Intracellular electrophysiological evidence: how pain is experienced by neurons

Author: Orlov Valery I., Shikhlyarova Alla I.
Journal: Cardiometry @cardiometry
Section: Original research
Article in issue: 17, 2020.
Free access
Aims. The aim hereof is to identify changes in the performance of the CNS neurons under the prolonged action of low-threshold pain factor according to indicators of the membrane potential level, parameters of the action potential and the pattern of the firing activity distribution of the neurons. Materials and methods. As a model, functionally active neurons of the central nervous system in the grape snail Helix pomatia were selected as an integral part of the organism in vivo. Used has been the author’s original electrophysiological method of continuous recording of intracellular biopotentials of animal neurons both under the background conditions and by applying a long-term low-threshold action (LTA) to the main nerves due to a graded pressure to produce a pain effect of the subthreshold level that over time turns into increasing pain. Results. For the first time in world science, objective evidence for the long-term low-intensity action of pain on the electrophysiological characteristics in a CNS neuron has been obtained. Revealed are the changes in the amplitude and the action potential firing rate, the membrane potential level and the pattern of the neuron pulse activity, traced continuously, up to full neuron potential turning-off. It has been detected that, when removing the pain factor, there are some points of return to the norm available, and, in case of longevity of the pain action, the functional state of the neuron passes the point of no return, after which the neuron electrophysiological potentials reach zero value, indicating the cessation of its life. Conclusions. The dynamics of the neuron functional state in vivo, under prolonged action of the pain factor, reflects nonlinear changes in the membrane potential, firing rate and amplitude of AP. This characterizes the phase process of forming a pathologically stable pain state of the neuron, despite the readiness of the membrane mechanisms at the transition points to a response aimed at returning to life. The evidence obtained in our experiments provides more insight into the mechanisms of chronic neurogenic pain in experimental and clinical medicine, as well as the diagnostics of the brain death.
Neuron, cardiomyocyte, membrane potential, firing rate, low-intensity stimulation, pain
Short address: https://sciup.org/148311480
IDR: 148311480 | DOI: 10.12710/cardiometry.2020.17.821
Text of the scientific article Intracellular electrophysiological evidence: how pain is experienced by neurons
Valery I. Orlov, Alla I. Shikhlyarova. Intracellular electrophysiological evidence: how pain is experienced by neurons. Cardiometry; Issue 17; November 2020; p.8-21; DOI: 10.12710/cardiometry.2020.17.821; Available from:
Biological systems can exist only within certain limitations of environmental parameters, which have specified minimum and maximum values that are compatible with the life of a given biological system [1]. The rhythm and intensity of the biological system response change in accordance with the quality and strength of a stimulus due to the mechanisms of internal plasticity of neurons [2, 3]. If the value of the stimulus is within the physiological limits, has sufficient intensity and is short-lived, then upon completion of the stimulus action, the system issues commands aimed at an appropriate regulation, or compensation for changes produced by the stimulus [4].
It is known that short strong stimuli, considerably exceeding the threshold values, or very long-term stimulation are incompatible with the physiological norms of a living organism; they cause not only local damage, but may also lead to the death of a biological individual. A weak, near-threshold or ultra-weak, below the threshold level, stimulus has been initially considered by physiologists as safe, as a state of rest, restoration of the normal functional state of the biological system. Subthreshold stimuli are those which in acting on the tissue do not produce visible changes, but however are accompanied by certain physical and chemical shifts, while the degree of their changes is not sufficient to initiate excitation. An example of such a subthreshold action may be pain occurring with the development of a tumor process: initially imperceptible, but, over time, gaining a factor of the strong action.
Since the object of research in our experiments is represented by the neurons, it is appropriate to recall the general property of temporality, common to all cells in an organism, and the concept of "chronaxie" introduced by French physiologist Louis Lapic in 1909 [10]. In a diagram shown below (Figure 1 herein), we have marked pink a specific zone that is located outside the area of the cell's response to a stimulus of different modality, the zone of insensitivity, areactivity. It may be assumed that the neurons within the pink zone do not respond to short-term stimuli of low intensity. However, constant or long-acting stimuli or environmental factors, the parameters of which are within this zone, produce cell responses, so that the cells either die or experience their evolutionary adaptation. So, known are microorganisms that live at high temperatures in the ocean in a zone of the constant action of volcanic hot lava, permanently escaping from magma chambers below the Earth surface. All organisms living on the Earth have adapted to the action of the
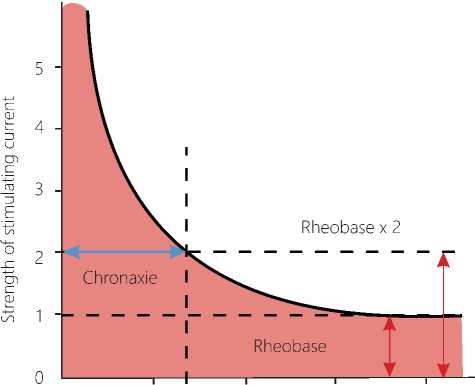
Figure 1. Flowchart of the present study population
Duration of stimulation
Figure 2. Smile of the Life
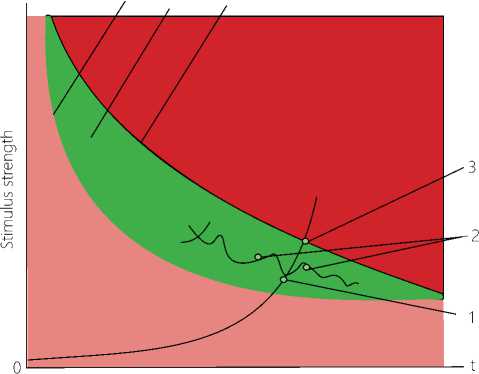
Legend: Pink color zone: action of subthreshold low-intensity stimuli on biosystem. 4. Limit of the irritation threshold. Red color zone: action of dangerous stimuli, which damage the biosystem and are higher than the maximum permissible values. 6. Limit of the maximum permissible levels of the stimulus. 5. Green color zone "Smile of the Life", the zone of the normal life activity, the zone of adequate environmental stimuli for a living system, permissible zone of life from the point of view of electrophysiology. 1. The curve of the action of a long-term subthreshold stimulus. 2. The curves of return points. 3. The point of no return. The scale and limits of the "Smile of the Life" zone have their own values for each biosystem.
