Инсулиноподобные факторы роста в патогенезе и прогнозе рака яичников

Автор: Бочкарева Наталья Валерьевна, Кондакова Ирина Викторовна, Коломиец Лариса Александровна, Мунтян Алиса Борисовна
Журнал: Сибирский онкологический журнал @siboncoj
Рубрика: Обзоры
Статья в выпуске: 3 (45), 2011 года.
Бесплатный доступ
Обсуждается роль системы инсулиноподобных факторов роста в нормальном фолликулогенезе и отдельных аспектах патогенеза эпителиального рака яичников. Дальнейшая комплесная оценка уровня экспрессии показателей системы инсулиноподобных факторов роста и параметров, ассоциированных с клеточной подвижностью, при эпителиальном раке яичников представляет определенный интерес как в плане углубленного изучения патогенеза этой гетерогенной группы опухолей, так и в плане поиска новых молекулярных прогностических и предикторных факторов.
Система инсулиноподобных факторов роста, эпителиальный рак яичников, клеточная подвижность, прогноз
Короткий адрес: https://sciup.org/14055917
IDR: 14055917 | УДК: 618.11-006.6-092-037
The system of insulin-like growth factors in pathogenesis and prognosis of epithelial ovarian cancer
The role of the system of insulin-like growth factors (IGF) in normal folliculogenesis and in different aspects of pathogenesis of epithelial ovarian cancer is discussed. Further comprehensive assessment of IGF-system expression levels and cell motility-associated parameters in epithelial ovarian cancer are of great interest both for the advanced study of pathogenesis of these heterogeneous tumors and for the development of new molecular prognostic and predictive factors.
Текст обзорной статьи Инсулиноподобные факторы роста в патогенезе и прогнозе рака яичников
От рака яичников (РЯ) ежегодно умирают больше женщин, чем от рака шейки матки и рака эндометрия вместе взятых. Несмотря на это, многие патогенетические аспекты данной патологии остаются малоизученными, в частности роль ростовых факторов [1, 4, 5]. В большинстве экспериментальных исследований показано, что инсулиноподобные факторы роста являются мощными митогенами, мотогенами (индукторами локомоторной активности клеток), снижают чувствительность клеток к апоптотическим стимулам и, таким образом, повышают выживаемость клеток. Опухоли яичников эпителиального происхождения составляют около 90 % всех злокачественных новообразований яичника, поэтому в данном обзоре система инсулиноподобных факторов роста при эпителиальном РЯ будет рассмотрена в плане изучения некоторых патогенетических аспектов и поиска возможных предикторных и прогностических показателей.
Характеристика инсулиноподобных факторов роста, их клеточных эффектов и путей регуляции биодоступности
В настоящее время сформировано понятие о сигнальном пути, в который входят инсулиноподобные факторы роста (ИФР) – ИФР-I, ИФР-II и рецептор ИФР I типа. Регуляция этой системы на клеточном и тканевом уровне осуществляется 6 белками, связывающими инсулиноподобные факторы роста (IGFBPs) и расщепляющими их протеиназами [13]. Основным источником ИФР в организме человека является печень, но эти пептиды также синтезируются в большинстве тканей организма [2]. Показана возможность продукции ИФР злокачественными опухолями [24].
Рецептор ИФР I типа (IGF-1R) – член семейства рецепторных тирозинкиназ, состоит из двух α-субъединиц и двух β-субъединиц. При активации IGF-1R путем связывания с ИФР с экстрацеллюлярной α-субъединицей происходит активация киназного домена, что, в свою очередь, ведет к рекрутированию в область β-субъединиц адапторных молекул, таких как субстраты инсулинового рецептора 1 и 2. В результате активируются множественные сигнальные пути, включающие путь с вовлечением митоген-активируемой протеинкиназы и фосфатидилинозитол-3-киназный путь, которые ведут к стимуляции клеточной пролиферации, подавлению апоптоза в клетках, клеточной подвижности и другим эффектам (рис. 1) [35].
