Интенсивная физическая нагрузка и энтеросорбция уменьшают массу жировой ткани у экспериментальных животных

Автор: Любаева Екатерина Владимировна, Малыхина Татьяна Викторовна, Марков Игорь Иванович, Альхимович Владимир Леонидович
Журнал: Морфологические ведомости @morpholetter
Рубрика: Оригинальные исследования
Статья в выпуске: 1 т.24, 2016 года.
Бесплатный доступ
Работа выполнена на беспородных белых крысах (возраст - 4-е месяца) массой 160,0+8,0г (n=15). Животные 1 группы (n=10) выполняли интенсивную мышечную работу динамического характера (плавание), животные II группы (n=5) являлись контрольными. Животным 1 группы per os вводился энтеросорбент «Мегамин» ежедневно по 0,5 г в течение 30 суток. Материал (камбаловидная и широкая латеральная мышка бедра, подкожно - жировая клетчатка, стенка тонкой кишки, большой сальник) изучался на серийных парафиновых срезах, окрашенных гематоксилином и эозином, по Гейденгайма, по Шабадашу. Полученные данные свидетельствуют об увеличении массы мышечной и уменьшению массы жировой ткани, повышении функциональной активности микрососудистого русла стенки тонкой кишки и большого сальника.
Физическая нагрузка, жировая ткань, энтеросорбция
Короткий адрес: https://sciup.org/143177094
IDR: 143177094
The intensive physical exercise and enterosorption decrease of the adipose tissue mass in experimental animals
The study was performed on outbred albino rats (aged 4 months) weighing 160,0+8,0г (n=15). The animals of group 1 (n=10) had strenuous dynamic exercise (swimming) while the animals of group P (n=5) were controls. The animals of group 1 were administered 0.5 g the Megamin enterosorbent daily for 30 days/ The material (soleus musdcle and vastus laterakis muscle of the thigh, hypodermically - fatty tissue, intestine walls, greater omemtum) was examined on serial paraffin sections staned by hematoxykin and eosin according to Heidenhain and to Shabadash. The findings attest the increase in muscle bulk and decrease in adipose tissue bulk, as well as the enhanced microvasculature activity of the intestine wall and greater omentum.
Текст научной статьи Интенсивная физическая нагрузка и энтеросорбция уменьшают массу жировой ткани у экспериментальных животных
The study was performed on outbred albino rats (aged 4 months) weighing 160,0+8,0г (n=15). The animals of group 1 (n=10) had strenuous dynamic exercise (swimming) while the animals of group P (n=5) were controls. The animals of group 1 were administered 0.5 g the Megamin enterosorbent daily for 30 days/ The material (soleus musdcle and vastus laterakis muscle of the thigh, hypodermically – fatty tissue, intestine walls, greater omemtum) was examined on serial paraffin sections staned by hematoxykin and eosin according to Heidenhain and to Shabadash.
The findings attest the increase in muscle bulk and decrease in adipose tissue bulk, as well as the enhanced microvasculature activity of the intestine wall and greater omentum.
Введение. Важное значение при диабете имеют изменения липидного обмена и связанные с ними нарушения функции сердечно-сосудистой и нервной систем [1, 3, 5]. Среди всех тканей живых организмов жировая ткань самая «молодая» [6]. Считается, что даже такие позвоночные, как рыбы, не имеют адипоцитов и депонируют триглицериды между миоцитами [7]. У млекопитающих же сформировались даже новые «молодые» органы: жировые тела щек [8] и большой сальник [9]. Общее количество жировой ткани у взрослого мужчины около 20 кг, а у тучных людей – на десятки килограммов больше [10]. Адипоциты – представители особой клеточной линии, 50% из них – это стромальные клетки. Жировая ткань – важный эндокринный орган [11], который секретирует в кровь гормоны (адипокины), регулирующие энергетический обмен и другие физиологические процессы. Среди них: адипонектин, лептин и резистин. Именно они играют решающую роль в развитии и поддержании инсулинрезистент-ности (ИР) - снижении биологического ответа тканей на один или несколько эффектов инсулина [1]. Гипергликемия сама по себе не может быть причиной как ИР, так и недостаточности β-клеток островков Лангерганса. Основным компонентом патогенеза СД-2-го типа является инсулинре-зистентность, наличие которой способствует повышенному образованию глюкозы печенью и снижение инсулинопосредовательной её утилизации периферическими тканями [12, 13, 14]. Формирование гипергликемии в филогенезе было вызвано изменением характера питания живых организмов - чередованием периодов избытка пищи и голоданием. Это характерно для многоклеточных организмов, ведущих малоподвижный образ жизни и приём пищи малыми порциями [7]. За миллионы лет до того, как был синтезирован инсулин, они же пассивно поглощали глюкозу и депонировали её в форме гликогена [15]. В цитоплазме гладких миоцитов животных, стоящих на разных ступенях филогенетической лестнице, гистохимическими методами гликоген был обнаружен многими исследователями [16, 17, 18, 19].
