Интенсивность фотосинтеза часто используемых древесных растений

Автор: Эркебаев Т.К., Аттокуров К.Ш., Капарова Н.
Журнал: Бюллетень науки и практики @bulletennauki
Рубрика: Биологические науки
Статья в выпуске: 12 т.8, 2022 года.
Бесплатный доступ
Фотосинтез является жизненно важным источником энергии в природе. Разные методики определения интенсивности фотосинтеза у растений позволяют однозначно сопоставить все имеющиеся экспериментальные данные и общие выводы об особенностях фотосинтеза у растений. Цели исследования: определение интенсивности фотосинтеза у древесных растений. Сбор данных по интенсивности фотосинтеза изученных видов растений на первом и втором году и определение влияния внешних факторов температуры, интенсивности света, концентрации CO2 и O2 в воздухе, влажности воздуха и почвы на процесс фотосинтеза. Результаты исследования: низкая интенсивность фотосинтеза отмечается у видов, произрастающих в областях с умеренным или мягким климатом. Выводы: высокой интенсивностью фотосинтеза обладают виды растений, обитающие в основном в областях с континентальным климатом или в горных условиях.
Фотосинтез, древесные растения, факторы окружающей среды, плодовые культуры, измерение
Короткий адрес: https://sciup.org/14126151
IDR: 14126151 | УДК: 581: | DOI: 10.33619/2414-2948/85/14
Photosynthesis intensity in frequently used woody plants
Photosynthesis is a vital source of energy in nature. Various methods for determining the intensity of photosynthesis in plants make it possible to unambiguously compare all available experimental data and general conclusions about the features of photosynthesis in plants. Research objectives: determination of the intensity of photosynthesis in woody plants. Collection of data on the intensity of photosynthesis of the studied plant species in the first and second years and determination of the influence of external factors of temperature, light intensity, CO2 and O2 concentrations in the air, air and soil humidity on the photosynthesis process. Research results: low intensity of photosynthesis is observed in species growing in areas with a temperate or mild climate. Conclusion: plant species that live mainly in areas with a continental climate or. in mountainous conditions have a high intensity of photosynthesis.
Текст научной статьи Интенсивность фотосинтеза часто используемых древесных растений
Бюллетень науки и практики / Bulletin of Science and Practice
При изучении интенсивности фотосинтеза многими авторами отмечается видоспецифичность этого показателя у различных растений [1–4]. В природных условиях эти видовые различия проявляются в большей или меньшей степени в зависимости от внешних условий. Интенсивность фотосинтеза определяли по методу Л. А. Иванова и И. Л. Косовича [5], основанного на титрометрическом определении количества СО 2 , поглощенного листьями при фотосинтезе. Количество СО 2 выражалось в мг/дм2 час.
Площадь поверхности листа определяли весовым методом [5], площадь поверхности хвои по формуле М. Я. Оскреткова [6, 7].
Определение интенсивности фотосинтеза проводили на деревьях, произрастающих в Ботаническому саду АН РУз. Опыты проводились в трехкратной повторности с мая по сентябрь (15–25 числа), в утренние часы (730–900), в безоблачную погоду при t = 20–28°С в течение года. Листья и хвоя брались из среднего яруса кроны.
Материалы и методы исследования
По максимальному значению интенсивности фотосинтеза, изученные нами видны по результатам первого года, были разделены на три группы.
Первая группа состоит из видов с высокой интенсивностью фотосинтеза, ассимилирующие больше 8 мг СО 2 /дм2 час.
Вторая группа включает виды со средней интенсивностью фотосинтеза, ассимилирующие от 6 до 8 мг СО 2 /дм2 час.
Третья группа включает виды с низкой интенсивностью фотосинтеза, их показатель меньше 6 мг СО 2 /дм2 в час.
Результаты и обсуждение
В Таблице 1 и 2 приведены данные по интенсивности фотосинтеза изученных нами видов на первом и втором году. При сравнении результатов двух лет видно, что на первом году у всех видов, за исключением Pinus pallasiana D. Don, показатели были выше, хотя достоверные различия наблюдались только у 5 видов: Robinia pseudoacacia L., Catalpa speciosa (Warder ex Barney) Warder ex Engelm., Quercus robur L., Fraxinus excelsior L. и Acer campestre L. (t> 2,75).
