Индекс фрагментации ДНК -маркер фертильности спермопродукции норок

Автор: Абрамов П.Н., Борунова С.М., Иолчиев Б.С.
Статья в выпуске: 4 т.244, 2020 года.
Бесплатный доступ
В ходе выполнения эксперимента была установлена прямая зависимость биологической полноценности спермы от белкового питания, а именно - отсутствие необходимого состава аминокислот в базовом рационе норок. Результаты исследования показывают, что в эякуляте самцов, получавших в рационе по 2 и 4 мл 15 % раствора белкового гидролизата по сравнению с контрольной группой, получавшей только основной рацион, содержание подвижных сперматозоидов больше на 17 и 15 % (p
Норка, сперма, гидролизат, дезоксирибонуклеиновая кислота, индекс фрагментации
Короткий адрес: https://sciup.org/142226083
IDR: 142226083 | УДК: 575:636.934.56 | DOI: 10.31588/2413-4201-1883-244-4-4-10
DNA fragmentation index - a marker of fertility mink sperm products
The main direction of the research was to study the morphological and functional characteristics of spermatozoa in the ejaculate of male minks and to identify the fragmentation index of nuclear and mitochondrial DNA of spermatozoa.In the course of the experiment, a direct dependence of the biological usefulness of sperm on protein nutrition was established, namely, the lack of the necessary composition of amino acids in the basic diet of minks.The results of the study showed that in the ejaculate of males who received 2 and 4 ml of a 15 % solution of protein hydrolyzate in the diet, compared with the control group, who received only the basic diet, the content of motile spermatozoa is 17 and 15 % more (p
Текст научной статьи Индекс фрагментации ДНК -маркер фертильности спермопродукции норок
Успехи современной зооветеринарной практики в вопросах лечения бесплодия сельскохозяйственных самцов-прозводителей в комплексе с применением вспомогательных репродуктивных технологий определяют острую необходимость совершенствования методов ветеринарной лабораторной диагностики по установлению этиопатогенетических механизмов развития инфертильности.
Перевод животноводства на промышленную основу спровоцировал изменение условий содержания пушных зверей в условиях доместикации, лишая их привычной естественной среды обитания.
Известно, что современные индустриальные способы содержания животных (изменение функциональных нагрузок организма, адаптационные реакции на внешние раздражители, вызывающие стресс) негативно влияют на гомеостаз и продуктивность в целом.
В таких сложившихся условиях чаще всего страдает молодняк зверей из-за несовершенства физиологического состояния организма и снижения естественной резистентности, ведущей к снижению продуктивности в пушно-меховой отрасли.
Для увеличения пушно-меховой продуктивности требуется улучшение кормовой базы, за счет оптимизации структуры рациона, использования раз- личных доступных нетрадиционных кормовых добавок, улучшающих качество корма и оказывающих положительное влияние на физиологическое состояние организма.
В настоящее время широко интегрируется в отечественную кормоинду-стрию перспективное направление по применению гидролизатов животных белков в составе базового питания. Так, белковые добавки способны оказывать активное влияние на гомеостаз организма животных, на его физиологические функции, а также репродуктивные показатели [11]. Репродуктивная функция одна из важных функций организма, которая обладает мощным инструментом в обеспечении зачатия, рождения и выхаживания зрелого жизнеспособного потомства и является наиболее чувствительным маркером, точно отражающим любое отклонение от показателей нормы в организме и напрямую зависящим от полноценного кормления, которое занимает приоритетное значение в норководстве.
В соответствии с вышесказанным, цель нашего исследования состояла в определении роли белкового питания на репродуктивные функции самцов норок, путем введения белкового гидролизата в качестве кормовой добавки в их основной рацион. Кормление обеспечивает опти- мальную потребность организма в энергии, протеине, незаменимых аминокислотах, минеральных веществах, витаминах, необходимых для нормального протекания морфофизиологических, гормональных, биохимических, биофизических процессов сперматогенеза и овогенеза в репродуктивных органах любого вида животных. Морфофункциональное строение, целостность, активность половых клеток напрямую зависит от правильного баланса белков, жиров и углеводов в базовом питании, а также от аминокислотного состава рациона, поскольку дефицит какой либо одной аминокислоты (лимитирующей) в рационе может привести к частичному неусвоению всех остальных, возникновению дисбаланса гомеостаза организма, приводящего к различным цитоатипиям гамет в эякуляте, где атипичные структуры половых клеток представлены различными морфологическими аномалиями сперматозоидов и функциональными нарушениями отдельных структурных единиц.