Изменение условий питания животных в воде, а затем и на суше потребовало повышение физической активности [20], и совершенствования функции локомоции, необходимой для актив- ного и длительного перемещения в пространстве: поисков пищи, сезонных миграций рыб и млекопитающих, перелётов птиц и насекомых.
Это, в первую очередь, потребовало совершенствования энергетического обеспечения гладких миоцитов и скелетной мускулатуры. Гликогена же скелетных мышц хватает только для покрытия энергетической потребности их кратковременных, и как правило, внезапно возникших нагрузок [10].
Природа же тонуса гладких миоцитов, сходна с природой обычного сократительного акта [21], но отличается низким окислительным обменом и вялым течением анаэробного превращения углеводов [22]. Реализация новых функций мышечной ткани в филогенезе и привела в итоге к необходимости синтеза инсулина, который далее, как полагает [7], создал систему депонирования энергетического субстрата. В связи с этим, основная роль инсулина [7] - энергетическое обеспечение функции локомоции. Для неё инсулин сформировал на мембране гладких миоцитов и мышечных волокон специализированные глюкозные транспортеры (Глют-4), работающие в 6 раз быстрее, чем более древние Глют-1 [7], и депо гликогена. Количество глюкозы, которое можно депонировать в форме гликогена, определяется, в первую очередь, массой мышечной ткани. При этом значительно усиливается перфузия мышц, а число функционирующих капилляров увеличивается в 4-6 раз [20].
Цель исследования - доказать возможность уменьшения жировой массы тела экспериментальных животных при интенсивной физической нагрузке и энтеросорбции.
Материал и методы исследования. Работа выполнена на беспородных белых крысах 4-х месячного возраста массой 160,0 ± 8,0 г (n=15). Животные 1-ой группы (n = 10) выполняли интенсивную мышечную работу динамического характера (плавание). Животные 2-ой группы (п = 5) являлись контрольными.
Плавание осуществлялось в ванне с подогретой до 37°С водой. Находящиеся в воде животные вынуждены были делать быстрые плавательные движения. После плавания животные насухо вытирались и обсушивались в специальном ящике при температуре 37°-38°С. Тренировки проводились ежедневно, в строго определённое время (в 10-12 часов). Длительность нагрузки первого дня эксперимента составляла 1 мин и возрастала через каждые 2 дня ещё на 1 мин. К концу эксперимента (через 75 суток) длительность нагрузки достигала 1 часа. Животные 1-ой и 2-ой групп находились на разнообразном пищевом рационе с добавлением корма «Кitекаt:» [23] и свободным доступом к воде в виварии ветеринарной клиники «Друг». Животным 1-ой группы перорально вводился энторосорбент «Мегамин» ежедневно по 0,5 г в течение 75 суток эксперимента. Животные выводились из эксперимента под эфирным наркозом после импрегнации сосудистого русла солями серебра [24].
Энтеросорбент «Мегамин» представляет собой минеральный препарат, содержащий 50% трибомеханически активированного цеолита [25]. Благодаря своему химическому составу и структуре кристаллической решётки мегамин является хорошим ионообменником и обладает сильно выраженными свойствами адсорбции и абсорбции. Ионы мегамина не поступают из кишечника в общую циркуляцию в больших количествах, но они могут включаться в липидные липосомы и образовывать мембранные системы [26].
Материалом для исследования служили мышцы задних конечностей: камболовидная (m.soleus) и широкая латеральная бедра (m.vastus lateralis), подкожножировая клетчатка, тонкая кишка, жировая капсула почки и большой сальник. Материал фиксировался в 10% растворе нейтрального формалина. Серийные парафиновые срезы толщиной 5-7 мкм окрашивались гематоксилином и эозином, железным гематоксилином но Гейденгайму и азаном. Микрососудистое русло исследуемых объектов импрегнировалось солями серебра [24]. Для изучения гликогена использованы животные 1-ой группы (n = 5) и 2-ой группы (n = 2).