Таблица 1
ИНТЕНСИВНОСТЬ ФОТОСИНТЕЗА ИССЛЕДОВАННЫХ ВИДОВ РАСТЕНИЙ мг СО 2 /дм2 час (первый год)
|
Виды |
май |
июнь |
июль |
август |
сентябрь |
|
Juglans regia L. |
5,24±0,36 |
16,41±1,01 |
14,11±0,98 |
14,04±0,91 |
7,71±0,45 |
|
Juniperus virginiana L. |
4,47±0,30 |
11,39±0,72 |
9,65±0,63 |
6,27±0,40 |
3,11±0,28 |
|
Betula pendula Roth |
4,64±0,28 |
10,68±0,6 |
7,68±0,50 |
4,19±0,27 |
4,87±0,30 |
|
Fraxinus excelsior L. |
9,91±0,56 |
9,70±0,53 |
6,83±0,55 |
6,56±0,35 |
3,46±0,23 |
|
Robinia pseudoacacia L. |
9,57±0,72 |
9,72±0,61 |
9,59±0,62 |
8,97±0,68 |
6,69±0,59 |
|
Platycladus orientalis (L.) Franco |
5,87±0,42 |
8,37±0,63 |
5,56±0,39 |
4,03±0,24 |
3,53±0,19 |
|
Ulmus pumila L. |
4,33±0,29 |
5,59±0,35 |
7,84±0,42 |
6,59±0,45 |
4,43±0,26 |
|
Catalpa speciosa (Warder ex Barney) Warder ex Engelm. |
7,62±0,48 |
5,80±0,37 |
5,75±0,42 |
4,12±0,32 |
2,38±0,15 |
|
Quercus robur L. |
4,79±0,32 |
7,57±0,45 |
5,96±0,31 |
4,40±0,27 |
2,01±0,13 |
|
Cercis canadensis L. |
6,90±0,45 |
4,90±0,31 |
3,06±0,21 |
3,90±0,29 |
2,70±0,20 |
|
Gleditsia triacanthos L. |
3,60±0,24 |
6,69±0,38 |
5,89±0,32 |
6,70±0,37 |
2,12±0,17 |
|
Styphnolobium japonicum (L.) Schott |
4,40±0,31 |
5,92±0,34 |
5,67±0,42 |
5,48±0,36 |
5,46±0,40 |
Бюллетень науки и практики / Bulletin of Science and Practice Т. 8. №12. 2022
|
Виды |
май |
июнь |
июль |
август |
сентябрь |
|
Acer campestre L. |
5,91±0,42 |
5,02±0,55 |
4,53±0,30 |
3,46±0,28 |
0,70±0,04 |
|
Albizia julibrissin Durazz. |
3,40±0,20 |
5,45±0,35 |
4,45±0,27 |
3,32±0,26 |
3,91±0,29 |
|
Pinus pallasiana D. Don |
3,42±0,25 |
5,46±0,37 |
5,03±0,33 |
4,45±0,22 |
2,34±0,15 |
|
Aesculus hippocastanum L. |
4,62±0,27 |
3,91±0,25 |
3,83±0,28 |
3,77±0,30 |
1,64±0,12 |
|
Populus nigra L. |
3,12±0,21 |
4,42±0,29 |
4,11±0,14 |
3,08±0,19 |
2,81±0,11 |
|
Platanus orientalis L. |
2,58±0,81 |
3,74±0,21 |
3,60±0,24 |
2,74±017 |
2,43±0,15 |
|
Salix alba L. |
3,18±0,23 |
3,49±0,30 |
3,12±0,18 |
3,11±0,21 |
2,90±0,12 |
|
Prunus domestica L. |
11,5±1,0 |
9,65±0,64 |
7,44±0,60 |
5,72±043 |
|
|
Malus sylvestris Mill. |
7,91±0,66 |
10,73±0,81 |
13,56±1,31 |
9,43±0,72 |
|
|
Prunus armeniaca L. |
9,42±0,72 |
8,73±0,73 |
6,42±0,54 |
5,32±0,41 |
|
|
Prunus subgen. Cerasus (Mill.) A. |
8,33±0,61 |
6,34±0,41 |
5,73±0,42 |
5,43±0,25 |
Gray
Таблица 2
ИНТЕНСИВНОСТЬ ФОТОСИНТЕЗА ИССЛЕДОВАННЫХ ВИДОВ РАСТЕНИЙ, мг CO 2 /дм2 час (второй год) t 0,05 ≥ 2,78
|
Виды |
май |
июнь |
июль |
август |
сентябрь |
t |
|
Juglans regia L. |
6,86±0,36 |
14,56±0,97 |
13,29±1,0 |
12,86±1,10 |
7,45±0,62 |
1,34 |
|
Juniperus virginiana L. |
5,45±0,20 |
