Использование математических методов при изучении взаимоотношений подсолнечника с патогенными организмами

Автор: Дьяков А.Б., Васильева Т.А., Головин А.В., Войко Ю.Г.
Статья в выпуске: 2 (131), 2004 года.
Бесплатный доступ
На основе анализа литературных данных и результатов собственных исследований авторов показано, что количественные подходы с использованием математических методов позволяют повышать эффективность исследований взаимоотношений возделываемых растений с патогенными организмами. На ряде конкретных примеров показано, что математическое моделирование позволяет' получать результаты, которые не могут быть получены на основе использования качественных или полуколичественных методов исследования. Продемонстрированы возможности математическою описания развития процессов поражения растений патогенами, прогноза эпифитотий, зависимостей потерь урожая от степени поражения посевов в разных агроэкологических условиях, что необходимо для вычисления порогов вредоносности в различных экономических ситуациях. Кроме того, показана возможность и степень эффективности селекции на полигенно контролируемую устойчивость к патогену, предлагается способ оценки толерантности генотипов к поражению болезнью.
Короткий адрес: https://sciup.org/142150688
IDR: 142150688 | УДК: 633.854.78:622.0[517
Using of mathematics methods in studying sunflower reactions on pathogenic organisms
It is shown on the base of literary data and results of author's researches that quantitative approaches with usage of mathematics methods allow' increasing the efficiency of researches of cultivated plant reactions on pathogenic organisms. It is shown on the number of concrete examples that mathematics simulation allows to get results, which can’t be obtained on the base of qualitative and semi-quantitative research methods. Opportunities of mathematics description of development of plant affection process by pathogens, forecast of epiphitoties, dependence of yield losses of sowing affection extent in different agroecological conditions are demonstrated. It is necessary for calculation of harmfulness threshold in various economic situations. Besides that opportunity and extent of influence of breeding on polygene-controlled resistance to pathogen, method of estimation genotype tolerance to disease is proposed.
Текст научной статьи Использование математических методов при изучении взаимоотношений подсолнечника с патогенными организмами
Объекгом исследований специалистов по защи те растений являются не агроценозы. изучаемые агрономами. а более сложные системы - биоценозы, включающие в себя сообщества не только возделываемых растений, но и комплексов других организмов. Использование пестицидов, приемов агротехники, устойчивых сортов и других способов воздействия па фитосанитарное состояние полей должно интегрироваться таким образом, чтобы, но формулировке Д.А.Робертса (1981), "... держать численность популяций вредителей растений на уровне экономического порога вредоносности или на еще более низком'1 (с. 241-242). При этом он включает в понятие "вредители растений" все вызывающие потери урожая организмы, в лом числе возбудителей болезней, насекомых и других членистоногих. грызунов, птиц, сорных растений (Робертс, 1981, с. 15-16). Применение методов биологической борьбы еще более усложняет задачи исследования взаимосвязей в биоценозах, г. к. при злом надо понимать еще и детали взаимоотношений вредоносных видов с их естественными врагами и разрабатывать на этой основе способы воздействия на численность этих популяций.
Закономерное л и изменения численности популяций и их взаимоотношений не голько в природных, ио и в созданных человеком сообществах (поля, сады, зернохранилища и г. и.) изучает экология, причем решающую роль в лаких исследованиях играют маземагические меюды (Бигон и др.. 1989. с. 11-12). В частности, прогнозы, основанные на понимании таких закономерностей, позволяют своевременно принимать надлежащие меры для сбережения урожаев. Дж. Э. Ван дер Планк (1977) подчеркивает необходимость правильно определять время применения пестицидов: если химикаты применяются слишком рано, они могул оказалься неэффективными, если слишком потно - ущерб можеі быль уже непоправим. Поэтому он считает важным знание кривой развития конкретной боле ши с уче том специфичных для нее де талей, а также умение аккуратно количественно переводить проінозы лаких кривых на размеры потерь урожая, чтобы обеспечивал а эффективность принимаемых мер. Сам Дж. Э. Ван дер Планк впервые применил математические методы изучения эпифитотий разных типов. Это позволило создать стройную теорию эпифиютиологии, ставшую поворотным пунктом в развитии патологии растений, оцененную как "выдающееся интеллектуальное достижение" (Тарр, 1975. с. 353).