Этим животным осуществлялась интрасосу-дистая фиксация нейтральным фиксатором Ша-бадаша через брюшную аорту с предварительным введением (12%) раствора азотнокислого натрия подогретого до температуры 38-39°С. Парафиновые срезы мышц толщиной 7 мкм приклеивались к предметным стёклам методом сухой наклейки с белком. Цветная реакция на гликоген проводилась по А.Л.Шабадашу [27]. Для контроля использовались срезы, предварительно обработанные амилазы слюны, изучение препаратов проводилось на МБИ-15. Морфометрия микрососудов проводилась на поперечных срезах исследуемых мышц.
Результаты исследования и их обсуждение. Камбаловидная мышца белых крыс в норме фиксирует голеностопный сустав, выполняя при этом сравнительно малонапряженную, но длительную тоническую работу. В связи с этим её мышечные волокна имеют выраженную продольную и поперечную исчерченность, многочисленные ядра и рыхло расположенные миофибриллы. На 1 мм2 мышечной ткани определяется ~ до 3100 капилляров. Гликоген же распределен в мышечных волокнах крайне неравномерно. При этом гранулированный гликоген выявлялся как в межфибриллярной, так и в периферической саркоплазме, а диффузный - как в саркоплазме, так и
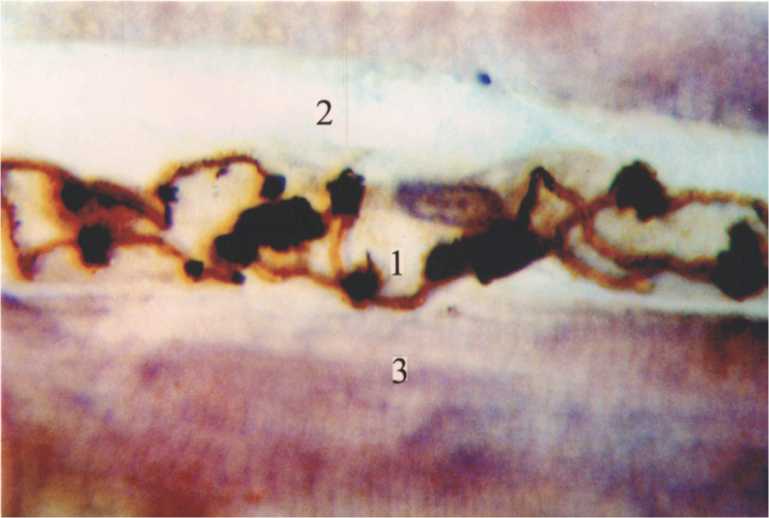
Рис. 1. Расширенный, активно функционирующий гемокапилляр (1) в широкой латеральной мышце (2, 3) белой крысы. 15-е сутки эксперимента. Импрегнация по Ранвье. Докраска гематоксилином. Ув.900.
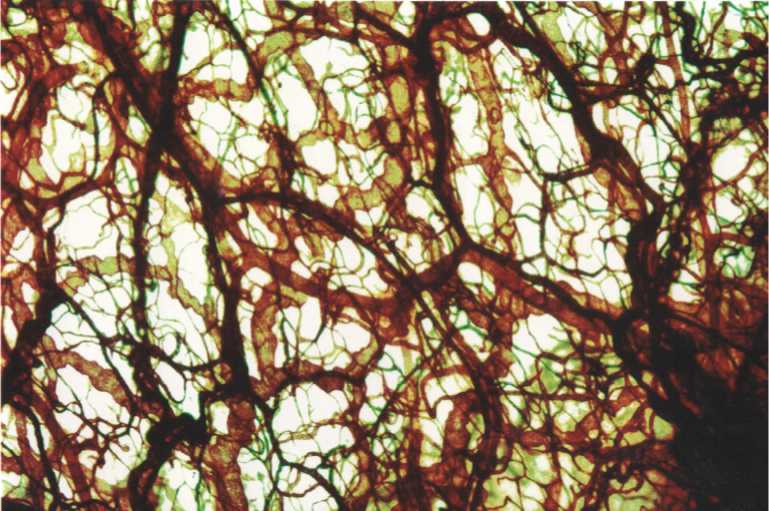
Рис. 2. Микрососудистое русло подслизистой основы тонкой кишки белой крысы. 15-е сутки эксперимента. Импрегнация по Ранвье. Ув.100.
в миофибриллах.