9,51±0,50 |
7,54±0,52 |
7,45±0,43 |
5.63±0,33 |
2,16 |
|
Betula pendula Roth |
5,56±0,37 |
9,40±0,62 |
7,40±0,51 |
6,85±0,36 |
3,64±0,23 |
1,43 |
|
Fraxinus excelsior L. |
4,57±0,13 |
6,57±0,35 |
5,61±0,33 |
5,14±0,47 |
3,41±0,25 |
4,5 |
|
Robinia pseudoacacia L. |
7,85±0,42 |
6,00±0,30 |
6,93±0,37 |
6,86±0,41 |
4,40±0,26 |
2,94 |
|
Platycladus orientalis (L.) Franco |
3,87±0,28 |
6,93±0,48 |
5,97±0,42 |
4,16±0,28 |
3,21±0,19 |
1,82 |
|
Ulmus pumila L. |
3,57±0,30 |
6,05±045 |
6,85±0,40 |
4,47±0,28 |
4,57±0,32 |
1,60 |
|
Catalpa speciosa (Warder ex Barney) Warder ex Engelm. |
5,28±0,31 |
5,06±0,38 |
4,57±0,23 |
2,87±0,18 |
1,96±0,13 |
4,10 |
|
Quercus robur L. |
2,64±0,15 |
5,54±0,34 |
4,92±0.33 |
3,15±0,21 |
2,43±0,15 |
3,63 |
|
Cercis canadensis L. |
5,14±0,40 |
4,40±0,33 |
4,44±0,23 |
2,96±0,17 |
2,50±0,1 |
2,43 |
|
Gleditsia triacanthos L. |
2,76±0,16 |
5,81±0,39 |
4,75±0,30 |
5,46±0,40 |
4,44±0,36 |
1,62 |
|
Styphnolobium japonicum (L.) Schott |
4,40±0,26 |
5,43±0,32 |
5,05±0,39 |
5,43±0,42 |
2,50±0,17 |
1,04 |
|
Acer campestre L. |
4,14±0,29 |
3,78±0,20 |
3,56±0,23 |
3,14±0,16 |
2,10±0,15 |
3,47 |
|
Albizia julibrissin Durazz. |
3,50±0,23 |
5,05±0,40 |
4,06±0,31 |
4,46±0,23 |
3,35±0,23 |
0,74 |
|
Pinus pallasiana D. Don |
3,98±0,30 |
5,45±0,39 |
4,97±0,35 |
4,45±0,31 |
4,40±0,2 |
0,00 |
|
Aesculus hippocastanum L. |
4,28±0,24 |
3,65±0,19 |
3,28±0,15 |
3,15±0,22 |
1,40±0,10 |
0,97 |
|
Populus nigra L. |
2,85±0,15 |
4,65±0,29 |
4,20±0,31 |
3,43±0,15 |
2,92±0,11 |
0,57 |
|
Platanus orientalis L. |
2,82±0,21 |
3,43±0,24 |
3,14±0,21 |
3,14±0,30 |
3,17±0,19 |
0,31 |
|
Salix alba L. |
2,67±0,19 |
2,92±0,20 |
2,28±0,15 |
2,05±0,12 |
1,83±0,13 |
1,58 |
|
Prunus domestica L. |
10,5±1,1 |
9,0±0,7 |
6,25±0,5 |
3,50±0,3 |
— |
0,80 |
|
Malus sylvestris Mill. |
10,69±0,50 |
11,42±0,90 |
12,87±0,91 |
9,11±0,62 |
— |
0,44 |
|
Prunus armeniaca L. |
8,31±0,60 |
7,42±0,50 |
5,91±0,41 |
5,05±0,41 |
— |
0,21 |
|
Prunus subgen. Cerasus (Mill.) A. Gray |
8,11±0,70 |
6,43±0,60 |
4,92±0,52 |
3,43±0,13 |
— |
0,23 |
Примечание: t — достоверность различий максимальных значений первого и второго года
Причины наблюдаемых расхождений мы видим, прежде всего, в разнохарактерности внешних факторов таких как температуры, интенсивности света, концентрации CO2 и О2 в воздухе, влажности воздуха и почвы, которые, как установлено, оказывают самое непосредственное влияние на процесс фотосинтеза [1, 8–10]. Так, весна второго года была необычайной влажной. Количество осадков за февраль-май второго года составило 474 мм, тогда как первом году всего 269 мм.