Earth's permanent magnetic field, the cosmic background radiation, ultraviolet and other electromagnetic radiation located within some specific limits of the pink zone.
Is it possible to assume that under certain conditions, the pink region can be dangerous for neurons? It
Issue 17. November 2020 | Cardiometry | 9
is accepted that low-intensity, subthreshold, stimulus of any modality cannot immediately produce a response by a biocell, but the long-term, permanent action of the stimulus contributes to cumulative effects: complex tuning of biological systems in the organism, sensitization and gaining values exceeding the stimulus threshold. In this case, the constant action of such a factor (chronization) can lead to deep and significant disturbances in the performance of a cell and cell systems. For example, the term "silent death" is used to refer to radiation exposure. A person being in a radiation zone does not immediately receive alarm signals from his/ her sensory system. Ultraviolet radiation is either not sensed or perceived as a light heat, but it causes damage to skin and vision in case of its prolonged exposure.
To verify our concept, provide ease in use and make a vivid presentation of the process phases in the neuron, we have constructed a new diagram that includes another curve to indicate the limit of the maximum permissible levels of the stimulus (see Figure 2 herein). It should be noted that the pink color zone shows the action of subthreshold, low-intensity stimuli exerted on a biosystem. The red color zone located near the OY axis, “the strength of the stimulus", is the region of high-intensity stimulation that operate for a short time. These are short-term ionizing radiation of high intensity destroying the biosystem, the action of vacuum, high pressure, acceleration, temperature, etc., in other words, the action of dangerous stimuli exceeding the maximum permissible limits that damage the biosystem. The green zone is an area of the normal life activity, and we have given it the name "Smile of the Life” (SL). This green zone being a zone of adequate environmental stimuli for a living system is a permissible region of life in the context of physiology.
Thus, while the red zone located outside the "Smile of the Life" has a clear detrimental effect on the biosystem, the pink region located near the OX axis seems to be safe. However, it is just the zone of chronization, i.e. the long-term action of subthreshold stimuli of low intensity, which over time can cause sensibilization of the organism's systems, and, further, act as a damaging factor, similar to the long-term effect of the pain factor with its biochemical and neurotic disorders in the organism. It is just the region of low-intensity, long-term stimuli of the subthreshold level acting on the nervous system and neurons, where adaptive changes at certain points of phase transitions in a self-organizing living system, with a high degree of probability, can be de- 10 | Cardiometry | Issue 17. November 2020
tected, and our paper is designed to address the above critical points and their role.
The aim of our research work is to identify changes in the functional state of the CNS neurons under the prolonged action of the low-threshold pain factor according to indicators of the membrane potential level, the action potential parameters and the pattern of the firing activity of the neurons.
Materials and Methods
The pain model, herein further referred to as the low-threshold action (LTA), is a modification of the “Method of preparation..." [11], improved by the authors for the purpose of this research. During four years, 93 experiments have been conducted on this research topic, and 32 experiments with the observation of the background state of the neurons among them. It has been shown that the technique provides the desired observation of the background active neuron for 3-4 days [12]. The rest of the experiments have been conducted using LTA, 61 experiments in total, including 34 experiment with recording LTA at the point of return followed by further tracing of the functional state of the neurons, and 27 experiments with LTA continuing up to the disappearance of intracellular electrophysiological potentials in the neurons and recording their state up to 5 hours after their termination. The maximum duration of the experiments using LTA has not exceeded 2 days. The criterion to finish an experiment with the above model has been a stable functional state of the neuron recorded within 3-5 hours after passing the point of return or no-return.
Object of research
In our work, we were guided by the document regulating the conduct of experiments on animals "Rules for conducting research using experimental animals”, Appendix to Order No. 755 “Measures for Further Improvement of Organizational Forms of Work with the Use of Experimental Animals”, dd. August, 12, 1977, issued by the USSR Ministry of Health. We note that the animals we have used are invertebrates which are beyond the scope of the relevant international regulatory documents for working with experimental animals. The neurons of the central nervous system studied in our experiments have been found in vivo in the grape snail Helix pomatia. In all experiments, in order to minimize the impact on the nervous system, we have used recording of only one identified command neu- ron RPaG3. This neuron and its properties have been previously well studied (see Figure 5 herein). A low-intensity, subthreshold level, pain effect has been produced by applying graduated pressure (Figure 3. Item 3 herein) to four main nerves (Figure 3. Item 4 herein) extending from the dorsal surface of the suboesopha-gal ganglion to the visceral sac in the snail Helix poma-tia. For this purpose, we have used Fresnel hairs of the selected diameter, which provide a constant weak level of the nerve pressure applied after switching on the LTA mode in the experiment. The pressure application mode was managed in a gradual manner. 10 minutes after the pressure plates had occupied their fixed position, the hardware check-up of the immutability of the recorded electrophysiological data has been performed before and after switching on the LTA mode.
It is generally recognized that, despite the diversity of the nervous system design and the variety in its complexity, every nervous system is governed by the same general laws at the level of the morphological, physiological and biochemical activity of the nervous cell, the intra-neuronal organization of procedures and mechanisms responsible for transmitting the required data [13].
We have recorded and analyzed the following electrophysiological parameters: membrane potential (MP), action potential (AP) and firing rate (FR) of intracellular biopotentials of neurons using an ultra-thin glass microelectrode filled with electrolyte. The animal was placed on a special substrate made of a composite material developed and designed by the Research Institute of Chemical Industry for the purpose of research of the brain performance. Additional devices have been employed to maintain the viability of the animal, the specified comfortable temperature and the supply of saline solution with nutritional supplements at a strictly constant flow rate. The obtained experimental data have been processed using the Statistics 10 software.
Results and discussion
The initial state. After operating impacts on the organism, the neuron’s FS normalizes within 3-4 hours. During this period of time, the operating level of MP is set, which is relatively stable and changes under the influence of data communication between the neurons. The established background state of the neuron is characterized by a spontaneously changing FR, i.e., the generation of action potentials, which depends on
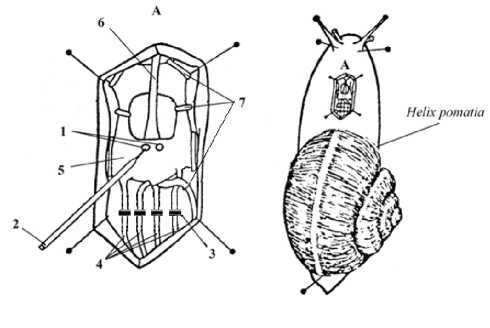
Figure 3. Preparation technique according to Orlov V. I. (1988) Fragment A. 1 - neurons; 2-microelectrode; 3 - device for graded application of pressure to nerve; 4 - four main nerves of the suboesophagal ganglion dorsal side; 5 – dorsal part of the suboesophagal ganglion; 6 – aorta; 7 – fixing, but not pressing brackets. The esophagus is not shown in the figure.
