Индуцирование антиоксидантных ферментов в растениях гороха избытком хлористого аммония

Автор: Мурзаева С.В.
Журнал: Известия Самарского научного центра Российской академии наук @izvestiya-ssc
Рубрика: Проблемы прикладной экологии
Статья в выпуске: 2 т.2, 2000 года.
Бесплатный доступ
Исследовали действие хлористого аммония (10-6 10-2 М) на горох при добавлении в среду выращивания. Увеличение концентрации этой соли приводило к замедлению роста растений, уменьшению содержания белка и хлорофилла в листьях и снижению скорости синтеза АТФ в изолированных хлоропластах. При избытке аммония (2,5 x 10-2 M) и длительном действии меньших концентраций (10-6 -10-4 М) увеличивалась активность ферментов супероксиддисмутазы, каталазы и пероксидазы в листьях и корнях. Сравнение растворимых (цитозольных) и труднорастворимых (мембранных) белковых фракций, а также субклеточных структур, выделенных из проростков, показало активирование ферментов хлористым аммонием преимущественно в мембранных структурах ядрах, субчастицах, митохондриях и хлоропластах. Обсуждается активирование антиоксидантных ферментов в растениях гороха в связи с мембранотропным действием хлористого аммония. Сделан вывод об уси лении окислительной активности пероксидазы при токсичном избытке соли.
Короткий адрес: https://sciup.org/148197612
IDR: 148197612
The induction of antioxidative enzymes in plants (Pisum sativum) by excess ammonium-chloride
Investigated action of ammonium-chloride (10-6-10-2 М) on peas at addition on medium of cultivation. The augmentation of concentration of this salt result ined to retardation of growth plants, decrease of the contents of protein and of chlorophyll in leaves and depression of rate of synthesis ATP in the isolated chloroplasts. At excess of ammonium (2,5 x 10-2 M) and long action of smaller concentrations (10-6 -10-4 М) the activity of enzymes of a superoxide dismutase, catalase and peroxidase in leaves and roots was enlarged. The comparison soluble (cytosoling) and hardsoluble (membrane-bound) proteinaceous fractions, and also subcellular of fragments, secured of young growth peas, has shown activating enzymes by ammonium-chloride mainly in membranous structures (nuclei, subparticles, mitochondrions and chloroplasts). The activation antioxidative of enzymes in plants of peas is discussed in connection with by membranotropism action of ammonium-chloride. The conclusion about intensifying of oxidative activity of peroxidase is made at toxicity excess salt.
Текст научной статьи Индуцирование антиоксидантных ферментов в растениях гороха избытком хлористого аммония
Институт экологии Волжского бассейна РАН, г. Тольятти
Исследовали действие хлористого аммония (10-6 -10-2 М) на горох при добавлении в среду выращивания. Увеличение концентрации этой соли приводило к замедлению роста растений, уменьшению содержания белка и хлорофилла в листьях и снижению скорости синтеза АТФ в изолированных хлоропластах. При избытке аммония (2,5 x 10-2 M) и длительном действии меньших концентраций (10-6 -10-4 М) увеличивалась активность ферментов супероксиддисмутазы, каталазы и пероксидазы в листьях и корнях. Сравнение растворимых (цитозольных) и труднорастворимых (мембранных) белковых фракций, а также субклеточных структур, выделенных из проростков, показало активирование ферментов хлористым аммонием преимущественно в мембранных структурах - ядрах, субчастицах, митохондриях и хлоропластах.
Обсуждается активирование антиоксидантных ферментов в растениях гороха в связи с мембранотропным действием хлористого аммония. Сделан вывод об усилении окислительной активности пероксидазы при токсичном избытке соли.
Загрязнение окружающей среды делает необходимым изучение трансформации вредных соединений (ксенобиотиков) в растениях, выявление первичных мишеней воздействия, метаболических и детоксикационных путей их в живой клетке. К настоящему времени накоплен достаточно обширный материал, показывающий неспецифическое действие любых ксенобиотиков на клетку, проявляющуюся в активировании перекисного окисления липидов (ПОЛ) в клеточных мембранах, образовании активных форм кислорода (АФК), индуцирующих свободно-радикальные механизмы и провоцирующих окислительные стрессы [1-2].