Трофическое обеспечение гамет зависит от каскада биохимических реакций, синтеза и катаболизма нуклеопротеидов метаболического процесса. Нуклеопротеиды поступают в организм вместе с пищей и распадаются на белки, в дальнейшем на аминокислоты в тонком отделе кишечника, а блокировка или локальное нарушение какой–либо из фаз процесса метаболизма ведет к возникновению аномальных форм.
По данным Е.Е. Брагиной (2014), в последние годы к лабораторному критерию качества сперматозоидов относят оценку структурных нарушений хромосом – это не правильная упаковка и наличие разрывов нитей ДНК в головке спермия [2]. Уже известно, что сперматозоиды с фрагментацией ДНК могут повреждать яйцеклетку, в результате чего она теряет функциональность, эмбрион не образуется, протокол оплодотворения оканчивается неудачно. Если уровень фрагментации ДНК невысок, то в некоторых случаях молодая и здоровая яйцеклетка может восстановить структуру нуклеотидной цепочки из имеющихся нуклеотидных остатков в цитотоли клетки. Считается, что окислительный стресс сперматозоидов выражает- ся в повреждении свободными радикалами их мембран, белков и ДНК. В норме радикалы нейтрализуются антиоксидантами, но при метаболических нарушениях, при воспалительных процессах, иммунных расстройствах баланс гомеостаза нарушается, активных форм кислорода становится больше, и они начинают проявлять свое негативное влияние на те или иные органы.
Чаще всего этот процесс запускается на стадии мейоза в половых железах норок и продолжается до выхода сперматозоидов из их придатка, поэтому при снижении фертильности животного организма необходимо в базовый рацион животных вводить различного рода добавки, на основе аминокислот, для регулирования белкового, липидного и углеводного метаболизма.
Материал и методы исследований. Объектом исследования были самцы норки. Для проведения исследования было сформировано три группы: I и II группа -опытные, III контрольная, при этом в каждой группе – по семь голов самцов годовалого возраста. Животным первой и второй групп к основному рациону добавляли по 2 и 4 мл 15 % раствора белкового гидролизата, соответственно, в то время как контрольные аналоги находились на обычном рационе. Введение гидролизата опытным группам в рацион начинали за два месяца до начала гона и в период него. Белковый гидролизат, полученный нами в ЗАО «Биопрогресс» [1, 3], содержал в своем составе весь спектр незаменимых аминокислот, учитываемых в пушном звероводстве. Дозу гидролизата определяли из необходимой потребности животных в аминокислотах и по фактическому содержанию ЛНАК (лимитирующие аминокислоты) в рационе. Сбор спермы проводили из влагалища самок норок, после прерванного коитуса, с помощью пипетки. Активность сперматозоидов определяли с помощью пакет программы «AndroVision®». Для оценки морфологии сперматозоидов использовали набор реагентов SpermBlue и программное обеспечение Argus-CASA. Состояние ДНК в сперматозоидах изучали методом акридин-оранжевого теста (АО- тест) с использованием флуоресцентной микроскопии. Целостность акросом сперматозоидов изучали с помощью ДИФ-Квик окрашивания. Для статистического анализа полученных данных использовали пакет программы анализа данных программы IBM SPSS Statistics 23.0 («IBM Corp.», США).
Результаты исследований. Подвижность сперматозоидов в опытной и контрольной группе существенно отличались. В I и II опытных группах содержание подвижных сперматозоидов составило соответственно 62±1,6 и 61±0,9 в контрольной группе 53±1,9 % (Таблица 1). Между опытными группами по содержанию прогрессивно подвижных сперматозоидов ста- тистическое значимое различие не установлено. Первая опытная группа по данному показателю превосходила контрольной группу на 17 % (p≤0,01) и вторая на 15 % (p≤0,01). Анализ траектории подвижности активных сперматозоидов показывает, что движение сперматозоидов норок отличается по характеру движения. Сперматозоиды двигаются прогрессивно криволинейно.