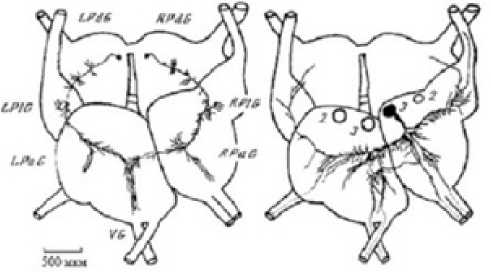
Figure 4. Diagram of processes of the RPaG3 command neuron and the RPDG4 and LPDG4 modulating neurons upon staining with introducing cobalt ions into the neurons. Modulating neurons of the pedal ganglion have branching endings in the area of the branching processes of RPaG3, which extends branches to the main nerves.
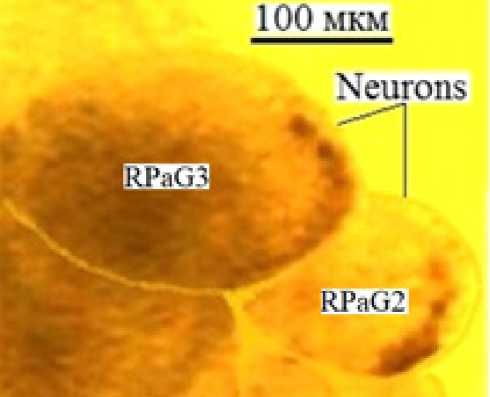
Figure 5. Giant neurons RPaG3 and RPaG2 in the snail Helix pomatia CNS in vitro. Photo by Shikhlyarova A. I.
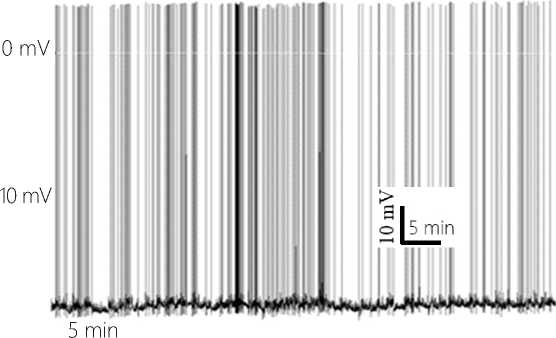
Figure 6. Natural background activity of the neuron 8.5 hours after insertion of the ME tip into the neuron. The 60-minute fragment of the recording.
the changing supply of synaptic pulses associated with the activity of the neurons in the network and their own feedback pulses generated at the dendritic tree of the neuron. The background state of the MP level is additionally determined by the small-amplitude, endogenously generated by the neuron, hyperpolarization and depolarization waves, properly reducing or increasing the probability of changes in FR (see Figure 6 herein). The established background state of the neuron activity has been observed during 5-6 hours.
After establishing the stable mode of the neuron performance, LTA has been turned on. To understand the nature of phase transitions in the neuron functional state (FS), we have attempted to structure them into separate stages that conditionally correlate with the level of pain in accordance with the known laws of electrophysiology, psychophysiology of the sensory systems and image perception, the realization of which is performed by the functional state of the nervous system and the neurons as its structural units.
Neuron activity and pain levels under the LTA conditions: Stage 1. No pain is experienced. The neuron’s activity is equivalent to its background state. (The level of consciousness in human individuals is adequate to the environment). Switching on the LTA mode does not change the nature of the electrophysiological parameters in the neuron background state immediately upon switching-on LTA and later on for a long time. The regular type of communication with other neurons is continued (see Figure 7. 1 herein).
Noticeable changes in the neuron functional state began to appear after about 10-14 hours of the LTA mode functioning.
Stage 2. Slight pain. (Consciousness is sometimes distracted by the appearing discomfort.) At this stage
12 | Cardiometry | Issue 17. November 2020
of the persistent LTA, at first rarely, and later on more often, observed is a change in the neuron background activity associated with the development of a sensitization response in the area of LTA application and the secondary hyperalgesia, leading to the central sensitization at the subsequent stages. These changes are elec-trophysiologically manifested by an increase in the AP generation rate by that neuron, which has an increased excitatory synaptic supply. In the second half of stage 2, the synaptically conditioned, pacemaker rhythmic activity begins to prevail (see Figure 7.2 herein). The spontaneous activity of the neuron with its adequate activation of EPSP and rest pauses gradually goes to a rhythmic pacemaker mode, which smoothly becomes dominant in the activity performance of the neuron.
Stage 3. Dull persistent pain. (Consciousness is more often distracted by the disturbing factor).
Weak pain is experienced. An increased rate of synaptic signal groups and APs initiated by them are found. The membrane potential is within the limits of the background level. The supply of the secondary hyperalgesia EPSP pulses to the neuron increases the AP rhythm. At this stage, the continuous spiking is periodically interrupted by the neuron, which creates waves of endogenous hyperpolarization of the neuron membrane. The initial AP grouping to short bursts under the influence of an increasing supply by EPSP is observed. The wave of the membrane exogenous depolarization appearing due to EPSP is insignificant, and it is easily compensated by the neuron with the subsequent conventional endogenous mechanisms of the membrane hyperpolarization. This endogenous hyperpolarizing wave (EHW) completely locks the AP generation, and, after the action by EHW at this stage, the AP firing rate of the neuron decreases, giving the neuron a relaxation. This is the most important factor in the neuron's survival program or "the Smile of the Life" (refer to Figure 7. 3 herein)
Stage 4. Precoma. Intense pain appears and disappears. Throbbing pain. (State of somnolence, a slight disturbance of consciousness).
1-1.5 hours after stage 3, the rhythmic generation of AP is interrupted by pulses of small EHP membrane amplitude, producing a pause in the AP firing. EHP at this stage keeps the membrane potential of the neuron within the functional norm. Pulsations of the neuron functional state take place (refer to Figure 7. 4).