При создании эффективных интегральных систем методов борьбы с вредными организмами особенно важно выявлять и изучать факторы, влияющие на размножение и гибель особей, гак как, воздействуя на эли факторы, можно регулировать численность популяций. Надежно выявлять такие факторы и количественно оценивать эффекты их действия и взаимодействия на динамику популяций, различающихся по типу репродукции, можно только на основе использования имитационных математических моделей (Смит, 1970: Waggoner, 1974; Франс, Торили, 1987, с. 221-223), Для практического прогнозирования возникновения и скорости развития болезней используют также эмпирические формулы (Франс, Торили, 1987, с. 220-221).
Учитывая важность и разнообразие задач, при решении которых необходимо использование математических методов, Дж. Э. Ван дер Планк (1977) считает неправильным сложившееся положение, при котором в штатах отделов фитопатологии и энтомологии много химиков, ио мало математиков. При этом он подчеркивает, что имеет в виду математику, отличную от той, которая связана с планированием опытов и проверкой значимости результатов. Однако в отечественном руководстве по математическим методам в фитопатологии (Минкевич, Захарова, 1977) изложены только статистические методы изучения вариабельности признаков, оценки существенности различий между средними, вычисления коэффициентов корреляции, но даже нет упоминания о регрессионном анализе. хотя в аннотации заявлено о методах оценки вредоносности болезней.
Среди комплекса факторов, влияющих на численность причиняющих вред организмов, важнейшими являются: количество первоначального инокулюма, скорость увеличения численности популяции особей, поражающих возделываемые растения, ограниченная емкость среды для развития вредоносных организмов, лимитирующая рост популяции патогена (Ван дер Планк, 1977; Рббертс, 1981, с. 68-69, 75-76). Влияние этих факторов на характер эпифитотий и описывающих их математических моделей существенно различно в зависимости от соотношения длительностей циклов развития паразита и хозяина. В частности, по продолжительности онтогенезов до созревания семян подсолнечник и шразиха мало различаются, поэтому первостепенное значение приобретает количество инокулюма. 1л о влияние на характер поражения корневых систем растений восприимчивой популяции подсолнечника оценивали количественно по данным вегетационного опыта (табл.). При этом обнаружено, что характер зависимости пораженности корней подсолнечника заразихой от количества инокулюма различен при разных сроках учета.
Таблица. Пораженность корней подсолнечника заразихой в зависимости от срока Учета и количества инокулюма
|
Семян шразихи 1 на сосуд (х) |
Учет на 25-й день по клубенькам |
Учет- на 70-й день по цветоносам |
||||
|
нораже-но растений, % |
клубеньков шт./раст. (у) |
поражено растений, % |
цветоносов шт./раст. (z) |
|||
|
найдено |
вычислено у -43х |
найдено |
вычислено z-63x/(0.041 х) |
|||
|
0,00 |
0 |
0 |
0 |
0 |
0 |
0 |
|
0.01 |
50 |
0,7 |
0.4 |
100 |
13.6 |
12,6 |
|
0,03 |
75 |
1.6 |
1,3 |
100 |
28.0 |
27,0 |
|
0.09 |
100 |
4,1 |
3,9 |
100 |
42,5 |
43.6 |
|
0.27 |
100 |
10.9 |
1 1,6 |
100 |
52.2 |
54.8 |
|
0.81 |
100 |
35.6 |
34,8 |
100 |
61,7 |
59.3 |