В дополнение к связыванию с IGF-1R на клеточной мембране, ИФР с высокой аффи-ностью связываются с 6 специфическими протеинами – белками, связывающими инсулиноподобные факторы роста (IGFBPs). IGFBPs обнаружены в широком спектре биологических жидкостей: в сыворотке крови, в амниотической, цереброспинальной, семенной, фолликулярной жидкостях, а также в культуральных клеточных линиях [2, 13]. Важнейшей функцией всех IGFBPs является ограничение эффектов ИФР путем связывания с ними вблизи мембраны в экстраклеточном матриксе. Некоторые посттрансляционные модификации IGFBPs, такие как фосфорилирование, протеолиз и полимеризация, существенно изменяют их функции и, в первую очередь, влияют на способность связывать ИФР [25].
В физиологических концентрациях ИФР-I и ИФР-II защищают многие типы клеток в экспериментах in vitro от различных проапоптотиче-ских стимулов, включая экспозицию этопозида, сверхэкспрессию c-myc, полное удаление факторов роста из инкубационной среды, ультрафиолетовое облучение, активацию Fas-рецептора. В экспериментах in vivo сниженная экспрессия как ИФР-I, так и ИФР-II ассоциировалась с активацией апоптоза. Например, использование антисмысловой олигонуклеотидной последовательности к мРНК ИФР-II значительно редуцирует пролиферацию клеток панкреатической карциномы и индуцирует апоптоз [10]. В ряде исследований было показано, что при
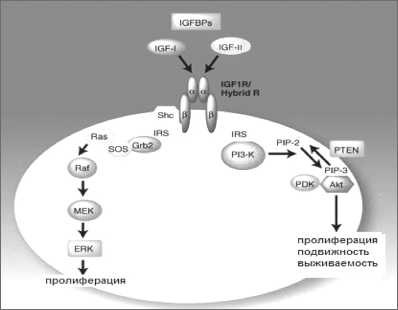
Рис. 1. ИФР-опосредованный сигнальный путь. Примечание: IGF-1R – ИФР-рецептор I типа, IGF-I и IGF-II – инсулиноподобные факторы роста – лиганды IGF-1R, IGFPs – протеины, связывающие ИФР и регулирующие их биодоступность, IRS – внутриклеточные cубстраты инсулинового рецептора, Ras, Raf, MEK и ERK – компоненты митоген-активируемого протеинкиназного сигнального пути, PI3-K – фосфатидилинозитол-3 киназа, PIP-2 – фосфатидилинозитолдифосфат, PIP-3 – фосфатидилинозитолтрифосфат, PDK – фосфонозитидзави-симая киназа, Akt – компонент PI3-K/Akt сигнального пути, PTEN – внутриклеточная фосфатаза, продукт гена-супрессора, регулирует активность PI3-K/Akt сигнального пути связывании ИФР со своим рецептором в клетках ингибируются эффекторные каспазы-3 и -7 на этапе их процессирования [28]. Кроме того, выявлена активация экспрессии мРНК IGFBP-3 при экспозиции ДНК-повреждающих факторов в клетках рака молочной железы, экспрессирующих дикий, но не мутантный р53 [10]. В исследованиях F. Garouste et al. [14] было выявлено, что ИФР-I и ИФР-II защищают клетки колоректальной карциномы человека HT29-D4 от апоптоза, индуцированного фактором некроза опухолей альфа.
Следует отметить, что большинство данных о роли ИФР-опосредованного сигнального пути в процессах пролиферации, апоптоза и выживаемости получены на клеточных культурах и некоторых клеточных опухолевых линиях. В то же время отсутствуют аналогичные данные в отношении нетрансформированных и трансформированных тканей, что представляет значительный интерес.
Внутриклеточная деградация компонентов IGF-1R/IGF комплекса
Регуляция ИФР-зависимого сигнального пути осуществляется также посредством эн- доцитоза и транспортировки внутрь клетки самого рецептора или рецептор-лигандного комплекса. Детальные механизмы интернализации IGF-1R не выяснены, но выявлено, что митогенный эффект от связывания IGF-1R со своими лигандами регулируется специфической молекулой – эндоцитоз-зависимым адаптерным протеином-2. Документировано лиганд-индуцированное рекрутирование рецептора внутрь кальвеолы при эндоцитозе [26]. Протеолитическая деградация интернализированных ИФР-I и ИФР-II охарактеризована не полностью, но в этом процессе важную роль отводят катепсину L и E-64-зависимой цистеиновой протеиназе [23]. Основную роль в деградации самого рецептора играет внутриклеточная убик-витинзависимая протеасомная система деградации. Показано, что связывание протеина MDM2 с β-субъединицей IGF-1R через адаптерный белок β-аррестин приводит к рекрутированию убиквитина к рецептору, таким образом, инициируется его деградация [16].