К АФК относят: очень токсичные - гидроксильный радикал (ОН,), гидроксид ион радикал (ОН") и супероксидрадикал (О2") и менее токсичные - гидроксид ион кислорода (НО2-) и перекись водорода (Н2О2) [3]. В живых клетках, в том числе и растительных, существуют механизмы противостоящие окислительному стрессу, так называемые антиоксидантные системы, включающие ферменты и низкомолекулярные соединения. Ключевыми ферментами можно рассматривать супе-роксиддисмутазу, каталазу и пероксидазу связанных последовательно механизмом каталитического действия. Супероксиддисмутаза (СОД), КФ 1.15.11, осуществляет дисмутацию супероксидного радикала до перекиси водорода и кислорода, каталаза (КАТ), КФ 1.11.1.6, разрушает перекись водорода на воду и кислород [4], а пероксидаза (ПО), КФ 1.11.1.7, окисляет широкий спектр восстановителей с участием Н,О, [5]. Совместное действие этих ферментов с низкомолекулярными антиоксидантами, такими как а- токоферол (витамин Е), восстановленный глутатион, аскорбат и др., обеспечивает защиту от АФК в живых организмах [6,7].
В литературе сообщается об активации ферментов - СОД и КАТ [8] или ПО [9-11] при увеличении адаптационных возможностей растений в различных неблагоприятных условиях, в том числе и при действии ксенобиотиков [12], однако не представлены данные об одновременной активации всех трех ферментов при действии токсичных концентраций загрязнителей. Сопоставление этих активностей в растениях в различных неблагоприятных условиях помогло бы разрешить вопрос об использовании их в биотестировании, который дискутируется в специальной литературе [13]. С другой стороны, активация СОД и ПО может указывать на развитие окислительного процесса, особенно в случае фермента пероксидазы. Каталитический механизм действия этого фермента не является однозначно антиоксидативным [10,14]. В последнее время ПО приписывают не только защитную роль от АФК, но и окислителя при детоксикации чужеродных соединений [15]. Нас этот вопрос заинтересовал в связи с выявлением первичных мишеней воздействия ксенобиотиков на растения и разработкой биохимических тестов на загрязнение окружающей среды.
С этой целью в настоящей работе исследовали активность СОД, КАТ и ПО в горохе при добавлении в среду выращивания хлористого аммония в широком диапазоне концентраций, включая токсические. Предстояло оценить изменения активности каждого фермента в разных частях растения - листьях и корнях, в субклеточных мембранных структурах и фракциях растворимых (цитозольных) и мембранных белков.
Методика
Горох (Pisum sativum L.) сорт "Неосыпа-ющийся" выращивали в почвенной или водной культуре на среде Чеснокова-Базыриной, при естественном освещении и температуре 22-27 ° С. Полив и смену среды гидропоники проводили два раза в неделю. Хлористый аммоний (10-6 -10-2 М) добавляли в среду (или в почву при поливе) в течение всей вегетации. Концентрация соли в каждом опыте различалась десятикратно, кроме того испытаны концентрации 2,5х10-3 и 2,5х10-2 М. Опыты с каждой концентрацией проводили в 2-х или 4-х кратной повторности. Растительный материал для выделения белков и субклеточных фракций отбирали через 1214, 27, 35 и 70 сут.