Широкая латеральная мышца у крыс является основным разгибателем коленного сустава. Её сократительный аппарат хорошо развит: многочисленные миофибриллы в мышечном волокне плотно упакованы, а свободная саркоплазма практически отсутствует. Количество капил- ляров на 1 мм2 в широкой латеральной мышце меньше (~ 2700), чем в камбаловидной мышце, но гликогена больше. Его размещение в мионах стабильное и соответствует ходу анизотропных дисков. Динамическая нагрузка усиливает характеристики, свойственные для интактной широкой латеральной мышцы. Так число капилляров на Д
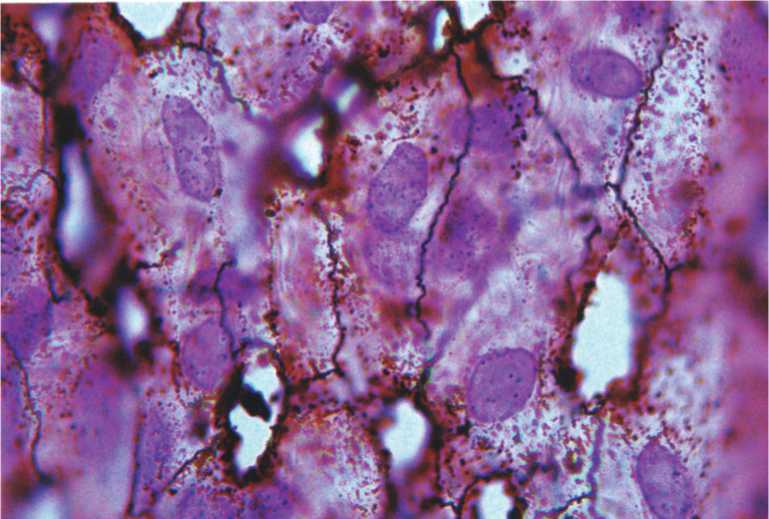
Рис. 3. Высокая транспортная активность эндотелия вены подслизистой основы тонкой кишки белой крысы. Сформированные «стоматы». Импрегнация по Ранвье. Докраска гематоксилином и эозином. Ув.900.
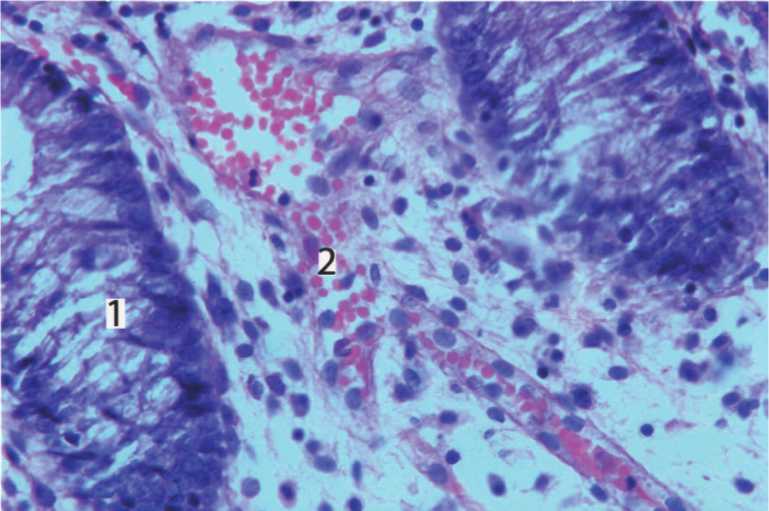
Рис. 4. Собственная пластинка слизистой оболочки тонкой кишки белой крысы. 15-е сутки эксперимента. 1)кишечные железы; 2)микрососуды. Окраска гематоксилином и эозином. Ув.400.
мм увеличивается на 27,7% (~ до 3450), а масса мышцы - на 50% (рис. 1). Все виды физических нагрузок, особенно плавание, сопровождаются новообразованием мышечных волокон и кровеносных капилляров [28, 29]. Морфологические изменения в мышцах, связанные с уменьшением количества быстрых окислительных мышечных волокон II А типа и увеличение количества быстрых гликолитических мышечных волокон II Б типа, а так же нарушение кровоснабжения мышечной ткани, рассматривается как одна из главных причин развития инсулинрезистентности [30].
