колебания, популяции на небольшом участке ареала достигают критических значений, что приводит к сокращению их численности на данном интервале до нуля.
При нарушении условия (1.5) в отсутствии эффекта Олли ( а = 0) система «хищник-жертва» совершает незатухающие колебания, показанные на рис. 4.
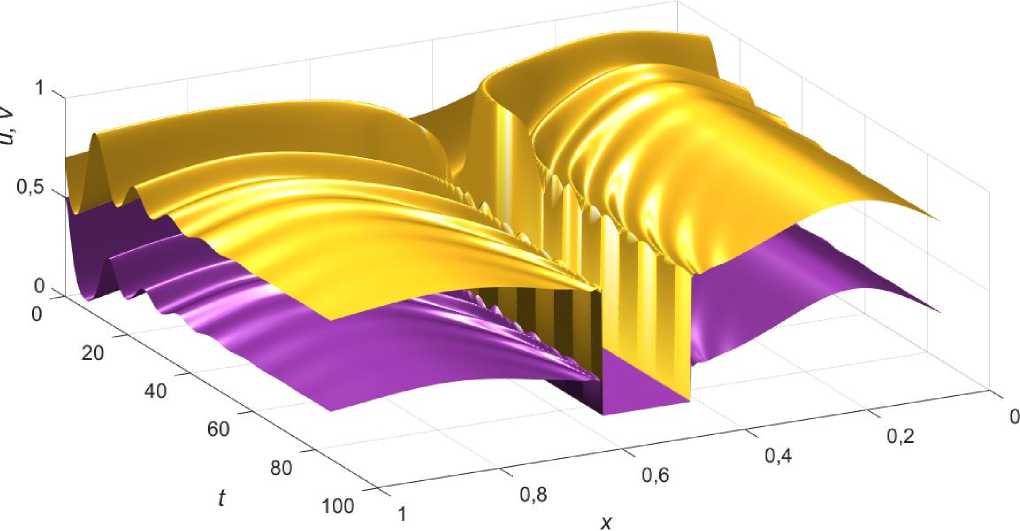
Рис. 3. Выход на стационарное распределения жертвы (желтый) и хищника (фиолетовый) во времени.
Значения коэффициентов а = 0,3; ц = ц2 = 2; \ = 1; Z2 = 1,4; u 0 ( x ) = е • u ( x ); v 0 ( x ) = e • v ( x ); e = 0,92 .
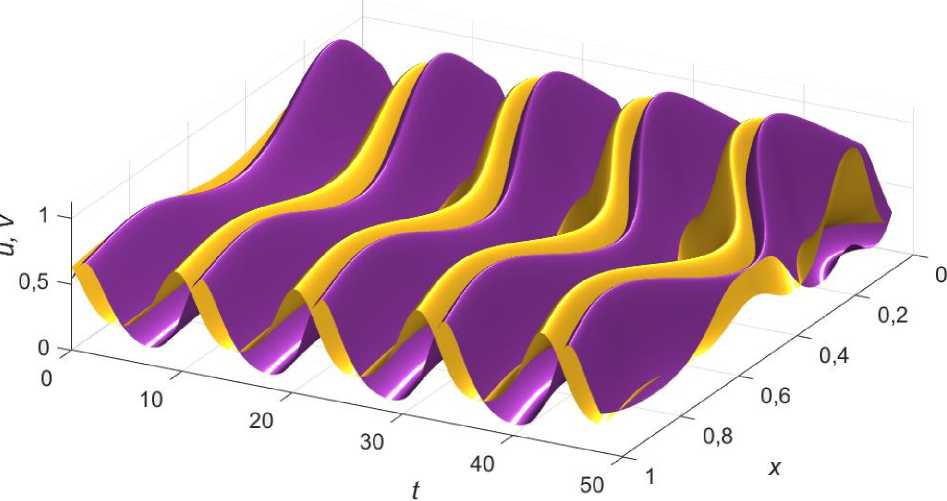
Рис. 4. Колебательный режим при нарушении условия (1.5) в системе без эффекта Олли.