Выделение белковых фракций. Навеску растительного материала, 1-5 г, растирали в ступке с кварцевым песком в 50 мл среды, содержащей 0,8 М сахарозу 0,05 М Трис -HCl буфер, pH 8,2. Гомогенат фильтровали через четыре слоя капрона и центрифугировали 30 мин при 17000 g. В супернатанте, предварительно пропущенном через обеззоленный бумажный фильтр, определяли активность растворимых цитозольных ферментов. Труднорастворимые мембранные белки получали из осадка. Его обрабатывали охлажденным до -180 С абсолютным ацетоном (объемное отношение осадка к ацетону 1: 9), выдержи вали в холодильнике 2 ч, затем, после осаждения на центрифуге (режим указан выше), промывали охлажденным 80% ацетоном до полного удаления пигментов. После промывки осадок подсушивали (под феном), и из него экстрагировали мембранные белки 0,05 М Трис - HCl буфером, рН 8,2. Полученные грубые экстракты цитозольных и мембранных белков подвергали частичной очистке. Вначале их высаливали сульфатом аммония до 70 % насыщения, затем обессоливали на колонке с сефадексом g-25, уравновешенной 0,05 М Трис - НCl буфером, рН 8,2. Удержание белка в белковых фракциях определяли по методу Bradford [16].
Выделение субклеточных фракций. Хлоропласты получали из листьев, по методу West, Wischich [17]. Среда для выделения содержала: 0,4 М сахарозу 0,01 М MgCl2,0,01 М этилендиаминтетрацетат (ЭДТА) и 0,5 % бычий сывороточный альбумин (БСА) в 0,03 М Т рис- HCl буфере, рН 7,4. Среда для суспендирования хлоропластов: 0,4 М сахароза и 0,5 % БСА, рН 7,4.
Реакции фотофосфорилирования проводили в термостатируемой ячейке при 250С и освещении белым светом интенсивностью ФАР 50 Вт на м2. Количество синтезируемого АТФ определяли биолюминесцентным методом, согласно работе [18]. Реакционная среда для фотофосфорилирования: 4 мМ АДФ, 2 мМ KH 2 PO 4 , 2 мМ MgCl2, 0,17 мМ метилвиологен (МВ), 0,03 мМ Феназинметосульфат (ФМС), 0,05 мМ диурон (ДХММ), 20 мкг хлорофилла на 1,5 мл реакционной среды в 30 мМ Трис-HCl буфере рН 7,9.
Ядра и пластиды (объединенная фракция), а также митохондрии и микрочастицы выделяли из корней. Промытые корни (5 г) измельчали ножницами, а затем - в гомогенизаторе 30 сек при 1500 об/мин в 75мл среды, содержащей 0,3 М сахарозу, 0,015 М фосфатный и 0,02 М Трис -HCl буферы, рН 7,3. Гомогенат фильтровали через четыре слоя капрона и центрифугировали 30 мин при 15000 g, осаждая общую мембранную фракцию органелл и микрочастиц. Эту фракцию суспендировали в указанной выше среде, после чего осаждали дифференциальным центрифугированием ядра + пластиды (10
мин при 1300 g), затем митохондрии (10 мин при 8000 g) [19]. Оставшийся супернатант, содержащий пероксисомы и другие субклеточные структуры использовали как источник субчастиц [20]. Осажденные органеллы (ядра + пластиды, митохондрии) ресуспенди-ровали в среде, содержащей 0,3 М сахарозу и 0,05 М Трис-HCl буфер, рН 8,2. В суспензию выделенных органелл и в супернатант с субчастицами, а также к хлоропластам, добавляли Тритон Х-100 до конечной концентрации 0,1%, выдерживали 1 ч при 40С и определяли активности антиоксидантных ферментов в полученных экстрактах.
Определение активностей ферментов. Активность СОД определяли спектрофотометрически по ингибированию восстановления нитросинего тетразолия (по увеличению поглощения при 560 нм) в реакционной смеси с ФМС и НАДН [21]. За единицу активности фермента принято его количество, ингибирующее восстановление нитросинего тетразолия на 50 % , принятое за 1 относительную единицу в расчете на мг белка в пробе.
Активность КАТ определяли полярографическим методом с помощью электрода Кларка по скорости выделения кислорода при разложении Н2О2 [18]. Пероксидазную активность измеряли спектрофотометрически по скорости окисления бензидина перекисью водорода методом Бояркина [22]. За единицу активности этих ферментов принято их количество, разлагавшее 1 мкмоль Н2О2 в мин на мг белка в пробе.