Все исследованные плодовые культуры обладают высокой интенсивностью фотосинтеза (больше 8 кг СО 2 /дм2 час).
Неодинаковые методики определения интенсивности фотосинтеза, используемые различными исследователями, не позволяют однозначно сопоставить все имеющиеся экспериментальные данные, но самые общие выводы об особенностях фотосинтеза растений все же сделать можно. Известно, что наиболее низкая интенсивность фотосинтеза характерна для видов, произрастающих в областях с мягким климатом, тогда как с усилением континентальности климата интенсивность фотосинтеза возрастает, достигая наибольших величин в пустынях и в высокогорьях [2].
В результате наших исследований эта закономерность проявляется довольно четко. Так, низкая интенсивность фотосинтеза в основном отмечается у видов, произрастающих в областях с умеренным или мягким климатом. Из видов третьей группы Aesculus hippocastanum L. распространен на Балканском полуострове, Acer campestre L. в Крыму и на Кавказе, Platanus orientalis L. в Средиземноморье (Балканский полуостров), Albizia julibrissin Durazz. и Pinus pallasiana D. Don на Кавказе.
С другой стороны, большинство видов первой и второй групп распространены в областях с более и менее континентальным характером климата или в горных условиях. Исследованные виды различаются также и по сезонной динамике интенсивности фотосинтеза. Как видно из Рисунков 1–3, у всех видов этот показатель имеет вид одно- или двухвершинной кривой, но пик интенсивности фотосинтеза у различных видов приходится на разные месяцы. По этому признаку изученные виды можно разделить на три группы: Первая группа — это виды, у которых пик приходится на май Aesculus hippocastanum L., Acer campestre L., Fraxinus excelsior L., Catalpa speciosa (Warder ex Barney) Warder ex Engelm. и Cercis canadensis L. (Рисунок 1).

Рисунок 1. Aesculus hippocastanum L.

Вторая группа состоит из видов, у которых максимальная интенсивность фотосинтеза наблюдается в июне: Quercus robur L., Betula pendula Roth , Juglans regia L. , Platycladus orientalis
(L.) Franco , Albizia julibrissin Durazz., Juniperus virginiana L. , Pinus pallasiana D. Don , Biota falcata Lindl., Populus nigra L. и Salix alba L. (Рисунок 2).

Рисунок 2. Quercus robur L.

Третья группа — виды с неясно выраженным пиком интенсивности фотосинтеза: Sophora japonica , Robinia pseudoacacia и Gleditsia triacanthos (Рисунок 3).

Рисунок 3. Styphnolobium japonicum (L.) Schott
Интересно, что виды первой группы, кроме Catalpa speciosa, являются теневыносливыми растениями, тогда как входящие во вторую группу светолюбивыми. Таким образом, среди изученных нами пород светолюбивые виды проявляют максимальную способность к фотосинтезу в период высокой интенсивности света в июне. Наши наблюдения подтверждают данные о том, у светолюбивых растений насыщение световой кривой фотосинтеза наступает при более высокой интенсивности света и мало изменяется при изменении условий освещения. Этим, по-видимому, объясняется то, что, несмотря на разнохарактерность внешних условий первого и второго года виды второй группы, за исключением Quercus robur не имеют достоверных различий по интенсивности фотосинтеза (t = 0,31–2,16), т. е. они мало реагируют на изменение факторов внешней среды (освещение).
В то же время у видов первой группы, за исключением Aesculus hippocastanum L., интенсивность фотосинтеза первом годы достоверно отличалась от данных второго года (t > 2,45), что подтверждает данные о большей лабильности теневыносливых растений [11].