Роль инсулиноподобных факторов роста и связывающих их белков в фолликулогенезе
В последнее время установлена роль системы ИФР в фолликулогенезе у женщин. Показано, что доминантный фолликул содержит ИФР-I в большей концентрации, чем недоминантные. Уровень ИФР-I в фолликулярной жидкости положительно коррелировал как с концентрацией эстрадиола и прогестерона, так и с объемом фолликулярной жидкости. При использовании метода гибридизации in situ транскрипты ИФР-I были обнаружены в теке малых (3–5 мм) развивающихся фолликулов, но не найдены в гранулезных клетках. Однако ИФР-II мРНК наиболее часто встречались в гранулезных клетках больших развивающихся фолликулов и в желтом теле. Низкий уровень ИФР-II мРНК и IGFBP-6 был обнаружен в текаклетках женщин с синдромом поликистозных яичников, так же как и в малых антральных фолликулах у женщин с нормальной функцией яичников [2].
ИФР-зависимая IGFBP-2, -4 и -5-протеаз-ная активность была выявлена в различных типах клеток (фибробласты, остеобласты, децидуальные клетки, клетки гранулезы яичника, эндометриальные стромальные клетки). Данная протеаза была идентифицирована как ассоциированный с беременностью протеин (pregnancy-associated plasma protein-A (PAPP-A) [18, 20]. Показано, что у млекопитающих рост доминантного фолликула в терминальной фазе сопровождается значительным снижением ин-трафолликулярного уровня IGFBP-2 и IGFBP-4. В исследовании N. Gerard et al. [15] высокий уровень PAPP-A выявлен в фолликулярной жидкости ранних и поздних доминантных фолликулов у лошадей, однако практически не определялся в антральных фолликулах, что свидетельствует о возможной роли PAPP-A в фолликулогенезе и овуляции.
Значение инсулиноподобных факторов роста, их рецептора и белков, связывающих инсулиноподобные факторы роста в патогенезе рака яичников. Связь параметров ИФР-системы с эффективностью химио- и гормонотерапии при раке яичников
В литературе имеются немногочисленные данные об уровне отдельных показателей ИФР-системы при РЯ эпителиального происхождения. Так, с использованием метода ПЦР в реальном времени был изучен уровень экспрессии IGF-I и IGF-1R в злокачественных и доброкачественных опухолях яичника, который оказался выше в злокачественных опухолях с максимальными значениями у больных с плохим прогнозом, после субоптимальных резекций и с большим объемом остаточной опухоли [7]. Уровень IGFBP-2 и IGFBP-5 был повышен в низкодифференцированных серозных карциномах яичников по сравнению с нормальными тканями яичников, серозными кистами, пограничными серозными опухолями и высокодифференцированными аденокарциномами яичников [33].
Ряд интересных данных получен на экспериментальных клеточных линиях. Так, гиперэкспрессия IGF-1R и фосфатидилинозитол-3-киназы (компонента ИФР-зависимого сигнального пути) ассоциирована с платиноре-зистентностью некоторых клеточных линий РЯ [12]. На примере экспериментальной клеточной линии эндометриоидного РЯ было показано, что IGFBP-3 действует как опухолевый супрессор, эффективно подавляющий клеточную миграцию, инвазию, метастазирование, но не влияющий на клеточную пролиферацию. Клинически, у 35 больных с эндометриоидным РЯ с низким уровнем IGFBP-3 рак ассоциировался с низкой степенью дифференцировки, запущенностью процесса и плохим прогнозом [31]. При исследовании влияния таксола на степень фосфорилирования Akt – компонента инозитолтрифосфат/ Akt-сигнального пути, активирующегося при связывании ИФР с IGF-1R, – было показано, что таксол-индуцированное фосфорилирование Akt происходит при участии тирозинкиназы IGF-1R (активирование рецептора). Клеточная линия РЯ, резистентная к таксанам, имела высокий уровень IGF-II, а снижение уровня IGF-II восстанавливало чувствительность клеток к таксанам. Также было показано, что высокий уровень IGF-II в опухоли коррелировал с запущенной стадией, низкой степенью дифференцировки и редуцированной безрецидивной выживаемостью [19].