Одновременно с морфофункциональными изменения в соматической мускулатуре под влиянием физической нагрузки происходили изменения и в жировой ткани. У крыс контрольной группы в интактной жировой ткани (большой сальник, паранефральная жировая капсула) пролиферативная активность адипоцитов была снижена. Синтез ДНК в них практически отсутствовал. После интенсивной физической нагрузки число типичных адипоцитов белой жировой ткани большого сальника резко уменьшалось. Они замеш;ались незрелыми клетками округлой или полигональной формы, содержащими многочисленные жировые капли, окруженные простой мембраной. В пара-нефральной клетчатке основную массу жировой ткани к окончанию эксперимента составляли многокамерные адиноциты. Бурая жировая ткань, сформированная многокамерными адипоцитами, в значительной степени отличается от белой жировой ткани, сформированной однокамерными адипоцитами, как по гистологическому строению и тканевому потенциалу, так и по своему функциональному значению [31, 32]. Зачатки жировой ткани описываются как «первичные органы» у 18-дневных плодов крысы. Многокамерные адиноциты бурой жировой ткани отличаются от недифференцированных клеток соединительной ткани наличием в них глыбок гликогена и линидных капель [33]. В адипоцитах белой жировой ткани гликоген отсутствует [34]. В эксперименте после интенсивной физической нагрузки и в большом сальнике, и паранефральной клетчатке кроме двух типичных форм адипоцитов белой и бурой жировой ткани, появились различные их переходные формы! (рис. 2) Они характеризовались наличием в цитоплазме адипоцитов различного количества неодинаковых но размеру жировых капель. При этом большая часть жировых долек включала в себя адипоциты и белой, и бурой жировой ткани. В этих случаях клетки преобладающего типа располагались в центре, а остальные - на периферии дольки. Таким образом, адипоциты белой жировой ткани способны не только к дедифференцировке и фагоцитозу, но и к трансформации в подвижные макрофаги [31], и, очевидно, и в адипоциты бурой жировой ткани [35].
После приёма мегамина у животных 1-ой группы наблюдалось значительное усиление функциональной активности микрососудистого русла стенки тонкой кишки и большого сальника: увеличение числа функционируюгцих капилляров, увеличение их диаметра, повышение транспортной активности эндотелиоцитов (рис. 3). В цитоплазме эндотелиоцитов увеличивалось число рибосом, зона Гольджи гипертрофировалась, формировались лизосомные структуры.
Повышение функциональной активности микрососудов сопровождалось усилением миграционной активности иммуноцитов и появление расширенных лимфатических микрососудов. В последующие сроки наблюдения - 30, 60, 90 суток, в области куполов лимфоидных бляшек кишки и лимфоидных узелков большого сальника определялось значительное число макрофагов и плазмоцитов. Абсолютное число клеток в одиночных лимфоидных узелках принадлежало В- лимфоцитам. В собственной пластинке слизистой оболочки кишки определялось большое число лимфоцитов и малодифференцированных лимфоидных клеток (рис. 4).
Обсуждая полученные данные и анализируя данные литературы следует отметить, что пищеварительной системе принадлежит особая роль в обеспечении гомеостаза в организме. Прежде всего потому, что «биологический смысл совокупностей пищеварительной системы заключается в образовании плазмы крови, которая в последующем обеспечивает жизнедеятельность всех клеток, органов и организма в целом» [36, с. 187].
Современная концепция постоянства химуса и представления о механизмах, обеспечивающих гомеостатирование энтеральной среды, впервые была сформулирована А.Д.Синещёковым [37].
Морфофункциональной основой энтеросорбции является способность стенки тонкой кишки и её кровеносного русла к транссудации плазмы крови в просвет кишки и к последующей реабсобции в кровеносное и лимфатическое русло уже гоместатированного химуса. Так, свыше 40% липидов, транспортируемых по кишечному лимфатическому стволу, вновь возвращается в тонкую кишку в составе липидов желчи [38]. Одновременно было установлено, что поток веществ, поступающих в процессе пищеварения из просвета тонкой кишки в кровь, близок по своему составу к плазме крови [39, 40].
В связи с этим, количество углеводов в химусе, почти в 10 раз меньше, чем в смешанных рационах. Это обусловлено не только разведением среды эндогенной плазмой крови, но и избирательным всасыванием углеводов: масса их в химусе, поступающем в тонкую кишку, в среднем составляет лишь 19% от массы в рационе [41].