В эякуляте контрольной группы доля сперматозоидов с непрогрессивным движением составила более 18 %, что в 1,9 больше по сравнению со второй опытной группой. Разница между опытными группами и контрольной группой оказалась статистическая достоверная при p≤0,001.
Таблица 1 – Качество спермы норок в зависимости от состава рациона
|
Показатель |
Группы |
||
|
I опытная |
II опытная |
Контрольная |
|
|
Содержание сперматозоидов, %: прогрессивно-подвижные |
62,0±1,6** |
61,0±0,9** |
53,0±1,9 |
|
непрогрессивно-подвижные |
11,4±0,6*** |
9,6±0,7*** |
18,3±0,3 |
|
Неподвижные |
26,6±1,4 |
29,4±1,6 |
28,7±0,8 |
|
Морфологические аномалии, % |
12,7±0,6** |
13,5±0,9** |
19,3±0,8 |
|
Индекс фрагментации ДНК, % |
2,0±0,1*** |
2,1 ±0,1*** |
30±0,8 |
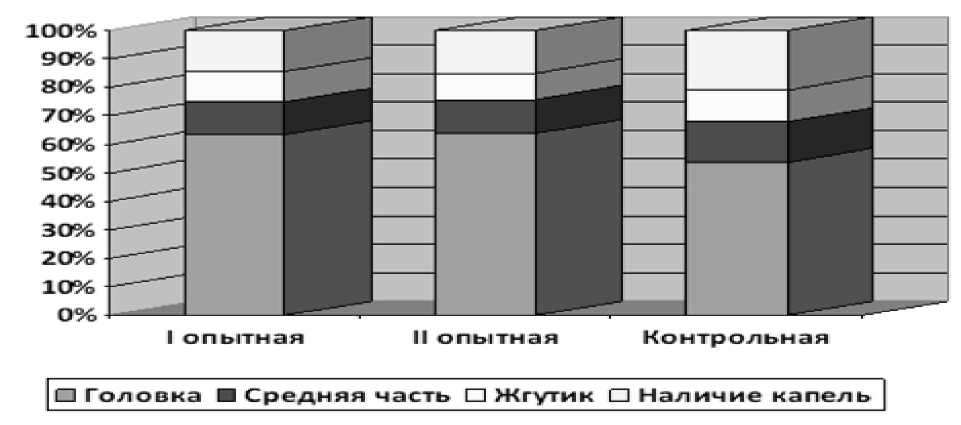
Рисунок 1 – Частота встречаемости патологии в отдельных сегментах сперматозоидов
Наивысшая частота аномалии морфологии в эякуляте наблюдалась в контрольной группе. Содержание сперматозоидов с различными морфологическими нарушениями в данной группе в среднем составило 19,3 % и в зависимости от индивидуальных особенностей самца варьиро- вало от 14 до 26 %.
Исследуемые группы отличались по частоте встречаемости аномалии отдельных сегментов сперматозоидов. При этом сохранялась общая закономерность, когда наиболее часто встречались патологии в головке и средней части сперматозоидов
(Рисунок 1).
В исследованиях, проведенных нами, были зафиксированы следующие патологические формы сперматозоидов норок: дефекты головок - присутствие образцов безъядерных, шаровидных и конусовидных форм. При этом в головках аномальных спермиев наблюдали изменение локализации акросомы и даже её отсутствие в контрольной группе. Дефекты шейки и средней части гамет в образцах были представлены сперматозоидами во всех экспериментальных группах, наличие спиралевидно сложенного жгутика в контрольной группе.
В 70 % образцах, полученных от самцов контрольной группы, встречались сперматозоиды с фрагментацией ядерной и митохондриальной ДНК. Индекс фрагментации ДНК в сперматозоидах самцов контрольной группы в среднем составил 30 %, у первой опытной группы в полученных эякулятах индекс фрагментации ДНК в среднем составил 2 %, во второй опытной группе 2,1 %. Разница между контрольной и опытными группами была статистически значимой при p<0,001. Часто встречалась фрагментация ДНК в хроматине сперматозоидов контрольной группы норок (Рисунок 2).