Первый учет был проведен в период интенсивного заселения корней подсолнечника паки сном. В это время число особей паразита на корневых системах еще не ограничивалось емкостью среды обитания, т. е. ресурсами растения-хозяина. Поэтому при учете в первый срок численность популяции особей заразихи, поразивших подсолнечник, зависела в основном от биотического потенциала патогена, который в данном случае определяется количеством семян паразита в почве. Это определяет характер зависимости числа клубеньков заразихи (у) в среднем на одну корневую систему подсолнечника от оличества семян паразита на сосуд (х). Но данным учета на 25-й день, эта зависимость в пученной области описывается уравнением прямой линии, проходящей через начало аорт шаг:
У bx. 111
іс b - коэффициент регрессии. В условиях проведенного опыта b :43. Следовательно, в жальный период, когда процесс заражения еще полностью не завершился, количество ■павших нарази тировать заразих зависело только от вероятности встречи хозяина с на-исном. К концу вегетации этот (ракгор имеет уже меньшее значение вследствие того, о чем больше число наразитирующих особей заразихи, гем в большей степени истощатся ресурсы рас гения-хозяина. Но згой причине соответственно усиливается лимити-аощее влияние ограниченной емкости среды обитания на увеличение числа особей фазихи. достигающих (разы образования цветоносов. Поэтому зависимость между коли-ттпюм семян паразита в почве (х) и числом учтенных на 70-й день цветоносов заразихи установится криволинейной и описывается уравнением гиперболы:
Z тах*х
2[2]
О + X де а - константа, /шах максимальное число растений заразихи, развитие которых до чраювания цветоносов могут обеспечить ресурсы корневой системы одного растения мдеолнечника. Очевидно, чем больше размеры растения-хозяина, тем большее предель-юе число особей паразита может развит ься на его корневой системе. В условиях описываемого опыта в среднем /шах 63. Многочисленные наблюдения в этом и других штах свидетельствуют также о том, что в равных условиях внешней среды при больном числе заразих на корневой системе подсолнечника уменьшается средний размер добей паразита. Это означает что между отдельными растениями заразихи на одной орневой системе наблюдается конкуренция за ограниченные ресурсы питания, которыми располагает растение-хозяин.
Вычисленные но уравнениям [1] и [2] числа клубеньков и цветоносов хорошо анласхюгся с экспериментальными данными (см. таблицу). Полученные результаты жтетельствмот о 'том, ч то при инфекционной нагрузке 0.81 г на сосуд семян заразихи фактически достигается предельное число особей паразита, которые способны развиться а корнях одною растения подсолнечника. Поэтому вывод, к которому пришли В.13. Кихтенко с соавторами (1987), что при оценке на заразихоустойчивость оптимальной на-:р\зкой является 1-2 і на сосуд семян заразихи, мог быть получен лишь при низкой чгшссиособност и семян паразита. Судя но данным таблицы, для отбраковки восприим-ивых растений подсолнечника при учетах по цветоносам достаточно вносить 0.03-0,08 г і;і сосуд семян шра шхи.
Наряду с моделированием эпифитотий использование математических методов сзусловно необходимо в исследованиях по определению экономических порогов вредо-осносіи организмов, наносящих ущерб урожаю возделываемых растений. Это очень ажная задача, однако до сих пор исследования в этой области ведутся в крайне малом бьсме. Но мнению Д.А. Робертса (1981, с. 78), такие исследования крайне необходимы, ак как мы часто нс знаем не только, когда популяция снижающих урожай организмов ревзойдеі допустимый уровень, но и что означает этот допустимый уровень. Подчерки-ж что меры защиты растений должны применяться лишь в ситуациях, когда они эффек-.тшны и экономически оправданы, Дж. Франс и Дж. X. М. Торили (1987, с. 218) пишут: Оценка потерь урожая от болезней и вредителей..... необходимый элемент" любой схемы управления, в основе которой лежат экономические критерии".