Stage 5. Exhausting pain. (Coma scale degree 1. Severe pain. Constant increasing pain is experienced.
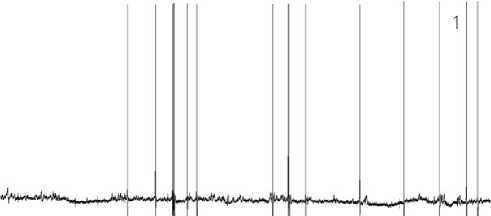
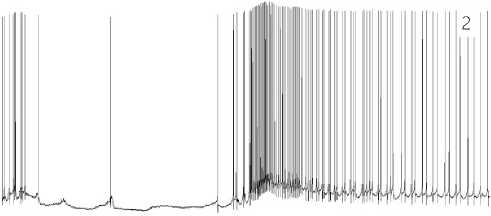
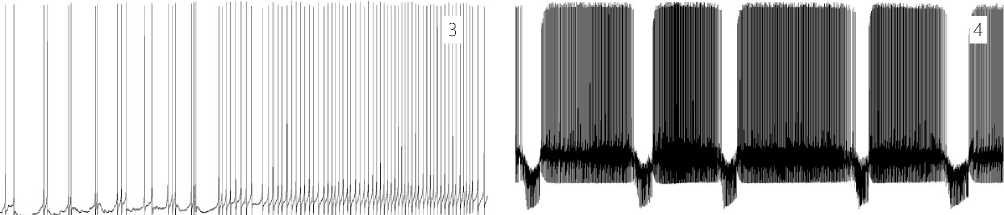


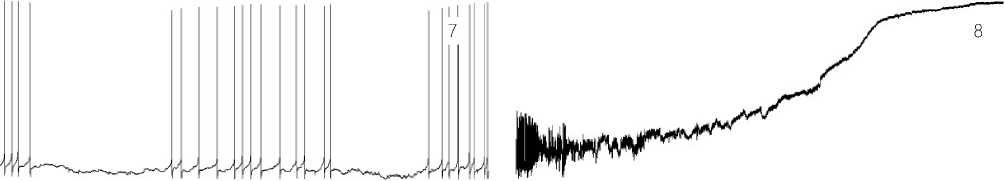
Figure 7. 1 – 8. Stages of the neuron functional state under the LTA conditions.
Confusion of consciousness, drowsiness. Central sensitization is developing).
The membrane potential of the neuron is depolarized due to a more intense synaptic influx. The high-amplitude Щ-type activity appears. In this case, the law of the membrane physiology is applicable according to which those HP waves, which have higher amplitudes, produce greater output effects, and the AP firing rate becomes higher upon the EHP completion (see Figure 7.5 herein).
It should be noted that we use definitions “the Щ-type activity” and “the anti-Щ-type activity” of the neurons, where letter Щ from the Cyrillic alphabet is applied as the most suitable symbol to illustrate the specific pattern of the neuron activity. This terminology is introduced herein in order to provide the most illustrative image of the repetitive pattern found in the response by neurons to LTA. So, the Щ-type activity is a type of the activity of the neuron, which produces a pattern shape in recording, similar to shaping style typical for Cyrillic letter Щ. The three vertical lines going upwards in the above Cyrillic letter denote a higher firing rate of action potentials. The descender of letter Щ, i.e. the portion of the letter that extends below the baseline of the font, is a symbol of the wave of the endogenous hyperpolarization of the membrane, which promotes the full termination of generation of action potentials. In other words, the Щ-type-activity represents first an exogenous excitation, realized by frequent action potentials, and by the end we can identify an inhibition produced by the EHP wave of the neuron, with promoting its transition into a more quite rest state. The anti-Щ-type activity is a Щ-activity antagonistic wave. This sort of waves consists of a wave of the endogenous depolarization (EDP) of the membrane, where the more frequent APs are found according to the respective level of depolarization. The anti-Щ-type activity excites the neuron. The anti-Щ-type activity becomes dominant at final stages after the point of no return, accelerating the neuron switching-off.
Pain is very strong or pulsating (Coma scale degree II. Neuropathic pain. Epileptiform activity. Obnubilation). Depolarization of the neuron membrane uninterruptedly increases. The EHP high-amplitude waves seek to return the level of MP to its normal value, but a growing excitatory exogenous stream due to irradiation of the processes of sensitization results in a steady increase in depolarization of MP (see Figure 7.6 herein). An initially moderate onset process of the EDP waves transforms to the anti-Щ-type activity. Figure 8 herein exhibits a sample of responses by the neuron to LTA, typical for stage 6 that represents the sequentially appearing endogenous waves, which depolarize the membrane of the neuron. The duration of the given recording segment in test No.84 is 3 hours 24 minutes.
Stage 7. Coma scale degree III. (No subjective perception of pain. Loss of consciousness. Strictly speaking, the consciousness is disordered.)
Depolarization of the membrane of the neuron is progressively increasing. The mode of operation of the neuron becomes extreme, close to the critical condition. Due to a multitude of the excitatory signals, in the neuron those programs are initiated, which make attempts to transfer it to its switch-off state. It is known that the neuron stops spiking, when its membrane is either completely hyperpolarized or depolarized. At this stage, one of the running programs generates powerful pulses depolarizing the neuron membrane. Depolarization of the membrane increases the firing rate of APs (see Figure 7.7 herein). The antagonist to this program is another program: the program of the neuron life – SL, which elaborates intensive endogenous mechanisms of hyperpolarization, which are trying to neutralize dangerous actions and effects produced by the excitatory signals.
Some results of our analysis of the above mentioned recording segment at stage 7 can be illustrated with graphics as given below. The red curve in Figure 9 herein indicates the dynamics in an increase in the firing rate of action potentials in the isolated classes. The blue curve illustrates the dynamics of the power of the HP waves in the same classes calculated by the formula as given below:
PHPW = k∑Si where PHPW is the power of the hyperpolarization wave; Si is the quantum area of the hyperpolarization wave; k is the quantization coefficient.