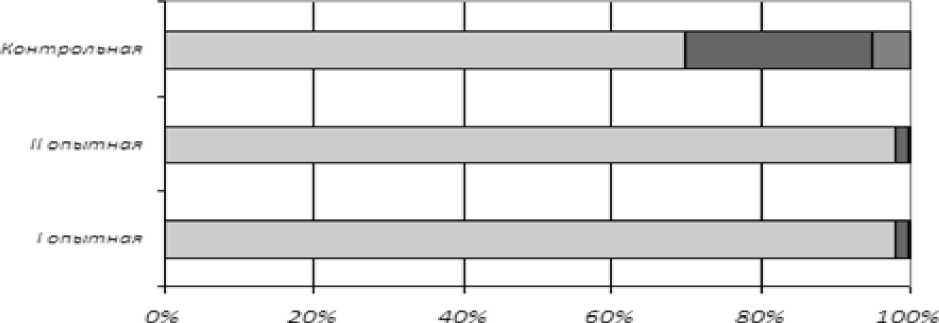
-
□ и ДНК ■ яДНК ■ вцДНК |
Рисунок 2 - Степень фрагментации ДНК в сперматозоидах иДНК - интактная ДНК; яДНК -ядерная ДНК; мДНК - митохондриальная ДНК
При исследовании образцов спер-мопродукции самцов-норок по показателю индекса фрагментации ДНК было зафиксировано следующее: образцы спермопро-дукции самцов–производителей контрольной группы имели структурные нарушения как в яДНК (головке сперматозоидов), так и в мДНК (средняя часть). Вероятно, это обстоятельство напрямую коррелирует с отсутствием недостаточного количества в базовом рационе аминокислот, участвующих в синтезе нуклеотидов и протаминов. В спермопродукции самцов-производителей норок опытной группы подобного рода деструктивные изменения не были обнаружены. Основную массу хроматина представляют белки гистоны. Гистоны - нуклеотидные белки, активно участвующие в переписывании информации с гена на информационную РНК, а также в репарации поврежденных локусов цепи ДНК; от плотности расположения гистонов в активно экспрессирующихся участках генома зависит и интенсивность экспрессии генов. Так, в норме хроматин зрелых сперматозоидов плотно упакован, чтобы защитить ДНК от потенциальных повреждающих воздействий во время транзита его по органам репродуктивной системы. Высокая плотность гистонов формирует «закрытую структуру» хроматина, что приводит к инактивации генов; если хроматин конденсирован, то гены находятся в инактивированном состоянии, если хроматин открыт (декондесирован) и активен, то это определяет активацию экс- прессии генов. В свою очередь, динамика состояния хроматина определяется и контролируются такими обратимыми процессами, как метилирование ДНК и модификация гистонов.
Сегодня уже известно, что «выключение» генов осуществляется при помощи метилирования ДНК (прикрепления к цитозиновым основаниям ДНК метильной группы СН 3 ). «Включение» происходит за счет ацетилирования гистонов (белков в составе хроматина, необходимых для сборки и упаковки ДНК); в результате плотного контакта ДНК и гистонов в хроматине половой клетки, происходит блокирование концевых участков «хвостов» ДНК (ограничивает свободу вращения концевых нитей), что препятствует специфическим ферментам доступ к определенным локусам ДНК при сканировании наследственной информации.
От того, как цепочка ДНК упаковывается в ядре половой клетки, зависит и количество того или иного белка, который синтезируется в клетке на нужды ее существования, а выявленные цитопатологии в половых клетках, при получении данных по проведенному эксперименту, яркое тому подтверждение. Это согласуется с данными зарубежной литературы Evenson D.P., Larson K.L., Jost L.K., (2002), Agarwal A., Said T.M. (2003), Erenpreiss J., Spano M., Erenpreisa J. (2006), Kempisty B., Depa-Martynow M., Lianeri M. (2007) [4, 7, 8, 9]. Возникновение цитопатологий в спермо-продукции животных характеризуется такими нарушениями как нарушение физической целостности молекулы ДНК в виде разрыва одной или обеих полинуклеотид-ных цепей (фрагментация); дефектов ядер-ных белков, препятствующих замене гистонов на протамины с последующим уплотнением ДНК; нарушением пространственной третичной структуры хроматина.