Сезонная динамика интенсивности фотосинтеза у видов группы растений за первый год приведены на Рисунках 4–6.
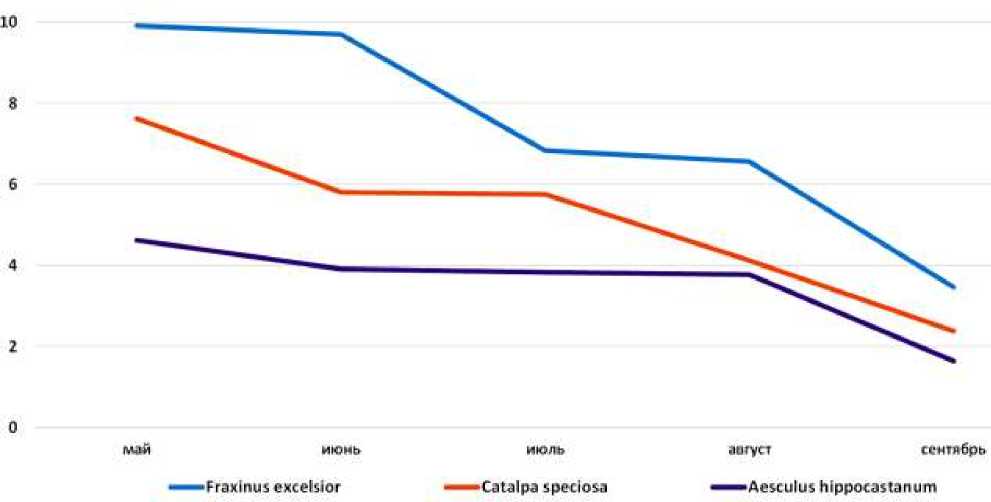
Рисунок 4. Сезонная динамика интенсивности фотосинтеза у видов первой группы, первый год
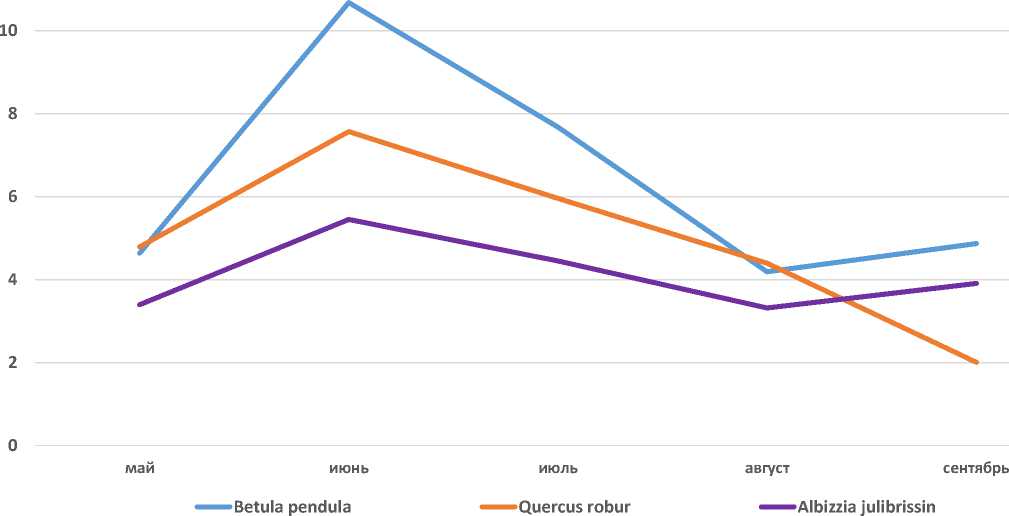
Рисунок 5. Сезонная динамика интенсивности фотосинтеза у видов второй группы, первый год
Необходимо отметить, что полученные данные надо принимать лишь в качестве сравнительных показателей, не рассматривая их как абсолютные величины, поскольку мы не учитывали всех факторов, влияющих на интенсивность фотосинтеза (концентрация CO2, О2 в воздухе, интенсивность света, температура, влажность, почвы, воздуха и др.).