When analyzing the dynamics of the two reciprocal processes shown in Figure 9 herein, we can see an amplification of the power output of the inhibitory hyperpolarization waves after a sharp increase in the AP firing rate due to the excitatory signals, depolarizing the membrane, supplied to the neuron. The entire process occurs with a constantly growing depolarization of the membrane. Portion one in the recording segment in question indicates that there are powerful hyperpolarization processes, which successfully reduce the AP firing rate. But as to portion two thereof, we can observe that the excitatory influences on the neuron dominate, the next endogenous program of the extreme depolarization of the neuron membrane is initiated to neutralize the action and the effect of the hyperpolarization shifts of MP, and a vigorous increase in the AP firing rate begins.
JP^IMJIf^.Wf'W^^ ।
hW ■ ^ -^ 1 *^' '.^ Я О A Г- .. ^ ^ ^ = ^ *
* I я A ■ Я> * i*A M^A #Лм . Д i^^^*i
ibm #^ ^< ll^^i R^* ч^^8 МР®1< *-*wii и^ь ^ji^.
Figure 8. Endogenous depolarizing waves, the anti-Щ-type activity at stage 6: the response by neuron to LTA.
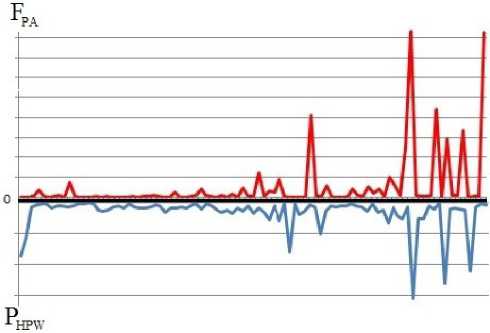
Figure 9. Dynamics of FPA of AP firing rate (the red curve) and power PHPW of the HP waves (the blue curve). Recording time is given along the ordinate axis. The recording segment duration is 10 minutes. Isolated class of 6 seconds.
Stage 8. Coma scale degree IV (terminal coma, no consciousness). Final stage. Neuron switching-off.
This stage is featured by two antagonistic processes. The first of them, namely the switching-off program is determined by the action of the exogenous factor, i.e. the supply of the excitatory influences exerted by the neuronal network on the neuron. The membrane bursting excitation initiates the depolarization anti-Щ-type waves. The second process, the SL program, is governed by the involvement of the Щ-type activity, i.e. by the endogenous factors, which hyperpolarize the membrane and which make attempts to return it to its normal state.
At this time observed is a critical phase, when the two competitive programs are running: the rivalry between the program of turning-off and that of the life. The membrane demonstrates the highest degree of its depolarization: specifically, the difference between the inside and the outside membrane potential becomes as small as possible. AP firing stops. The high-frequency EHP of low amplitudes gradually decays. No spikes are found. The winner in the competition is the program operating in synergy with the exogenous excitatory potentials. The generating mechanisms of the membrane are switched off. Intracellular electrical potentials of the neuron cannot be recorded. We observe markers of the end of the neuron biological life (see Figure 7.8 herein). The recorded line is matched with the isoelectrical baseline on the recording.
So, we have traced a stage-by-stage change in the functional state in the CNS neurons under the longterm action of the low-threshold pain factor and iden-
Table 1
It should be noted that the entire available period of our investigations of the neuron responsiveness in vivo has demonstrated that it is fully applicable to the performance of complex, open-end, oscillating, or to define it otherwise, self-organized systems that is a subject matter of synergetics [14, 15]. Adhering to this way, we have shown that weak fluctuations being prioritized lead to considerable adaptation re-settings in this sort of systems, i.e. these are precisely the weak fluctuations which govern their performance and periodical repetition of development of different types of the adaptational reaction archetypes (models) from the reaction of training up to and including stress at different levels of responsiveness from the cell level up to and including the organism level [16-18]. As to the LTA effects, the case in point is not an absolute value of the low-threshold pathogenic stimulus, but rather the durations of the action and capabilities of converting the action rates into a force, when deviations from the normal state are increasing, and, as a consequence, the mechanisms of the phase transition to another level of responsiveness come into operation. It has become possible for us to identify certain bifurcation points, when the system transforms to a new state, based on a more detailed analysis of the state of the neuron in the Smile-of-the-Life terms according to our definition as offered above (see Figure 2), where indicated are points of return 1 and 2 within the viability zone of a biological system.
F, Hz
1,6
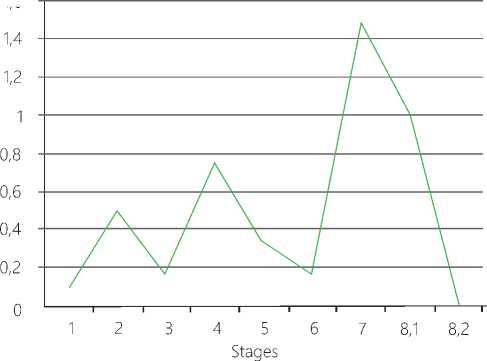
А. High-amplitude oscillation dynamics of firing rate activity.
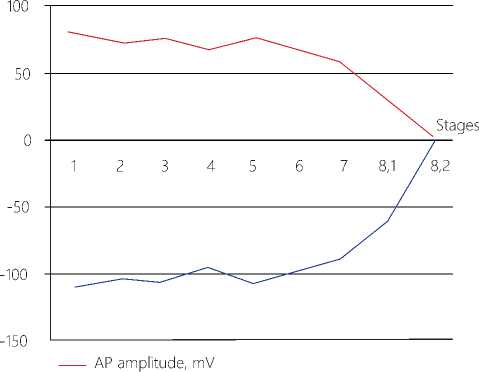
---Dynamic membrane potential, mV
-
B. Synergy between the dynamic MP and the AP amplitude
Figure 10. Generalized results upon processing of stage-by-stage changes in the functional state of the CNS neurons under long-term low-threshold pain factor conditions. The total recording time in 61 experiments with LTA is 1010 msec. Averaged values according to the t test with significance p ˂ 0,05 are given herein.
Point of return (Reversibility)
As is true of all points within the Smile-of-the-Life region, so with points 1 and 2 we indicate adaptive and most optimal functioning of all metabolic and nervous processes in an organism. The points of return (reversibility) stand for those transition points, where a dynamic natural process of the control and regulation of the organism performance passes a critical level in a given functional state in order to reach a new level of the normal vital functions.
A B C D
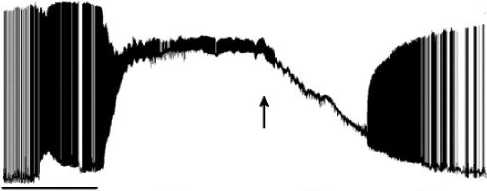
Figure 11. Biopotentials of the neuron before and after passing the point of return. The point of return (reversibility) is indicated with an arrow. Figure 13 exhibits magnified fragments A, B, C and D thereof.