При движении спермия по половым путям самки (подвижность гаметы сохраняется в анаэробных условиях) включается альтернативный механизм катабализма – гликолиз, в ходе которого и формируются молекулы АТФ, тем самым поддерживается двигательная активность спермия. Активация подвижности сперматозоидов за- висит от циклического аденозинмонофосфата (АМФ), производного АТФ, который, будучи универсальным регулятором метаболических реакций, активизирует цАМФ – зависимую протеинкиназу. Протеинки-наза фосфорилирует белки, а каталитические субъединицы протеинкиназ способны пересекать ядерные мембраны и фосфорилировать ядерные белки – гистоны, тем самым, регулировать генную активность половых клеток.
Полученные данные в ходе проведения эксперимента по выявлению и обнаружению фрагментированных участков ДНК в спермопродукции норок контрольной группы, не получавших в базовом рационе белковый гидролизат, позволяют нам комментировать данную ситуацию с позиции недополучения ими соответствующего белкового питания, что и отражается в процессах фрагментации цепей ДНК спермиев.
Высокое содержание оксидантов в ее окружении приводит к мутациям в мДНК с частотой в 10-20 раз большей, чем в яДНК. При репликации образуется трехнитчатая мДНК (промежуточное звено), которая очень восприимчива к делециям [13, 14]. Точечные мутации и делеция фрагментов мДНК приводят к патологии митохондриальной дыхательной дисфункции, что является одной из причин патологий спермий [5, 10, 15]. Исследования ультраструктур сперматозоидов показывают, что мутация ДНК, а также другие повреждения митохондрий сперматозоидов сопровождаются снижением их активности [6, 11]. Полученные нами данные согласуются с данными других авторов.
Белковый гидролизат, а именно аминокислотный триплет – гистидин, треонин, триптофан, не только вероятно перезапустил метаболизм гомеостаза организма норок в целом, но и выступил в роли донора по образованию заменимых аминокислот и выравниванию их по количеству: так гистидин является донором глутаминовой кислоты, нарушение метаболизма гистидина в тканях животного отражается в снижении концентрации гистидина в эритроцитах, тем самым увеличивается его содержание в сыворотке крови. Иниции- руются эти процессы тем, что тормозится реакция дезаминирования и усиливается реакция декарбоксилирования, при этом образуется биологически активное вещество – гистамин – естественный ингибитор перекисного окисления липидов.
Введение гистидина позволяет ограничить образование маланового диальдигида в тканях, приводящего к разрушению митохондрий и угнетающего процесс окислительного фосфорилирования.
Триптофан вместе с лизином образует трипептид (лизин-триптофан-лизин), который исправляет ошибки, возникающие при удвоении ДНК и возникновении различных поломок (фрагментация) в ее нитях. При нехватке этой незаменимой аминокислоты происходит выпадение шерсти у животных и нарушатся функция половых желез.
Триплет незаменимых аминокислот белкового гидролизата, поступающий путем применения его в составе базового рациона опытных групп самцов норок, инициировал, по-видимому, работу необходимых ферментов, поддерживающих равновесие концентрации молекул АТФ в спер-мии.
Созданное биохимическое равновесие метаболических процессов спермия позволяет избежать потребности в диффузии его на большие расстояния по половым путям. Отсутствие аминокислотного обогащения в базовом рационе норок приводит к морфофункциональным отклонениям половых клеток, что может отражаться не только на количестве коитусов самцов–производителей, но и на развитии эмбрионов, а также на рождении здорового приплода
Заключение. Таким образом, результаты исследования показывают, что добавление в рацион самцов норок 2 мл 15 % раствора белкового гидролизата улучшает биологическую полноценность сперматозоидов, снижает индекс фрагментации ядерной и митохондриальной ДНК, снижает частоту встречаемости патологии сперматозоидов и улучшает их активность.
В результате исследований отмечено, что между первой опытной (самцы по- лучали основной рацион + 2 мл 15 % раствора белкового гидролизата) и второй опытной группами (основной рацион + 4 мл 15 % раствора белкового гидролизата) статистическое значимое различие не установлено, следовательно, экономически эффективно добавление в рацион 2 мл белкового гидролизата.