Экономический порог вредоносности - это такая численность популяции наносящего вред организма, которая причиняет ущерб, равный стоимости мероприятий по прсдоівращению этого ущерба (Хедли, 1977; Робергс, 1981, с. 77). Поскольку в условиях рыночной экономики как цены на защищаемую продукцию, так и стоимость мероприятий по защите растений могут значительно меняться, соответственно должны изменяться и экономически допустимые уровни численности популяции причиняющего ущерб организма. Поэтому практически пользоваться оценками экономических порогов вредоносности можно лишь в том случае, если они представляют собой не постоянные показатели уровня популяции организма, наносящего ущерб урожаю, а формулу зависимости величины такою ущерба от сзепени поражения растений с учетом с тоимости продукции и мер ее защиты (Хедли, 1977). При решении таких задач сравнительно легко определяются цены продукции и стоимости затрат; значительно труднее получить формулы для вычисления величин потерь урожая в зависимости от степени поражения растений (Keiber, 1977; Франс. Торили, 1987, с. 218). Обусловлено эго не только сложностью оценки плотности популяции наносящего вред вида. Еще более трудно количественно определить величину потерь урожая (Робергс, 1981, с. 13; Расселл, 1982, с. 12). В связи с этим, например, в учебнике В.Ф. Пересыпкина (1989), в котором описано более 400 болезней возделываемых растений, ни для одного из возбудителей не указан экономический порог вредоносности, а описания степени вредоносности во всех случаях непригодны для принятия решений об экономической целесообразности принятия в конкретных ситуациях мер по нпцитс рас гений.
В абсолютном большинстве публикаций по защите подсолнечника от вредителей и болезней также отсутствую!' сведения об экономических порогах вредоносности. Сведения о критических уровнях численности популяций можно найти в описаниях мер защиты растений от вредителей, в том числе наносящих ущерб подсолнечнику (Вронских, 1988; Тихонов, Пивень, 1991). Однако обычно в таких публикациях для каждого насекомого указан постоянный критический уровень численности, не увязанный ни со степенью наносимого ущерба, ни с экономической конъюнктурой. Более того, критические уровни, указанные в разных публикациях, различаются в 2-10 раз. Возможность нахождения зависимости снижения урожайности подсолнечника от численности проволочников продемонстрировала Р.В. Бартенева (1978). Это позволяет вычислять экономические пороги вредоносности при разных соотношениях цен продукции и стоимости мер борьбы.
В описаниях мер борьбы с болезнями подсолнечника оценки экономического порога вредоносное!и встречаются очень редко. Например, М.Д. Вронских (1988) считает экономическим порогом вредоносности корзиночной формы белой гнили поражение в фазу физиолог ической спелости 15-17 % растений. Этот порог он считает критерием необходимости проведения химической десикации посевов. Однако обоснования величины этою порога. в том числе зависимости величины потерь урожая от доли пораженных расзений, автор не приводит. Для выявления таких зависимостей используют разные уравнения регрессии (Франс, Торили, 1987, с. 219-220). Предложены формулы для вычисления потерь урожая разных культур в зависимости от степени их поражения патогенами: сахарной свеклы - церкоспорозом (Keiber, 1977), пшеницы - септориозом и фузариозом колосьев (Hartmut, 1982), конских бобов - заразихой (Mesa-Garcia, Garcia-Torres, 1984; Zaitoun el al., 1991) и другие. Такие уравнения необходимы не 'только для оптимизации мер защиты растений, но и для оценки степени толерантности генотипов к поражению болезнями при селекции растений на горизонтальную устойчивость.
Хотя гибриды и сорта подсолнечника селекции ВНИИМК устойчивы к распространенной в стране популяции заразихи, проблема оценки ее вредоносности остается актуальной не только при возделывании восприимчивых иностранных гибридов, но и в связи с возникновением в Испании новой вирулентной расы заразихи, поражающей все сорта и гибриды подсолнечника (Castejon, Garcia. 1977). Однако до сих пор не только не была установлена количественная зависимость уровня потерь урожая от степени поражения подсолнечника заразихой, но даже самые общие оценки оказываются иротиворечи- выми. Например, В.Д. Вронских (1988) пишет, что поражение подсолнечника заразихой снижает урожай семян на 50-70 %, но в гой же статье приводит вдвое более низкие экспериментальные оценки разницы в урожаях пораженных и непораженных растений. А.В. Фисюнов (1977) приводит данные об урожаях подсолнечника в одних и тех же условиях 12.1 ц/га при пораженности заразихой 25,5 % и 0J ц/га при пораженности 87 %. Использованием элеменгарной математики можно показать несуразность второй оценки: если 13 % непораженных растений дали урожай 0,1 ц/га, то при отсутствии поражения 100 % таких рас гений должны были бы обеспечить урожай 0,77 ц/га, т. е. в 15,7 раза ниже урожая, полученного при пораженности 25,5 %.