В литературе имеются разноречивые данные о связи параметров ИФР-системы с эффективностью химио- и гормонотерапии РЯ. В исследованиях G. Walker et al. [32] на эстроген положительных клеточных линиях РЯ была показана позитивная регуляция 17β-эстрадиолом экспрессии гена IGFBP-4 и негативная – генов IGFBP-3 и IGFBP-5. Экспрессия генов IGFBP-1, -2 и -6 была эстроген независима. На основании полученных данных было высказано предположение о возможности использования данных по экспрессии генов или белков некоторых IGFBPs в качестве предикторных параметров в отношении эффективности гормонотерапии ингибиторами ароматазы у больных РЯ с рецидивами. Используя сывороточный уровень СА-125 как индикатор ответа на проводимую терапию, была выявлена статистическая разница между экспрессией некоторых IGFBPs в опухоли (иммуногистохимическое окрашивание) в группах больных со стабильным течением заболевания по сравнению с группой больных с прогрессированием на фоне гормонотерапии. Средний уровень экспрессии IGFBP-3 и-5 был значительно ниже, а IGFBP-4 – значительно выше в первичной опухоли у больных, демонстрирующих ответ на проводимую гормонотерапию, по сравнению с группой с прогрессированием. Необходимо также отметить высокую частоту экспрессии IGFBP-3, -4 и -5 в первичных злокачественных опухолях яичника, составившую
95, 30 и 44 % соответственно. При проведении математического анализа методом многофакторной логистической регрессии было показано, что наибольшей предсказательной силой в отношении отвечаемости на гормонтерапию было сочетание IGFBP-4, IGFBP-5 и ERα, однако использование одного IGFBP-3 было лишь незначительно хуже вышеупомянутой комбинации [32]. В то же время, в исследованиях L. Lu et al. (2006) не было выявлено никакой ассоциации между экспрессией гена IGF-II, IGFBP-3 в опухоли и ответом на платиносодержащую химиотерапию 1-й линии у больных РЯ [22].
Роль параметров ИФР-системы в инвазии и метастазировании опухолей: некоторые молекулярные механизмы
Метастазирование опухолей – многоступенчатый процесс, складывающийся из сложных взаимоотношений между диссеминирующими опухолевыми клетками и изменяющимся микроокружением. Среди основных этапов, лимитирующих этот процесс, выделяют опухоль-индуцированную неоваскуляризацию (ангиогенез и лимфоангиогенез), деградацию экстраклеточного матрикса, миграцию клеток через тканевые барьеры, включая вход и выход метастазирующих опухолевых клеток из кровеносных сосудов, пролиферацию и выживаемость клеток в новом, не свойственном им микроокружении. Ряд авторов считают, что опухолевая инвазия является по своей сути нерегулируемой клеточной подвижностью [21]. Суммируя данные, полученные в основном на клеточных линиях, можно сказать, что компоненты ИФР-системы играют регуляторную роль практически на всех этапах метастазирования (рис. 2) [26].
Относительно недавно получены данные о механизмах участия ИФР и IGF-1R в процессах клеточной адгезии и миграции. Хотя ингибирование IGF-1R сигнального пути существенно ограничивает рост клеток рака молочной железы in vitro и in vivo, однако очень низкий уровень IGF-1R в этих клетках ассоциировался с высоким риском метастазирования и неблагоприятным клиническим прогнозом. Показано, что сверхэкспрессия IGF-1R на клетках рака молочной железы сокращает их потребность в эстрогенах для пролиферации,

Роль ИФР-системы в выживаемости опухолевых клеток в новом микроокружении
- Экспериментальные данные о значительном уменьшении размеров метастазов в печень экспериментальной опухоли, экспрессирующей мутантную форму рецептора IGF-1R, по сравнению с размерами метастазов опухоли с нормальным вариантом IGF-1R у мышей.