JBI D A
.J^^MUl-ll^^
C
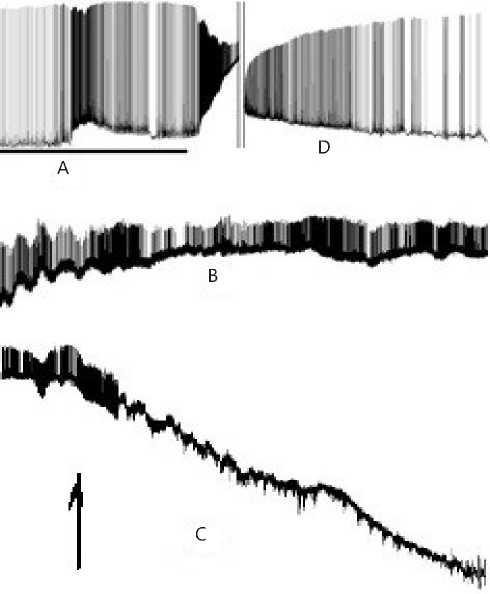
Figure 12. Magnified fragments A, B, C and D taken from Figure 11. Legend: A – activity of the neuron before the point of return. The solid line below shows the LTA continuation and the end. Next the process of return is in progress. B – Coming up closer to the point of return. C – Activity when passing the point of return and immediately upon passing thereof. Restorative hyperpolarization of membrane potential; the point of return (reversibility) is indicated with an arrow. D – Restoration of the normal activity of the neuron after the point of return.
We should clarify that in Figure 12.A the solid line below is an indication of the continuation and end of the LTA mode. An enhancing response as an excitation wave at the output is a sign of the restoration of the normal performance of the neuron axon after the sensitization induced by LTA.
Particular attention should be given to scarcely illustrative Figures 12 B and C herein as it might seem
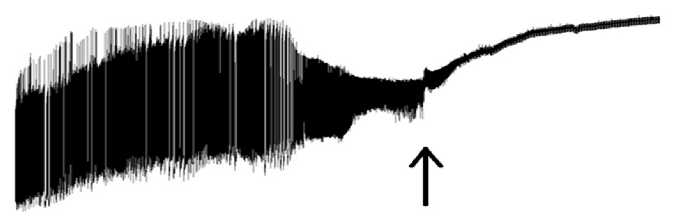
Figure 13. General view: final stage.

Figure 14. The termination.
first to our thoughtful reader. In our opinion, these are precisely the pictures, which are the best illustrations of the Great Mystery of the Life. These figure parts show strong differences from parts A and D in Figure 12, where we can clearly trace the life span of the given neuron, demonstrating its external interrelations with the network neurons by the AP generation. Parts B and C in Figure 12 seem to be rather scarcely expressed, that is especially true for part 13.C. However suddenly, as exhibited in part D in Figure 12, appearing is the life, APs are being generated by the seemingly silent neuron, which, nevertheless, has just passed its bifurcation point and transited to a new phase state.
Point of no return (Irreversibility)
A close look at the processes, which characterize the activity of the neuron, when it is approaching the point of no return, shows that the dynamics converts to an evolutionally more archaic rhythm. In the circumstances, when we deal with the continuing LTA, the membrane is highly depolarized up to the critical values. The sinusoidal activity pattern becomes more pronounced; it is waveform-amplitude-modulated. With time the amplitude of the waves decreases, the oscillation frequency becomes higher and instable. Some trains composed by 2-3 low-amplitude waves separated by a single high-amplitude wave can be found.
Our recording (see Figure 13 herein) has captured a fragment demonstrating a struggle between the neuron switching-off and its return to the SL region. This may be compared to the human unconsciousness state, when the nervous system is operating according to the internal reflex regulation. Figure 14 displays points of no return (irreversibility), when and where the life of the neuron is ended with “a guitar string breaking sound”, i.e. high-frequency decaying oscillations similar to that when a guitar string breaks, and the typical sound can be heard, but no further music is possible.
The bioelectrical activity of the neuron ceases. Some dying down HP shifts are recorded (indicated with arrows). The mechanism implies that, upon passing the point of no return, the HP high-amplitude waves are turned off, and the membrane of the neuron is further highly polarized. The neuron changes to the mode of generation of high-frequency waves having approximately the same amplitude. Upon expiration of certain time, some regulation features can be traced at a new level of excitation as amplitude modulation of the high-frequency waves with the EDP waves, generated by the neuron. This regulation is speeding up the switching-off of the neuron. The amplitude modulation is performed with depolarization waves, which are no more capable of increasing the limit frequency of the waves, but the effect produced by the endogenous DP waves decrease the amplitude of the waves, generated by the neuron. Further observed is a more frequent effect of the endogenous DP waves, the wave amplitude becomes lower, and the duration of the modulated wave falls. However against the background of a growing effect made by the DP waves, the endogenous mechanism responsible for their generation is also not capable of providing its proper operation: it fails. Let us discuss Figure 13 again: it is evident that some restoration of the amplitude of the high-frequency waves is available, and a lowered frequency in the action of the endogenous depolarization waves can be detected. But it is still a temporary effect, and again and again we observe the actions and effects due to constantly increasing depolarization influence: a reduction in amplitude of the wave and a rise in the number of the desynchronized waves. This pattern shows that generation of bio-potentials is managed according to an evolutionally ancient firing type. This sort of rhythms has been detected by us in recording of the bio-electrical activity in colony of fungi Fuzari-um oxisporus (unpublished data) as well as in protozoa according to our published data [19]. The depolarization of the neuron membrane becomes close to its limit value. There comes a time, when the biological activity of the neuron cannot be detected: the neuron is switched off.
However in our experiments some individual command neurons in CNS have demonstrated a somewhat different way of passing the point of no return (irre-
Issue 17. November 2020 | Cardiometry | 17
versibility) (please, see Figure 14 herein). The above mentioned Figure exhibits the operation of the mechanisms, which are responsible for the neuron control and regulation and which are targeted at the return of the performance of the neuron to its normal state, i.e. to provide the proper execution of life program SL. This implies generation by the neuron of the endogenous hyperpolarization pulses as mentioned above. However since the damaging factor remains still in action, a more dramatic activation of the neuron takes place due to a powerful stimulating synaptic supply, the membrane of the neuron is rapidly depolarized, and the Щ-type activity is no longer effective: it is suppressed. Appeared are the no-return-point conditions: although AP firing is supported, the AP amplitudes under growing depolarization of the membrane become lower and lower, so that they finally drop to zero.