В работе использовали реактивы отечественного производства марки Х.Ч. и О.С.Ч., кроме: БСА, НАДН и ЭДТА фирмы "Reanal"
(Венгрия), Тритон Х-100 фирмы "Merck" (ФРГ), Сефадекс g-25 средний и ФМС фирмы "Serva" (Швеция).
Результаты
Выращивание гороха в присутствии хлористого аммония в диапазоне испытанных концентраций 10-6-10-2 М показывало заметное влияние на рост и развитие растений в течение вегетационного периода. Действие меньших концентраций - 10-6-10-4 M, проявлялось в замедлении роста надземных частей и корней, которое отмечалось через 3-4 недели выращивания. По мере развития растений наблюдали утолщение главного осевого корня, скручивание и пожелтение листьев, опадание цветковых завязей. В водной культуре такие нарушения появлялись раньше, чем в почвенной.
Высокие концентрации NH 4 C1, 10-3-10-2 М, оказывали губительное действие на горох в первые недели выращивания. Двухнедельные проростки гороха, выращенные при указанных концентрациях аммония в водной культуре, отставали в росте в 1,5 - 3 раза и содержали меньше белка и хлорофилла в листьях, по сравнению с контрольным вариантом, табл.1. Надземная часть у таких растений желтела и погибала к концу третьей, а корни - к концу четвертой недели. В почвенной культуре растения гибли через 1,5-2 месяца.
Как оказалось, морфометрические повреждения в горохе, вызванные высокими концентрациями аммония, сопровождались усилением активностей антиоксидантных ферментов в листьях и корнях гороха. В табл.2.
Таблица 1. Влияние высоких концентраций NH 4 CI на рост гороха, содержание белка и хлорофилла в листьях
|
Концентрация NH4C1, М |
Высота побегов, см |
Содержание (мг на г сырой массы) белок хлорофилл |
|
|
0 |
20 ± 2 |
14 ± 2 |
1,1 ± 0,1 |
|
2,5x10-3 |
13 ± 2 |
12,5 ± 1,5 |
1,0 ± 0,2 |
|
2,5x10-2 |
6 ± 2 |
11,5 ± 2,5 |
0,3 ± 0,03 |
Примечание. Здесь и в табл.2,3 и 4 водная культура, возраст 14 сут. Представлены средние данные 4 опытов; ± стандартное отклонение от среднего,.
Таблица 2. Активность* антиоксидантных ферментов в горохе, выращенном при 2,5х10-2 М NH 4 CI
|
Ферменты |
Контроль (-NH 4 CI) |
Опыт (+NH4C1) |
|
Из листьев сод |
58 ± 6 |
178 ± 15 |
|
КАТ |
3,4 ± 0,4 |
4,2 ± 0,3 |
|
по |
400 ± 60 |
2700 ± 150 |
|
Из корней сод ** |
222 ± 32 |
337 ± 48 |
|
КАТ |
2,8 ± 0,3 |
5,0 ± 0,5 |
|
по ** |
1070 ± 150 |
15900 ± 230 |
Примечание. * Суммарная активность, цитозольных и мембранных белков частично очищенных, ** -в грубых неочищенных белковых фракциях. Активность ферментов здесь и в табл. 3, 4, 5: СОД -относительные ед., КАТ и ПО - мкмоль Н2О2/ (мин/ мг белка).
приведены данные по действию одной концентрации, другая, меньшая (2,5х10-3 М), вызывала подобные эффекты, но при более продолжительном выращивании (3-4 недели). Активности СОД и ПО, отмеченные в таблице знаком (**), были получены в неочищенных белковых фракциях из корней. Очистка, как правило, уменьшала активность указанных ферментов. Причиной тому могла быть неустойчивость ферментов при длительной процедуре выделения. Несмотря на варьирование активностей, действие аммония отчетливо проявлялось в активировании всех трех ферментов. В среднем, под влиянием NH 4 C1 в листьях увеличивалась активность СОД в 3; КАТ - в 1,2 и ПО - в 6,7 раз; в корнях, соответственно - в 1,5; 1,8 и 15 раз.