Однако, даже при полном отсутствии углеводов в рационе, в химусе, поступающем в тонкую кишку, глюкоза, хотя и небольших количествах, всё же обнаруживается. Она поступает в него из крови, транспортирующейся в просвет кишки. Наличие постоянного круговорота, т.е. повторного выделения в просвет пищеварительного канала веществ, ранее уже поступивших в общую циркуляцию, установлено для воды, ряда электролитов, мономеров, липидов. При этом липиды выделяются настолько интенсивно, что их масса в химусе возрастает на 173,4 ± 4,7 от их массы в рационе [42, 43]. В сформированном химусе масса фосфолипидов возрастает в 2,4 раза, холестерина - в 4,4 раза, НЭЖК - в 26 раз [44, 45]. Энтеросорбция способствует нормализации липидного метаболизма, приводит к снижению в крови уровня общего холестерина, общих липидов, триглицеридов [46]. Это делает энтеросорбцию «патогенетически обоснованной терапией, направленной на защиту организма от избыточного количества продуктов свободнорадикального окисления, нормализацию метаболизма в миокарде, сохранение его сократительной активности, предупреждение распространения повреждения» [47, 48].
Уже накоплен определённый опыт применения энтеросорбентов в терапии сахарного диабета: получены положительные эффекты компенсации нарушенного углеводного обмена, снижения содержания глюкозы в крови и преодоления толерантности к физической нагрузке [49]. Нарушения липидного обмена, в свою очередь, сопровождаются развитием гипертензии. При этом одновременное сочетание дислипидемии и гипертензии встречается у больных в 1,5 раза чаще, чем следовало ожидать, если бы эти два заболевания развивались независимо [50]. Наиболее частым проявлением нарушений регуляции сосудистого тонуса при дислипидемиях является дисфункция эндотелия [51, 52]. Имеющие в настоящее время данные литературы [53] показывают, что одним из механизмов развития патологических процессов при дислипидемии является вмешательство липопротеидов в нормальные процессы регуляции сосудистого гомеостаза. Частицы липопротеидов выступают в качестве «молекулярных саботажников» выводя из строя отдельные сигналпередающие системы крови и сосудистой стенки. Одновременно липопротеиды проводят «дезинформацию», вызывая активацию систем вторичных посредников там и тогда, где или когда эта активация в норме отсутствуют или вредна [53, 54].
Таким образом, человеком (Homo sapiens) потеряна из важнейших функций сохранения гомеостаза - функция локомоции. При гиподинамии, в состоянии которой находится практически вся человеческая популяция, развивается инсулинорезистентность и недостаточная утилизация глюкозы. В этих условиях в соответствии с функционированием цикла Рэндла основным энергетическим субстратом становятся жирные кислоты, а глюкоза становится токсичной для многих периферических тканей [55].
Дальнейшее развитие событий при этом сценарии приводит к снижению продукции соматотропных гормонов [56], ожирению [57], метаболической иммунодепрессии [56], X - син- дрому, как предвестнику многих неинфекционных заболеваний человека: сахарного диабета 2-го типа, атеросклероза, гипертонической болезни, остеопорозу, инфаркта миокарда, инсульта, канцерогенеза.
Список литературы Интенсивная физическая нагрузка и энтеросорбция уменьшают массу жировой ткани у экспериментальных животных
- Клебанова Е.М. Значение жировой ткани и её гормонов в механизмах инсулиновой резистентности и развитие сахарного диабета - 2-го типа/ Е.М.Клебанова, М.И.Балаболкин // Клин.мед., 2007, № 7, с.20-27.
- Наточин Ю.В. Архитектура физиологических функций: тот же фундамент, новые грани / Ю.В.Наточин // Рос.физиол. журн., 2002, № 2, с.130-143.
- Панков Ю.А. Роль лептина и его белковых медиаторов в нейрофизиологии/Ю.А.Панков // Вестн. РАМН, 2005, № 2, с.44-48.
- Saltiel A.R. Role of adipose tissur as an inflammatory organ in human disrases \ A.R. Saltiel, U.Miller-Lander et al. \ \ Endocr. Rev., 2006, v. 27, p.449-467.
- Панков Ю.А. Новый гормон адипонекти. Его роль в патогенезе сахарного диабета / Ю.А.Панков // Вестн. РАМН, 2006, № 9-10, с.99-103.
- Хэм А., Норман Д. Гистология. Мир. 1989, т. 2, 254 с.
- Титов В.Н. Фундаментальная медицина: функциональная роль инсулина как фактора обеспечения энергией биологической функции локомоции (обзор литературы) / В.Н.Титов // Клин.лаб. диагн., 2005, № 2, с.3-8.
- Марков А.И. Анатомия жировых тел щёк в постнатальном онтогенезе / Дис. канд. мед. наук. Самара, 1994, 120с.