Значения коэффициентов: a = 0; ^ = ц2 = 2; Z = 1,2; Z2 = 0,9; u 0( x ) = e • u ( x ); v 0( x ) = e • v ( x ); e = 1,3 .
При малых возмущениях начальных условий такой осциллирующий режим сохраняет свою устойчивость.
Наиболее интересным случаем является возникновение колебаний при локальном нарушении условия (3.6) в системе с эффектом Олли, показанное на рис.5.
Видно, что возмущение начальных условий приводит к затухающим колебаниям по краям ареала, в то время как в его центре зарождается осциллирующий режим, вызванный изменением функции ресурса по координате, в минимуме p ( x ) нарушается условие (3.6).
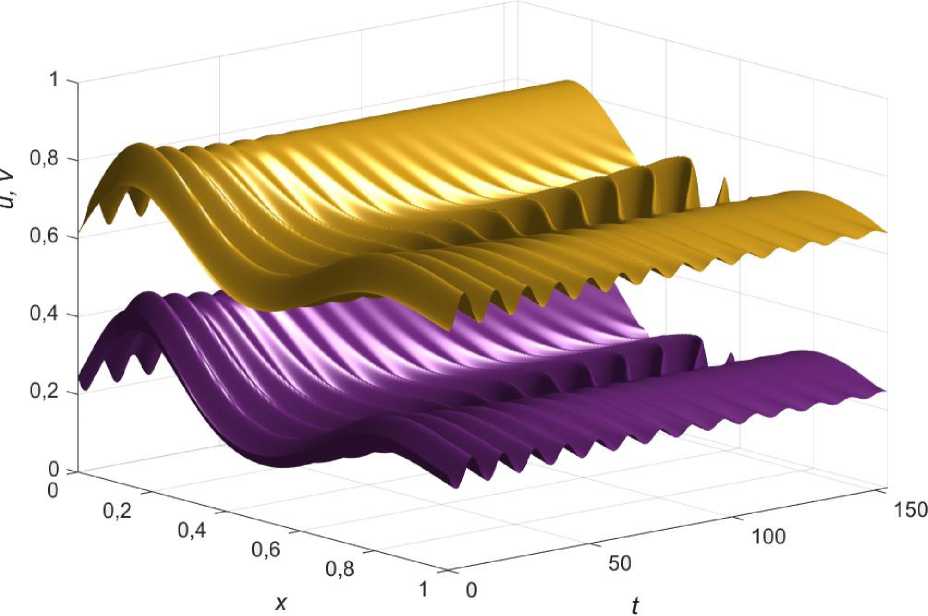
Рис. 5. Появление колебаний при локальном нарушении условия (3.6) в системе с эффектом Олли.
Значения коэффициентов: a = 0,3; Ц; = ц2 = 2; Z = 1; Z2 = 1,32; u0 ( x ) = e • u ( x ); v 0( x ) = e • v ( x ); e = 0,92
Заключение
В данном исследовании изучено влияние эффекта Олли на динамику модели «хищник–жертва» с идеальным свободным распределением (ИСР) на неоднородном ареале. Неоднородность ареала задавалась одномерной обобщенной функцией ресурса жертвы. Для наглядности, работа построена на сопоставлении двух систем: исходной – без эффекта, и модифицированной – с сильным эффектом Олли в популяции жертвы. Обе модели реализуют ИСР, позволяющее популяциям максимизировать свою приспособленность в гетерогенной среде и выстраивать эволюционно стабильную стратегию (ЭСС).
Проведенное сравнение стационарных решений, показало, что популяция жертвы всегда сохраняет ИСР, в то время как популяция хищника имеет сложные ИСР–подобные распределения, зависящие от функции ресурса и параметра Олли.
Показано, что наличие сильного эффекта Олли у жертвы приводит к смене устойчивости тривиального нулевого состояния равновесия и появлению нового неустойчивого решения без хищника, когда численность жертвы равна пороговому значению параметра Олли. При этом диапазон устойчивости стационарного состояния, отвечающего сосуществованию видов, уменьшается на величину пропорциональную параметру Олли и обратно пропорциональную ресурсу.