При разработке формул зависимости потерь урожая от уровня поражения растений важен правильный выбор критериев оценок как потерь, так и степени поражения для конкретных сочетаний хозяин-патоген (Haptniut, 1982). Например, при вычислениях по одним и гем же экспериментальным данным коэффициенты линейных корреляций между пропен гом пораженных заразихой расгений конских бобов и снижением их урожаев составили от г 0,130 до г 0,788 при оценках потерь урожая в тоннах и от г-:0,976 до г 0,996 при выражении потерь в процентах (Zaitoun, el al., 1991). При этом включение в уравнение еще и показателя среднего числа заразих на одно пораженное растение не повысило точность оценок. Y. Mesa-Garcia и L Garcia-Torres (1984) установили, что зависимость потерь урожая в процентах (у) от среднего числа побегов заразихи на одно растение конских бобов (х) описывается уравнением:
У - 12,4 х, [3]
однако теснота свя зи оказалась более низкой, г- 0,70. Обычно потери урожая определяются путем сравнения продуктивности пораженных и непораженных растений, отобранных из одного посева (Mesa-Garcia, Garcia-Torres, 1984; Zaitoun et al.. 1991). Полученные таким способом оценки ущерба завышаются, так как вследствие сниженной конкурентоспособности пораженных особей возрастает продуктивность растущих рядом с ними непораженных расгений. Поэтому более надежны прямые определения урожайности в разной степени пораженных посевов. Степень их поражения заразихой надежней оценивать средним числом цветоносов паразита на 1 м" посева, гак как это ценотический показатель и его использование позволяет избежать ошибок определения, на корнях каких растений паразитируют конкретные особи заразихи. Вследствие сложности проведения специальных опытов с учетом перечисленных требований мы использовали необычный подход к разработке математической модели влияния заразихи на урожайность подсолнечника.
Для оценок величин потерь урожая и числа цветоносов паразита мы обработали данные опытов по определению урожайности посевов подсолнечника при разных уровнях их поражения заразихой, результаты которых были опубликованы в разные годы различными авторами (Пустовойт, 1966. 1971; Жданов, 1964; Владимирская, 1940; Бейлин, 1967; Малыхин, 1983). Но показателям среднего числа цветоносов заразихи на 100 растений или процента пораженных растении и степени их поражения вычислили оценки числа цветоносов на I м“ посева, исходя из того, что густота опытных посевов подсолнечника не должна значительно отклоняться от величины 4 растения па I м". Урожайность пораженных заразихой посевов выразили в процентах от оценок этого признака непораженного контроля. Для тех опытов, в которых все изученные посевы были в разной степени поражены заразихой, урожайность, которую обеспечил бы в тех же условиях непораженный посев, оценивали для каждого опыта отдельно путем экстраполяции по регрессии величин урожаев на число цветоносов заразихи на I мл По откорректированным таким образом данным перечисленных авторов построили график зависимости процента снижения урожая посевов подсолнечника от числа цветоносов заразихи на 1 м" посева (рис. 1).