This moment is marked in Figure 15 with an arrow: refer to the wording “Action Potentials decay”. This is the start point for initiation and execution of the neuron switching-off program. Pulses of extra-depolarization of the neuron switching-off program can be found in the above Figure as high-amplitude spikes having a steep leading edge for the phase of strain and a flattened trailing edge for the relaxation. These pulses exhibit their high rhythmicity as in the case of epileptic seizure with the firing rate fall and the firing cease. It’s the final agony, the last surge of energy that supports and speeds up the membrane depolarization process. Upon the final energy surging, a chaotic process of firing and decreasing amplitude is detected; next the recording line becomes flat. The neuron with its own infrastructure attains equilibrium; the level of entropy reaches its maximum, and the membrane changes to its turned-off state..
Conclusion
The factors of the internal environment in an organism demonstrate always persistent dynamics by interacting with the external environment in order to control stability of some parameters and at the same time maintain instability of the other. It is because of the state of the permanent imbalance that always features a living system as opposed to inanimate nature [20]. The state of thermodynamic equilibrium, or the state of maximum entropy, is equivalent to the death for a living system. This state is attained by the neuron upon passing the point of no return. In case of passing the no-return-point, the question arises why the 18 | Cardiometry | Issue 17. November 2020
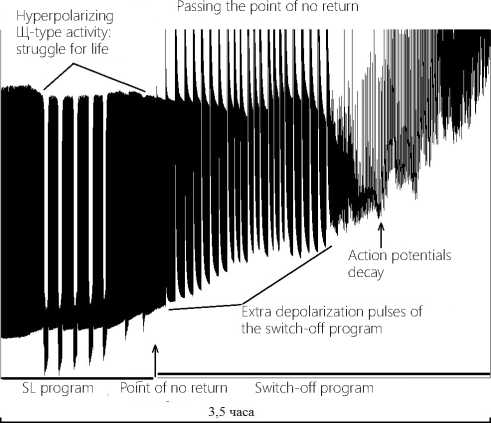
Figure 15. Dynamics of passing the point of no return by the command neuron in CNS.
According to our experimental evidence, the critical point may be achieved even in case when the neuron performance is in the Smile-of-the-Life region, when a low-intensity, persistently damaging, harmful factor acts. In this case, the persistence should be treated not only as a prolonged existence of an individual damaging factor, but also as the recurrence of some risk factor actions that may induce conversion of the recurrence rate into a power. By analogy, we assume that the above interpretation of the recurring risk factors may be applied to a human social behavior and comprise disputes in a family, job-related conflicts or some other unsolvable problems, i.e. recurring actions of risk factors with varying modality, which should be considered as stress situations affecting the internal environment in a human organism. In the circumstances, within a prolonged time, observed is an enhancement of the controller oscillations with permanently growing amplitudes, so that at a certain moment the bio-system is coming closely to the noreturn-point.
In such a manner our concept that under the longterm supply of excitatory pulses, causing the limit depolarization of the membrane, the program of the neuron death is activated and executed, has been supplemented and specified with evidence for switching on the program to protect the neuron from its death: the neuron initiates powerful firing to depolarize the membrane (see Figure 15 herein), so that vast consumption of energy used for excessive input excitation is prohibited. These pulses, or to simplify, the change of the membrane for its extra depolarization with the internal molecular membrane keys are emergency protection means of the proper neuron. The neuron in the circumstances is not capable of responding to input excitation. The neuron is turned off.
We cannot state that all neurons operate strictly in accordance with the same routine procedure. Each neuron has its own individual peculiarities and demonstrates its own specific features, especially in responding patterns. Our challenge has been to analyze a great variety of the responses by neurons, recorded in our experiments in a lot of the neurons, identify most general laws, typical for all neurons, which govern the regulation by the neurons of the functional state under the action of a damaging factor accompanied by cumulative pain, growing with time.
It may seem that the mechanisms of the regulation by the neuron of its own functional state are limited. We have succeeded to identify that the most effective regulator of the functional state employed by the neuron is the periodical repeated pattern excitation – inhibition, i.e. stage-by-stage change of the potential level of the cytoplasmatic membrane of the neuron by the EHP waves and the pulses of the hyperpolarization of the membrane. As a conclusion, we have discovered that the main neuron response tendency for the mode of chronization of LTA applied in our experimental trials is the transitions of the neuron to different levels of the excitation state (graded rising of the excitation state from one level to another).
It is of importance that it is precisely these transitions, detected at the neuron cell regulation level, which first are the key point in general biological regularity of periodical repetition of development of the archetypes of the adaptation responses at different levels of reactivity in a human organism. Years of research conducted by Scientific School headed by L.Kh. Garkavi have shown that the existence of different archetypes of general nonspecific responses at every level (period) provides for adequate responding attaining the points of return to the normal performance. This biologically reasonable, periodical, multi-level system, which reflects the principles of fractality, is designed to maintain the life both of an organism and a cell within the Smile-of-the-Life region. In our opinion, Smile of the Life is the general law applicable to all living systems.
Second the obtained evidence for the existence of the return and no-return points in the context of the performance of the neuron under the conditions of the LTA chronization, analyzed from the standpoint of synergetics, has demonstrated the phase transitions to a new discrete state, which must fully comply with
Issue 17. November 2020 | Cardiometry | 19
the available logistics and the laws of maintenance of the life in complex open-end nonlinear systems. Therefore it is critical to take into account for this sort of self-organizations that even a minor factor, which has a long-term action, may involve resetting of all interconnected processes in the intracellular homeostasis governed by the command centers of the integrated system as a whole.