- Экспериментальные данные о большем размере первичной опухоли и ее метастазах в печень у мышей с генетически детерминированным сниженным уровнем синтеза IGF-I в печени и сыворотке крови при дополнительных инъекциях IGF-I по сравнению с нелеченными.
Роль ИФР-системы в ангиогенезе и лимфоангиогенезе:
- ИФР-1 и II через HIF-la индуцируют синтез VEGF - ИФР-1 и II индуцируют миграцию эндотелиальных клеток - IGF-1R позитивный регулятор продукции VEGF-C - основного ангиогенного фактора, регулирующего лимфоангиогенез
Роль ИФР-системы в клеточной адгезии и подвижности:
- изменение клеточной адгезии и подвижности через регулирование стабильности кадгерин-катенинового комплекса IGF-1R
- ИФР-независимая стимуляция клеточной подвижности IGFBP-2 и -5
Деградация экстрацеллюлярного матрикса:
- Регуляция продукции и активации ММП-2 и ММ П-9 через экспрессию IGF-1R
- Регуляция экспрессии иРА и uPAR IGF-1R
Рис. 2. Предполагаемая роль ИФР-системы в инвазии и метастазировании опухолей.
Примечание: ММП-2 и -9 – матриксные металлопротеиназы-2 и -9; uPA и uPAR – урокиназный активатор плазминогена и его рецептор
повышает их выживаемость и тем не менее ограничивает их подвижность и рассеивание, способствуя E-кадгерин-опосредованной клеточной адгезии [17].
В исследовании A.P. Pennisi et al. [24] на клеточной линии MCF-7 (клетки высокодифференцированного рака молочной железы с низкометастатическим и низкоинвазивным фенотипом) была показана важная роль IGF-1R в клеточной адгезии и подвижности. Клетки MCF-7 трансфецировали антисмысловой олигонуклеотидной последовательностью к IGF-1R, что приводило к уменьшению экспрессии рецептора примерно на 50 %. Используя тесты для функциональной оценки подвижности, прилипания и агрегации, было выявлено 3-кратное повышение миграционной активности клеток, практически полная потеря способности к прилипанию и снижение клеточной агрегации. Функциональные изменения сопровождались снижением экспрессии E-кадгерина, повышением уровня р120 протеина в цитозоле опухолевых клеток и значительным снижением уровня р120 в Е-кадгерин-катенин-р120 комплексе. Эти изменения сопровождались существенными изменениями в соотношении белков семейства малых ГТФ-аз, ответственных за миграцию клеток [24]. Таким образом, доказано, что не связанный с лигандом IGF-1R необходим для формирования функционально активного и стабильного кадгерин-катенинового комплекса. Формирование данного комплекса приводит к полноценному межклеточному контакту, повышенной адгезии клеток и снижению миграционной активности. Снижение экспрессии
IGF-1R приводит к снижению количества кадгерин-катениновых комплексов и изменению их конформации. Далее происходит перераспределение β-катенина и р120 протеина в цитозоль опухолевых клеток, что ведет к дифференциальной активации белков – членов семейства Rho, повышению подвижности клеток и, как следствие, к метастазированию. Предполагают, что активация IGF-1R посредством связывания со своими лигандами (ИФР-I, ИФР-II) и дальнейшее повышение клеточной подвижности объясняются выходом IGF-1R из кадгеринового комплекса [24].
Сами ИФР являются для большинства клеток мощными факторами миграции и стимулируют клеточную подвижность [13]. В ряде исследований выявлена двойственная роль IGFBPs в отношении клеточной подвижности. Так, было показано, что IGFBP-1 в высокометастатической клеточной линии рака молочной железы MDA-231BO снижал клеточную подвижность, индуцированную ИФР-I [35]. Однако также было выявлено, что сами IGFBP-1 и IGFBP-2 благодаря наличию в своей структуре интегрин-связывающего мотива (интегрины участвуют в передаче внеклеточных сигналов до элементов цитоскелета) в отсутствие ИФР стимулируют клеточную подвижность [13]. IGFBP-5 также стимулирует ИФР-независимую миграцию мезангиальных клеток и формирование в них филоподий (морфогенетические изменения, присущие движущимся клеткам) [8]. Таким образом, все компоненты ИФР-зависимого сигнального пути, так или иначе, вовлечены в процессы клеточной адгезии и миграции, что может свидетельствовать об их участии в инвазивном росте и метастазировании.