В некоторых опытах в корнях проростков активность ПО увеличивалась при высоких концентрациях аммония в 30 и более раз.
Концентрации NH 4 C1 10 " 6-10-4 M, тоже активировали ферменты, но в меньшей степени и при продолжительном воздействии. Подробные данные вегетационных опытов не приводятся, но отметим, что после 4-5 недель выращивания гороха в почвенной культуре активность СОД увеличивалась в среднем на 30-50 %, КАТ - на 10-30 %, ПО -на 50-80 %, причем, СОД больше активировалась в листьях, а ПО - в корнях. С возрастом и появлением внешних признаков повреждений растений активность СОД и ПО, в основном, увеличивалась, а каталазная активность, как правило, уменьшалась.
Таблица 3. Активность антиоксидантных ферментов в мембранных и цитозольных белковых фракциях, выделенных из гороха, выращенного при избытке хлористого аммония
|
Условия выращивания |
А К Т И В Н О С Т Ь |
|||||
|
сод |
КАТ |
по |
||||
|
м |
Ц |
м |
ц |
м |
ц |
|
|
Контроль 2,5х10-2 М NH 4 C1 |
В листьях |
|||||
|
46 158 |
11 18 |
2,1 2,4 |
1,3 2,2 |
40 100 |
200 400 |
|
|
Контроль 2,5х10-2 М NH 4 C1 |
В корнях |
|||||
|
172 287 |
50 50 |
1,9 3,7 |
0,9 1,3 |
3700 7400 |
7000 8500 |
|
Примечание. Активность ферментов в неочищенных белковых фракциях: м - мембранные, ц -цитозольные. Здесь и в табл. 4,5 представлены средние данные 2 -х опытов, стандартное отклонение ± 10-15%.
Таблица 4. Активность антиоксидантных ферментов в субклеточных структурах, выделенных из корней гороха, выращенного при избытке хлористого аммония
|
Субклеточные структуры |
СОД |
КАТ |
ПО |
|||
|
+ |
- |
+ |
- |
+ |
||
|
Ядра, пластиды |
0,1 |
1,5 |
0,04 |
0,4 |
240 |
470 |
|
Митохондрии |
0,6 |
2,4 |
0,1 |
0,2 |
50 |
90 |
|
Субчастицы |
3,3 |
8,7 |
0,2 |
1,2 |
500 |
1000 |
Примечание. Знаки: - или + указывают на отсутствие и добавление 2,5x10 '2 M NH 4 CI в среду выращивания гороха.
Локализацию воздействия аммония на растения можно было проследить по изменению активностей исследуемых ферментов в мембранных и цитозольных белковых фракциях, в табл.3 представлены такие данные. В контроле, в листьях и корнях гороха активности СОД и КАТ в мембранных белках были выше, чем в цитозольных. Пероксидазная активность в цитозоле, наоборот, превышала мембранную. Под влиянием NH 4 C1 активировались как мембранные, так и цитозольные ферменты, однако отношение - мембранные/ цитозольные, в целом растении возрастало для СОД в 2-2,9 раза (преимущественно в листьях), а ПО - в 1,3-1,7 раза (больше в корнях). Активность КАТ листьев сильнее увеличивалась в цитозоле, уменьшая отношение - мембранная/цитозольная активность, но в корнях оно увеличивалось в 1,4 раза. Таким образом, в горохе под влиянием NH 4 C1 мембранные ферменты, в основном, активировались сильнее, чем цитозольные, указывая на мембранотропное действие этой соли.
На субклеточных мембранных структурах, выделенных из корней гороха, это положение подтвердилось. В табл.4 показано, что во фракции объединяющей ядра и пластиды, активность СОД увеличивалась в 15, КАТ - в
10, ПО - в 2 раза; в митохондриях, соответственно - в 4; 2 и 2,5 раза; в субчастицах - в 2,6; 6 и 2 раза, по сравнению с контролем. Интересно отметить 10-15 - кратное увеличение активностей СОД и КАТ в общей фракции ядер и пластид. Не исключено, что оно обусловлено повышенной "атакой" этих органоидов, но возможен также индуцированный синтез защитных ферментов de novo, подобно тому, как это происходит с СОД и ПО в зараженных растительных тканях [10, 11,23].