It can be illustrated by a great variety of exemplary cases pertaining to “strong actions of low-intensity stimulators” that has been discussed more than once in the framework of many international symposia and publications [25, 26]. Scientists at the Oncology Research Center in Rostov-on-Don (Russia) have been investigating and conducting experimental trials referred to their topical issue Chronic Pain as Tumor Growth Modulator [27-31]. They have arrived at a conclusion that it is just pain chro-nization produced by long-term compressive irritations of the neurons, which trigger complications of diseases in cardiology and oncology, when a tumor causes pressure and compression of the nerves, or produces disorders or even turns off the required nervous regulation. As to cardiology or oncology patients, we can often observe that they initially experience pain attacks or pain episodes, and next the pain episodes either become growing with a certain periodicity, or pain achieves longevity as it has been demonstrated by continuous recordings in our uninterrupted experiments in the neurons in question studied by us. The obtained evidence opens up prospects for the proper understanding of the electrophysiological component of the mechanism of chronic neurogenic pain and may be of interest to predict efficacy of pain relieving therapy in the high-risk long-term pathology low-threshold actions and effects.
Statement on ethical issues
Research involving people and/or animals is in full compliance with current national and international ethical standards.
Conflict of interest
None declared.
Author contributions
The authors read the ICMJE criteria for authorship and approved the final manuscript.
20 | Cardiometry | Issue 17. November 2020
References Intracellular electrophysiological evidence: how pain is experienced by neurons
- Shnol SE. Physical and chemical factors of biologi¬cal evolution. 1979. Moscow:Nauka. [in Russian]
- Aleksandrov YI, et al. Neuron. processing of signals. Plasticity. Modeling: A fundamental guide. Tyumen: Tyumen state University press, 2008. 548 p.
- Sakharov DA. Genealogy of neurons, Moscow, Nauka, 1974. [in Russian]
- Samygin FI. Neural mechanisms of the efferent link of the defensive reflex of the grape snail body retrac¬tion into the shell. Dissertation of the candidate of bi¬ological Sciences: 03.00.13. Rostov-on-don, 1981. 143 p. [in Russian]
- Burlakova EB. effect of ultra-low doses. Bulletin of the Russian Academy of Sciences. 1994;64(5):425-31. [in Russian]
- Spitkovsky DM. The concept of the effect of low doses of ionizing radiation on cells, and its possible use for the interpretation of the biomedical conse¬quences of the Chernobyl accident. Radiobiology. 1992;32(3):382-400. [in Russian]
- Orlov VI, et al. Cardiometry. 2018;12:39-53.
- Shikhlyarova AI. The role of biotropic parameters of electromagnetic fields in increasing non-specific antitumor resistance. Diss.Doct. Biol. Sciences. Ros¬tov-on-Don. 2001. 50 p. [in Russian]
- Shikhlyarova AI. On the possibility of overdosage with the medical applications of weak magnetic fields. Influence of magnetic fields on biological objects. Ka¬liningrad. 1975. [in Russian]
- Lapicque L. L'excitabilité en fonction du temps. La chronaxie, sa signification et sa mesure. 1926.
- Orlov VI, Laskov VN, Karpenko LD. Method of preparation of the Central nervous system in the grape snail. Author's certificate SU 1561962 A1, May, 7, 1990. Application no. 4455188 dated July 5,1988. [in Russian]
- Orlov VI, et al. Cardiometry. 2017;11:17-27.
- Sukhov AG, et al. Cholinergic and potential-de¬pendent mechanisms of local rhythmogenesis in neu¬ral columns of the rat somatic cortex Rostov-on-Don: SFedU publishing house, 2011. 346 p. [in Russian]
- Haken G.,1985. Synergetics. Moscow:Mir.1985 . 450p. [in Russian]
- Prigozhin I, Stengers I. Order out of chaos.Mos¬cow: Progress, 1986. [in Russian] 432 p.
- Garkavi LKh, Kvakina EB, Kuzmenko TS, shikhl¬yarova AI. Antistress reactions and activation therapy. Yekaterinburg: Philanthropist. V.1. 2002. 360p.; V. 2. 2003. 275p. [in Russian]
- Kit OI, et al. Cardiometry.2015.7:11-7.
- Shikhlyarova AI, et al. Influence of low-frequency low-intensity magnetic field on the functional state of the central nervous system and the dynamics of general non-specific adaptation reactions in patients with lung can¬cer. News of Higher Educational institutions. North Cauca¬sus region. Natural science. 2005;10(34):93-8. [in Russian]
- Grechenko TN, et al. Psychophysiological analysis of oscillatory processes in the behavior of biosocial systems. Psychological journal. 2015;36(6):76-87. [in Russian]
- Schrodinger E. What is life? The physical aspect of a living cell. Moscow, Izhevsk, 2002. 93 p. [in Russian]
- Shvyrkov VB. Neurophys. study of systemic mechanisms of behavior. Moscow: Nauka, 1978. 240 p. [in Russian]
- Draguhn A, Traub RD, Schmitz D, Jefferys JGR. Elec¬trical coupling underlies high-frequency oscillations in the hippocampus in vitro. Nature. 1998;394:189-92.
- Brivanlou AH. Should the Master Regulator Rest in Peace? Nature genetics. 1998;20(2):109-10. doi: 10.1038/2402
- Loewenstein WR. The touchstone of life. Molecu¬lar information, Cell communication, and the founda-tions of life: N. Y., 1999. P. 5-611.
- Petkov B. Medicine, organism, pharmacological effect. Sofia.1974. [in Russian]
- Podkolzin AA, Dontsov VI. Low-intensity factors in bioactivation and immunocorrection.Moscow: Panas-Aero. 1995.195 p. [in Russian]
- Kit OI, et al. Some mechanisms of increasing the malignancy of melanoma. Russian journal of pain. 2017;(2):14-20. [in Russian]
- Kit OI, et al. The way of the abolition of the ge¬netically determined growth inhibition of malig¬nant tumors in the experiment. Patent for invention No. 2718671 Russian Federation, application No. 2019124739/14 (048267) dated August, 01, 2019. Publ. 13.04.2020. Bul. No. 11
- Frantsiyants YE, et al. Method for stimulating chronic pain of malignant growth in the lungs of rats. BEBiM. 2020;169(2):257-9. [in Russian]
- Zhukova GV, et al. Effect of chronic pain of various etiologies on the tumor process in the experiment. Mod¬ern problems of science and education.2020;2:143-51. DOI: 10.17513/spno.29699. [in Russian]
- Zhukova GV, et al. Some Approaches to the Ac¬tivation of Antitumor Resistance Mechanisms and Functional Analogs in the Categories of Synerget¬ics. Biophysics. 2016;61(2):303-15. DOI: 10.1134/S0006350916020251 [in Russian]