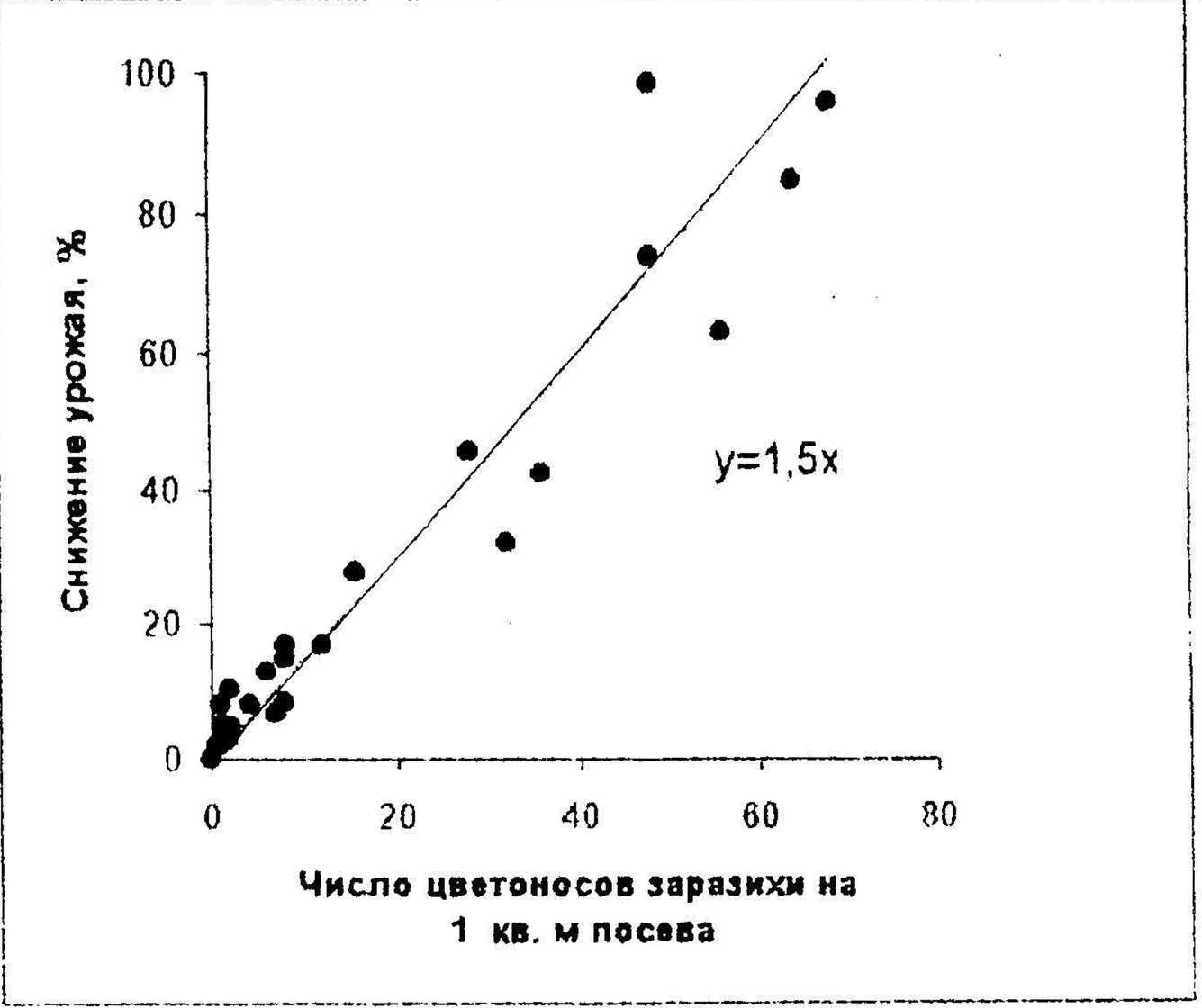
Рисунок 1. Зависимость снижения урожая семянок подсолнечника от степени поражения посевов заразихой.
Несмотря на го, что опыты проводились в разных зонах, с различными сортами подсолнечника и поражением растений заразихой разных рас, экспериментальные точки зависимости относительных оценок потерь урожая семянок (у, %) от числа цветоносов заразихи (х, шт./м") легли довольно тесно вдоль проходящей через начало координат линии регрессии
У = Ьх, [4]
где Ь--т1,5, т. е. в среднем по большому числу опытов с увеличением числа заразих в посеве подсолнечника на 1 шт./м2 урожайность семянок снижается на 1,5 %. Эта зависимость четко проявляется в диапазоне от 0 до 70 цветоносов на 1 м~ посева. В этом диапазоне коэффициент корреляции между величинами потерь урожая и чисел цветоносов составил г=0,912 (переходит ОД %-ный уровень существенности). Дальнейшее увеличение числа заразих сверх 70 цветоносов на 1 м" уже не сопровождается снижением урожайности. По-видимому, это объясняется меньшими размерами растений паразита вследствие сильной конкуренции между ними за ограниченные ресурсы пораженных растений подсолнечника.