Мембранотропное действие хлористого аммония подтверждалось также на мембранных структурах хлоропластов, выделенных из листьев гороха, табл.5. Функционально активные хлоропласты 2 класса [17], выделяли из гороха выращенного при 2,5x10-3 M NH 4 C1 (при концентрации соли на порядок больше, активные хлоропласты выделить не удалось). Как видно из таблицы хлоропласты растений, выращенных при подкормке хлористым аммонием активно синтезировали АТФ в присутствии экзогенных переносчиков электронов - МВ и ФМС, по сравнению с контрольными. В таких условиях, как известно, протекает псевдоциклический поток электронов, сопряженный с функционированием
Таблица 5. Влияние хлористого аммония на активность антиоксидантных ферментов и фотофосфорилирование в изолированных хлоропластах гороха
|
Условия опыта |
Активность, |
Синтез АТФ (мкмоль/ч мг хлорофилла) в присутствии |
|||
|
СОД |
КАТ |
ПО |
|||
|
МВ |
ФМС |
||||
|
Контроль 2,5x10-3M NH 4 C1 |
7 18 |
0,8 1,2 |
10 15 |
271 (93) 356 (95) |
518(123) 584 (123) |
Примечание. Водная культура, возраст 12 сут. В скобках указан синтез АТФ при добавлении в среду ДХММ. По фотофосфорилированию представлены средние данные 2 - х опытов, отклонение от среднего ± 3-5 %.
двух фотосистем, образованием АТФ и восстановлением кислорода до Н2О2 [18]. Доказательством такого потока электронов в модельных опытах с хлоропластами служит ингибирование его диуроном (ДХММ) [24], в данном случае происходило ингибирование синтеза АТФ, показанное в скобках. Активности всех трех ферментов в хлоропластах были повышены, по сравнению с контролем: СОД - в 2,6 , КАТ и ПО - в 1,5 раза. Заслуживает внимания тот факт, что СОД активировалась сильнее, чем два другие фермента, что свидетельствовало в пользу интенсивного образования супероксидного радикала кислорода. Можно думать, что индуцируемая аммонием, активация антиоксидантных ферментов в хлоропластах обеспечивала защиту тилакоидных мембран от окислительного разрушения. Возможная регуляция, по-видимо-му, способствовала стимулированию псевдо-циклического фотофосфорилирования, самого по себе, продуцирующего Н2О2 [18]. Оказалось, что эта защита уязвима при увеличении действующей концентрации соли на хлоропласты. Так, исследование псевдо-циклического транспорта электрона и фотофосфорилирования показало, что добавление 10-6-10-2М NH4C1 непосредственно в реакционную среду инкубации хлоропластов усиливает поглощение кислорода с МВ и ингибирует ФМС-зависимый синтез АТФ (рисунок А, Б). Очевидно, что при концентрациях NH4C1 больших, чем 10-3М, в хлоропластах наступало разобщение электронного транспорта от фотофосфорилирования, сопровождаемое усилением окислительной активности. Последнее, соответствует литературным данным, показывающим, что разобщение высокими концентрациями аммония приводит к нарушению структуры тилакоидных мембран [25, 26].