DNA FRAGMENTATION INDEX - A MARKER OF FERTILITY
Список литературы Индекс фрагментации ДНК -маркер фертильности спермопродукции норок
- Абрамов, П.Н. Разработка промышленного способа получения белкового гидролизата тушек норок / П.Н. Абрамов, Р.В. Рогов // Вопросы ветеринарии и ветеринарной биологии: Сборник научных трудов молодых ученых МГАВМиБ. -2011. - С. 121-123.
- Брагина, Е.Е. Протокол проведения спермиологического исследования / Е.Е. Брагина // Андрология и генитальная хирургия. - 2014. - № 1. - С. 15-24.
- Фролова, М.А. Получение промышленной партии белкового гидролизата из тушек норок и изучение его токсичности / М.А. Фролова, А.Я. Самуйленко, A. И. Албулов [и др.] // Известия Самарского научного центра РАН. - Самара. - 2011. - Т. 13. - № 5 (3). - С. 207-209.
- Agarwal, A. Role of sperm chromatin abnormalities and DNA damage in male infertility / A. Agarwal, T.M. Said // Hum Reprod Update. - 2003. - № 9 (4). P. 331-45.
- Amaral, A. Mitochondria functionality and sperm quality / A. Amaral, B. Lourenco, M. Marques [et. al] // Reproduction. - 2013. - Vol. 146 (5). - P. 163-174.
- Borunova, S.M. Astenozoospermiya u bykov-proizvoditeley / S.M. Borunova, B.S. Iolchiev, O.E. Badmaev [et. al] // Veteri-nariya, zootekhniya i biotekhnologiya. -2017. - № 11. - P. 57-64.
- Erenpreiss, J. Sperm chromatin structure and male fertility: biological and clinical aspects / J. Erenpreiss, M. Spano, J. Erenpreisa [et al.] // Asian J Androl. - 2006 - Vol. 8 (1). - P. 11-29.
- Evenson, D.P. Sperm chromatin structure assay: its clinical use for detecting sperm DNA fragmentation in male infertility and comparisons with other techniques / D.P. Evenson, K.L. Larson, L.K. Jost // J. Androl. - 2002. - Vol. 23. - P. 25-43.
- Kempisty, B. Evaluation of protamines 1 and 2 transcript contents in spermatozoa from asthenozoospermic men / B. Kempisty, M. Depa-Martynow, M. Lianeri [et al.] // Folia Histochem Cytobiol. - 2007. Vol. 45. - P.109-13.
- Kidun, K.A. Mitokhondrialnaya disfunktsiya spermatozoidov v patogenez epatospermiy priokislitelnomstresse (obzorliteratury) / K.A. Kidun, T.S. Ugolnik // Problemyzdorovya i ekologii. - 2013. -№ 2 (36). - P. 20-24.
- Pelliccione, F. Altered ultrastructure of mitochondrial membranes is strongly associated with unexplained asthenozoo-spermia / F. Pelliccione, A. Micillo, G. Cordeschi [et al.] // J. Fertility and Sterility. - 2011. - Vol. 95 (2). - P. 641-646.
- Rutherfurd-Markwick, K. Bioactive peptides derived from food / K. Rutherfurd-Markwick, P. Moughan // Journal of AOAC International. - 2005. - № 88 (3). -P. 955-966.
- Shamsi M.B., et al. Mitochondrial DNA mutations in etiopathogenesis of male infertility // Indian Journal of Urology. -2008. - Vol. 24 (2). - P. 150-154.
- Shu-Huei, K., Mitochondrial deoxyribonucleic acid 4977-bp deletion is associated with diminished fertility and motility of human sperm / K. Shu-Huei, Ch. Hsiang-Tai, W Yau-Huei // Biology of Reproduction. -1995. - Vol. 52 (4). - P. 729-736.
- St. John, J.C. The impact of mitochondrial genetics on male infertility / J.C. John, R.P. Jokhi, C.L. Barratt // Int. J. Androl. - 2005. - Vol. 28 (2). - P. 65-73.