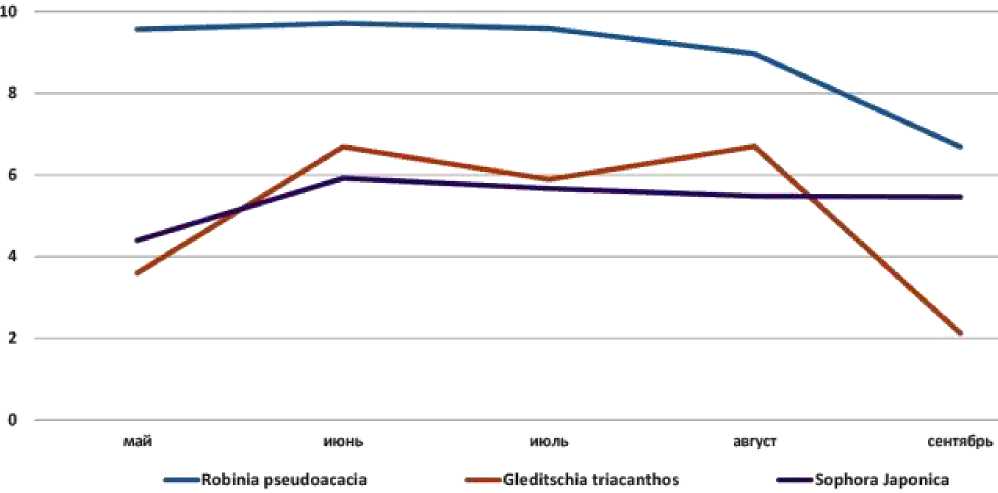
Рисунок 6. Сезонная динамика интенсивности фотосинтеза у видов третьей группы, первый год
Выводы
Таким образом, среди исследованных видов высокой интенсивностью фотосинтеза обладают виды, обитающие в основном в областях с континентальным климатом или в горных условиях Juglans regia L., Juniperus virginiana L., Betula pendula Roth, Robinia pseudoacacia L., Fraxinus excelsior L. и Biota falcata Lindl. Средней интенсивностью фотосинтеза характеризуются Ulmus pumila L., Catalpa speciosa (Warder ex Barney) Warder ex Engelm., Quercus robur L., Cercis canadensis L. и Gleditsia triacanthos L., низкой интенсивностью Platycladus orientalis (L.) Franco, Acer campestre L., Albizia julibrissin Durazz., Pinus pallasiana D. Don, Aesculus hippocastanum L. и Platanus orientalis L.
Полученные результаты подтверждают данные о том, что интенсивность фотосинтеза и ее проявление во многом определяется климатическими особенностями природных местообитаний видов [2].
Список литературы Интенсивность фотосинтеза часто используемых древесных растений
- Рабинович Е. Фотосинтез. М.: Изд-во иностр. лит., 1951-1959. 3 т.
- Заленский О. В. Фотосинтез растений в естественных условиях // Вопросы ботаники. М., 1954. С. 59-87.
- Заленский О. В., Семихатова О. А., Вознесенский В. Л. Методы применения радиоактивного углерода С14 для изучения фотосинтеза. М.; Л.: Изд-во Акад. наук СССР, 1955. 91 с.
- Ничипорович А. А. Световое и углеродное питание растений - фотосинтез. М.: Изд- во Акад. Наук СССР, 1955. 288 с.
- Викторов Д. П. Малый практикум по физиологии растений. М.: Высшая школа, 1969. 120 с.
- Молчанов А. А. Лес и окружающая среда. М.: Наука, 1968. 247 с.
- Смирнов В. И., Кожевников В. С., Гаврилов Г. М. Охрана окружающей среды при проектировании городов. Л.: Стройиздат, 1981. 168 с.
- Пономарева М. М. Влияние концентрации углекислоты на интенсивность фотосинтеза // Труды Ботанического института им. В. А. Комарова. 1960. Т. 4. С. 54-71.
- Глаголева Т. А. Зависимость фотосинтеза памирских растений от интенсивности освещения // Труды Ботанического института им. В. А. Комарова АН СССР. 1963. Т. 4. С. 91-112.
- Логинов М. А. О потенциальном фотосинтезе растений некоторых типов растительности Таджикистана: автореф. … канд. биол. наук. Душанбе, 1963. 17 с.
- Цельникер Ю. Л. Исследования по физиологии растений в лесу. М.: Наука. 1978. 215 с.