- Марков И.И. Сосудистое русло и иммунные структуры большого сальника / Самара, Изд-во «Саратовский университет», 1992, 105с.
- Климов А.Н., Никульчева Н.Г. Обмен липидов и липопротеидов и его нарушения / СПб. Питер ком, 1999, 513с.
- Панков Ю.А. Жировая ткань как эндокринный орган, регулирующий рост, половое созревание и другие физиологические функции / Ю.А.Панков //Биохимия, 1999, № 6, с.725-734.
- Diez J.J. The role of the novel adipocytederived hormone adiponectin in human disease \\ J.J.Diez, P.Jglesias // Eur. J.Endocrinol., 2003, v. 148, p. 293-300.
- Steppan C.M. The hormone resistin links - odesity to diadetes \\ C.M. Steppan, S.T.Balley et al. // Nature, 2001, v.409, p/307-312.
- Comds T.P. Endogenious glucosae production is inhibited by the adipose - deribed protein Acrp 30 \\ T.P. Comds, A.H.Bergetak. // J.Clin. Jnvest. 2001, b.108, p. 1875 - 1881.
- Shulman G.J. Cellular mechanisms of insulin resistance \\ G.J. Shulman // Clin. Jnvest. 2000, v.106, p.171-176.
- Barfurth D. Verrgleichend-Histochemiche Untesuchungen uber des Glycogen \ D. Barfurth \ Arch F. Micr. Anat., 1885, Bd. 25, S. 269-404.
- Бабурина Е.А. Мускульная клетка ноги Helix pomatia // Арх. анат., № 1, с.76-102.
- Грекова Н.А. К цитологии и цитофизиологии гладкой мускулатуры беспозвоночных / Н.А.Грекова // Автореф. дис.. канд. биол. наук. М. 1951, 23с.
- Шорникова М.В. Сравнительная гистохимия гликогена в гладкой мускулатуре некоторых беспозвоночных и позвоночных животных / М.В.Шорникова // Арх.анат., 1960, № 9, с.44-52.
- Линхард Г.Э. Как клетки поглощают глюкозу / Г.Э.Линхард, Я.У.Скотт/ в Мире наук, 1992, № 3, с.22-28.
- Жуков Е.К. Исследования о тонусе скелетных мышц / М.Медгиз, 1956, 179 с.
- Северин С.Е. Углеводно-фосфорный и окислительный обмен мышц и влияние на него канозина и ансерина / С.Е.Северин // Матер. VIII Всесоюзн. съезда физиол., биохим., фармакол. М. 1955, 191-193.
- Попова Н.К. Влияние мышечной деятельности на потребление мышцами азотсодержащих веществ / Н.К.Попова // Физиол. Журн. СССР, 1951, № 1, с.103-109.
- Марков И.И. Способ исследования микрососудистого русла большого сальника. Авт. свидет. № 1619100 от 08.09.1990г.
- Лелас Т. У. Устройство для динамического тонкого измельчения и микронизации Патент № РСТ 99/00757 от 27.04.99г.
- Pavelic K. Jmmunostimu Latory effect of natural elinoptilolite as a possible mechanism of cts antimetabolicability \\ K. Pavelic, V. Katic // J.Cancer Res. Clin., Oncol., 2002, v. 128, p. 37-44.
- Шабадащ А.Л. Гистохимия гликогена нормальной нервной системы. М.Медгиз, 1949, 213с.
- Видик Т.В. Реактивные изменения поврежденных поперечнополосатых мышечных в обычных условиях и при гиперфункции скелетной мускулатуры /Т.В.Видик // Арх.анат., 1957, № 3, с.88-94.
- Литвер Г.М. К вопросу о возможности и восстановления целых мускулов у крысы из пересаженной мышечной ткани / Г.М.Литвер, Н.Н.Дампель // Арх. анат., 1959, № 9, с.54-65.
- Krotkiewski M. Капиллеризация мышц, их морфология и патогенез метаболического синдрома / M. Krotkiewski// Пробл. эндокринол., 1996, № 4, с.42-46.
- Дмитриева Е.В. Реактивные изменения клеток белой и бурой жировой ткани/ Е.В.Дмитриева // Арх.анат., 1968, № 10, с.37-42.
- Труупыльд А.Ю. О реактивных изменениях ретроперитопеальной бурой жировой ткани у крыс после адреналэктомии / А.Ю.Труупыльд, П.К.Кырге и др. // Арх. анат., 1975, № 9, с.77-83.