Обсуждение результатов
Выбор хлористого аммония в качестве токсического агента в нашей работе был не случаен. Во-первых, аммиак относится к числу приоритетных загрязнителей сельского хозяйства и заводов производства минераль-
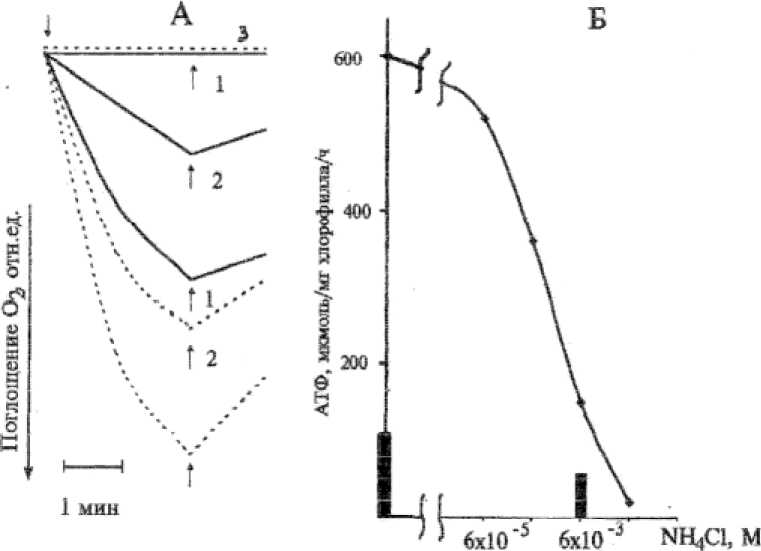
Рис. Влияние экзогенного NH4Cl на электронный транспорт и фотофосфорилирование в хлоропластах гороха.
А - Поглощение кислорода в присутствии МВ. 1 и 2 - горох выращен на почве в отсутствие и при подкормке NH4Cl, 10 -3 М, соответственно; прерывистые линии - то же, добавлен NH4Cl, 6х103 М в инкубационную среду хлоропластов; 3 - то же, что 1 и 2, добавлен ДХММ. Стрелками указано включение и выключение света.
Б - ФМС-зависимое фотофосфорилирование, столбиками отмечен уровень синтеза АТФ при добавлении ДХММ. Возраст гороха 27 сут.
ных удобрений в нашем регионе [27]. Во-вторых, механизм ингибирующего эффекта ионов аммония [10-2 М] на фотосинтез хорошо изучен [25, 26]. Как протонофор [28] он вызывает разобщение в энергетических мембранах. Исходя из того, что разобщение в мембранах сопряжено с инициированием ПОЛ, в регулировании которого принимают участие СОД, КАТ и ПО [14,29], нами было предположено изменение активностей антиоксидантных ферментов в растениях под влиянием ксенобиотка. Сопоставляя морфологические и биохимические изменения можно было идентифицировать мишени воздействия ксенобиотика по активностям ферментов, судить о регуляции и деструкции процесса по токсическому действию, а также выявить возможность биотестирования повреждений на биохимическом уровне.
Физиологические опыты показали, что постоянное и длительное введение в среду выращивания гороха даже невысоких концентраций хлористого аммония в пределах 10-610-4 M NH 4 C1 в течение вегетационного периода вызывали видимые нарушения в морфологии листьев и корней, несмотря на то, что аммонийный азот хорошо усваивается растениями. По-видимому, с течением времени в среде выращивания создается избыток его также как и при высоких концентрациях - 10-3-10-2 M, о чем свидетельствуют биохимические и морфологические нарушения, а также гибель растений.