Связь параметров ИФР-системы с прогрессированием рака яичников.
Молекулярные механизмы прогрессии РЯ в значительной степени не изучены. Это связано с тем, что для РЯ, как ни для какой другой опухоли, свойственны все возможные способы распространения – per continuitatem (по протяжению), лимфогенно и гематогенно [3]. На клиническом уровне прогрессирование РЯ обычно сопряжено с появлением новых «проявлений болезни» после достижения частичной или полной регрессии на фоне первой линии химиотерапии (рецидивирующий, резистентный и персистирующий РЯ), или отсутствием эффекта или прогрессированием на фоне первой линии химиотерапии (рефрактерный рак яичников) [4–6].
Несколько исследований посвящены изучению уровня показателей ИФР-системы во взаимосвязи с клинико-морфологическими параметрами, являющимися доказанными прогностическими факторами, и прогнозом в целом, при РЯ. Так, было показано, что женщины с эпителиальным РЯ и высоким уровнем мРНК ИФР-I и самого белка имеют худший прогноз по сравнению с женщинами с низкими показателями обоих маркеров. Высокий уровень свободного, но не общего ИФР-I в опухоли ассоциирован с высоким риском прогрессирования [9]. Высокий уровень ИФР-II, но не IGFBP-3, в злокачественных эпителиальных опухолях коррелировал с плохим прогнозом, а именно – с запущенной стадией, гистологическим типом опухоли – серозный рак, низкой степенью дифференцировки, размером остаточной опухоли более 2 см [22]. С использованием метода ПЦР в реальном времени была изучена экспрессия гена ИФР-II в 109 образцах РЯ. Показано, что экспрессия гена ИФР-II в опухолях более чем в 300 раз превышала экспрессию гена в нормальном эпителии яичника. Высокая экспрессия ИФР-II ассоциировалась с III–IV стадией заболевания, низкой степенью дифференцировки и субоптимальным объемом резекции. При построении логистической регрессионной модели экспрессия ИФР-II в опухоли явилась независимым прогностическим фактором только для серозных эпителиальных опухолей, но не для опухолей несерозного гистотипа [27]. В исследовании D. Spentzos et al. [30] изучались взаимосвязи экспрессии генов ИФР-системы и ИФР-зависимого сигнального пути с выживаемостью у больных РЯ. При проведении одно- и многофакторного анализа прогностической значимости изученных параметров выяснилось, что экспрессия гена IGFBP-4 и гена субстрата инсулинового рецептора, соответственно, негативно и позитивно ассоциировались с выживаемостью.
В связи с доступностью асцитической жид- кости для исследования, высоким уровнем экспрессии отдельных маркеров как в клеточной, так и во внеклеточной части асцитической жидкости ряд современных исследований сосредоточены на определении диагностической и прогностической роли белков экссудата [11, 34]. Что касается белков ИФР-системы, то опубликованы результаты исследования, в котором был изучен уровень ИФР-II и IGFBP-3 в опухоли и асцитической жидкости у больных РЯ. Высокий уровень IGFBP-3 до химиотерапии и высокий уровень IGF-II после химиотерапии в асцитической жидкости коррелировал с низкой общей выживаемостью. Последний фактор явился независимым прогностическим фактором по данным многофакторного регрессионного анализа [29].
Заключение. Таким образом, проведенный анализ данных литературы свидетельствует об определенном значении инсулиноподобных факторов роста, их рецептора и связывающих их белков как в фолликулогенезе, так и в отдельных звеньях патогенеза злокачественных опухолей яичников эпителиального происхождения. Дальнейшая комплексная оценка уровня экспрессии показателей ИФР-системы и параметров, ассоциированных с клеточной подвижностью, при раке яичников представляет определенный интерес как в плане углубленного изучения патогенеза этой гетерогенной группы опухолей, так и в плане поиска новых молекулярных прогностических и предиктор-ных факторов.