- Suter E.R. The ultrastructure of browu adipose tissue in perincetalrats \\ E.R. Suter // Experimentia, 1969, v. 25, p. 286-287.
- Mohr W. Histochemische Untersuchungen am Fettgewebe \\ W. Mohr, G.Beneke // Histochemie, 1969, v. 17, N 3, p. 248-255.
- Tirady C. Conversion from white to brown adipocytes: a strategy for control of fat mass \\ C. Tirady, D.Langin // Trends Endocrinol. Metad., 2003, v.14, N 10, p/439-441.
- Алиев А.А. Липидный обмен и продуктивность животных/ М.Колос, 1980, 381 с.
- Синещёков А.Д. Биология питания сельскохозяйственных животных/М.Колос, 1965, 399 с.
- Алиев А.А. Оперативные методы исследования сельскохозяйственных животных / Л.Наука, 1974, 366 с.
- Гальперин Ю.М. Организация процесса снабжения организма нутриентами в период активного пищеварения/Ю.М. Гальперин //Физиол.журн. СССР, 1985, № 2, с. 182 - 194.
- Гальперин Ю.М. Значение гомеостатированного химуса для всасывания в тонкой кишке /Ю.М.Гальперин, Т.С.Попова // Докл. АН СССР, 1979, № 3, с.801 - 804.
- Гальперин Ю.М. Пищеварение и гомеостаз / Ю.М.Гальперин, П.И.Лазарев// М.Наука, 1986, 303 с.
- Saunder D.R. Insignificance of the enterodiliary circulation of lecithin in man \\ D.R. Saunder // Gastroenterology, 1970, v.59, N 6, p. 848 - 852.
- Уилкинсон А. Водно -электролитный обмен в хирургии / М.Медицина, 1974, 336 с.
- Гальперин Ю.М. Влияние гоместатированного химуса на всасывание его компонентов в тонкой кишке в условиях естественного пищеварения / Ю.М.Гальперин, М.Я.Симонов и др. // Физиол. журн. СССР, 1985,; 5, с. 631 - 645.
- Wilson T.H. Interstinal absorption \ Phylalphia: Sauders, 1962, 263 p.
- Мамай А.В. Энтерособция как средство продления жизни/ А.В.Мамай, В.Н.Крутко // Физиология человека, 1996, № 3, с.131 - 135.
- Пархоменко А.Н. Применение энтеросорбции при ИБС и нарушениях липидного обмена / А.Н.Пархоменко, Е.Р.Понов // Тер.арх., 1999, № 8, с. 88 - 95.
- Давыдов В.И. О возможности использования углеродных энтеросорбентов для нормализации холестеринового обмена / В. И.Давыдов, С.С.Ставицкая и др. // Биохимия, 1994, № 2, с.304 - 317.
- Дониш Р.М. Толерантность больных сахарным диабетом к физической нагрузке / Р.М.Дониш, Д.В.Кириенко и др. // Врач. дело, 1991, № 7, с. 63- 70.
- Eaton C.B. Prevalence of hypertension, deslipidemia, and dyslipidemic hypertension \\ C.B. Eaton, A.R.Assf et al. // J.Jam. Pract., 1994, v.38, N 1., p.17 - 23.
- Harrison D.G. Cellular and molecular ofendothelial cell aysfunction \\ D.G. Harrison // J.Clin, Jnvest., 1997, v. 100, N 9, p. 2153 - 2557.
- Nickenig G. Hypercholesterolemia is associated With enhanced angiotensin AT-1 - receptor expression \\ G. Nickenig, O. Jung et ak. // Amer \\ J.Physol., 1997, v.272. N 6 pt 2, H 2701 - H 2707.
- Бочков В.Н. Влияние липопротеидов на системы передачи регуляторных сигналов в тромбоцитах и клетках сосудистой стенки В.Н.Бочков, В.А.Ткачук // Рос.физиол.журн., 2005, № 1, с.12-30.
- Saini H. K. Role of cholesterol in cardiovascular dysfunction \\ H. K. Saini, A. S. Arneja et al. // Can. J.Cardion., 2004, v.20, N 3, p. 333 - 346.
- Берштейн Л.М. Джокерная роль глюкозы в развитии основных неинфекционных заболеваний человека/ Л.М.Берштейн//Вестн. РАМН., 2005, № 2, с. 48 - 51.
- Дильман В.М. Четыре модели медицины / Л.Медицина, 1987, 437 с.
- Дедов И. И. Сахарный диабет / И. И.Дедов, М.В.Шестаков // М.Медицина, 2003, 217 с.