Как и предполагалось, действие данной соли на горох изменяло активность антиоксидантных ферментов - СОД, КАТ и ПО. Это происходило в листьях и корнях, в мембранах и цитозоле клеток, было связано с концентрацией токсиканта и развивалось со временем. При кратковременном воздействии на горох повышенных концентраций хлористого аммония, все составляющие системы СОД-КАТ-ПО активировались значительно сильнее, чем при меньших концентрациях. Следует отметить соответствие усиления активности СОД в листьях и ПО во всем растении при появлении повреждений, связанных с длительным введением меньших концентраций соли в среду. Примечательно, что до наступления деструктивных процессов, приво дящих растения к гибели, антиоксидантная система: СОД-КАТ-ПО сильнее активировалась в мембранных структурах, об этом свидетельствует больший рост активностей в мембранных белковых фракциях, по сравнению с цитозольными и в субклеточных мембранных структурах ядрах, пластидах, митохондриях и хлоропластах. Эти данные подтверждали мембранотропное действие хлористого аммония и указывали на повышенное образование супероксидрадикала, поскольку СОД во всех мембранных структурах активировалась сильнее, чем КАТ и ПО. На примере функционально активных хлоропластов, можно было видеть, что регуляция ферментами метаболизма АФК, образуемых при действии мембранотропного агента, в значительной степени определялась концентрацией токсиканта. Следовательно, активирование ферментов СОД, КАТ и ПО в горохе под действием хлористого аммония в малых и высоких концентрациях на ранних этапах развития растений можно рассматривать как защиту мембран от окислительного повреждения, вызванного этой солью. На наш взгляд, лабильность антиоксидантной системы, выраженная в широкой вариабельности активностей каждого фермента в целом растении, указывают на регуляцию, поддерживающую нормальный метаболизм в растениях. Такое положение не противоречит литературным данным о регулирующей роли СОД, КАТ и ПО в ПОЛ [8, 14, 29], участии СОД в предотвращении мутаций [30], а ПО -в иммунитете растений [9-11].
Однако, существует и другой аспект, а именно высокое активирование ПО в горохе, ее цитозольной формы, особенно в корнях, где хлористый аммоний непосредственно ассимилируется [31]. ПО способна к окислительной детоксикации чужеродных соединений в цитоплазме клеток [10, 15], как цитохромом Р- 450 в микросомах [20]. Особенность ПО заключается в том, что проявляя оксидазную активность, она вовлекает молекулярный кислород в пероксидазный механизм и продуцирует супероксид (О2-), гидроксильный радикал (ОН-) и гидроперекиси [32], иными словами, обладает не только анти-, но и прооксидативными свойствами.
Есть мнение о том, что продукты пероксидазного окисления, такие как (ОН-) ингибируют СОД, КАТ и разрушают мембраны, тонопласт и плазмолемму растительных клеток [9-11, 23]. Исходя из сказанного, можно думать, что сильная активация пероксидазы под действием хлористого аммония связана не только с защитой мембран, но и с окислением NH 4 C1 как токсиканта. При этом следует иметь ввиду возможность окисления продуктов ассимилированного иона аммония - аминокислот, амидов и азотистых оснований, образуемых по пути аммония из корней в листья [31]. Все эти продукты могут служить субстратами в пероксидазно-оксидазных реакциях [5]. При оксидазном механизме пероксидаза может использовать ион C1- [32, 33], что тоже следует учитывать при воздействии хлористого аммония на растения. Предпосылкой таких событий являются химические процессы, развивающиеся при проникновении ионов аммония и хлора через мембраны - повышение рН внутри клеток и органелл [34], образование хлорированных гидроперекисей. Последние особенно способствуют инициированию свободнорадикальных реакций ПОЛ и пероксидазно-ок-сидазного механизма [5, 32, 35].
Таким образом, сильное увеличение пероксидазной активности в цитозоле, особенно в корнях, где ксенобиотик непосредственно контактирует с мембранами клеток, и сопутствующая активация СОД под влиянием NH4C1, указывают на возможное окисление хлористого аммония и его метаболизирующих продуктов в горохе. Это подтверждается и существенным активированием антиоксидантных ферментов в субчастицах, в которых присутствуют пероксисомы, микросомы и др. мембранные структуры, содержащие окислительные системы [15,20]. Мы предполагаем, что токсичный избыток хлористого аммония, подобно другим факторам [3, 35, 36], инициирует окислительный стресс, сопровождающийся стимуляцией ПОЛ, бесконтрольным образованием новых радикалов и цепей радикальных реакций, деструктивными процессами в мембранах, приводящими к их деградации и, в конечном итоге, к гибели растений. Одновременное активирование фермен тов СОД и ПО, при более сильном увеличении пероксидазной активности (в 10 и более раз), вероятно, может служить индикатором действия загрязняющих токсических веществ на растения, когда повреждения еще не видны, а биохимическая структура мембран нарушается и находится на критическом уровне.