Появление особо опасного патогена подсолнечника - возбудителя фомопсиса привело к снижению урожайности посевов этой культуры в Югославии более чем на 50 % (Maric et al., 1982). В особо благоприятных для этого патогена метеорологических условиях потери урожая достигают 70 % и более (Игнатов, 1984). Поэтому возросла зависимость оценок урожайности подсолнечника от погодных условий. Анализ результатов сортоиспытаний отдела селекции и семеноводства гибридного подсолнечника ВНИИМК показал, что в 80-е годы коэффициенты вариации оценок урожайности разных гибридов и сортов находились в пределах 3-14 %, а после появления фомопсиса на Северном Кавказе в 90-е годы - 16-29 %. По данным серии наших агроэкологических испытаний гибридов и сортов подсолнечника в 1994-1997 гг., установлено, что генотипическая изменчивость коэффициентов вариации оценок урожайности (у) и массы 1000 семянок (z) тесно положительно связана с показателем степени поражения посевов фомопсисом (х,%). Коэффициенты корреляции этих зависимостей равны соответственно г 0,90 и* г 0,84, а описываются они уравнениями регрессии:
у()387х-7,2, [51
z-435 -H)J3x. [6]
Это означает; что генотипические различия по нестабильности оценок как урожайности, гак и качества урожая в основном определяются в настоящее время разной степенью восприимчивости изученных гибридов и сортов к поражению фомопсисом. С ама же нестабильность оценок величин урожая и его качества обусловлена тем, что в отличие от заразихи степень поражения подсолнечника фомопсисом в значительной мере зависит от складывающихся погодных условий во время вегетации подсолнечника. Поэтому для прогноза развит ия эпифитогий необходимо знание количественных параметров метеорологических факторов, необходимых для спороношения возбудителя фомопенса.
Важнейшим условием интенсивного развития фомонсиса является дожщіивая погода в период бутонизации и цветения подсолнечника, когда происходит массовый вылет аскоспор (Тихонов, Пивень, 1991). Для развития злифитотии продолжительность такой погоды должна быт ь достаточной для того, чтобы рост мицелия завершился егюро-пошенпем, а эго зависит от температурных условий (Iliescu cl al., 1985). Температурные параметры роста и спороношения возбудителя фомонсиса были установлены экспериментами в кон гролируемых условиях и качественно была показана возможность их использования для прогноза эпифитотий фомонсиса в полевых посевах подсолнечника (Iliescu ct al., 1985). Эт и авторы определили, что нижний предел эффективной температуры для роста мицелия фомонсиса равен 14 °C, для спороношения - 18 °C; оптимум для роста и спороношения равен 26 °C, верхние пределы .....32 °C, сумма эффективных температур. накопление которой необходимо для начала спороношения. равна 252 °C. На основании этих данных можно построить уравнение для прогноза, при какой продолжительности влажной погоды может начаться массовый вылет аскоспор фомонсиса:
^••.ул0 -1° *п - £t° *п 171 ‘-'i и ср 1 nun/ L mm и —хр 1 mm 1'1
при 1 Ч)-'і пш, И I ЧЛ 26 , где Xt'n - сумма накопленных положительных температур ш в суток; п - число суток влажной погоды, благоприятной для развития возбудителя фомонсиса, до начала спороношения; t°cp. .....среднесуточная температура; tr,mm - нижний предел эффективной темпера гуры; У1‘\ф - сумма эффективных температур, необходимая для начала спороношения.
В связи со значительной вариабельностью интенсивности развития фомонсиса на посевах подсолнечника и уровней потерь урожая вредоносность этою патогена должна выражаться уравнением связи оценок снижения урожайности с показателями степени поражения растений. Для выявления такой зависимости мы использовали данные проведенных в 1995 г. отделом биологических исследований ВНИИМК полевых испытаний гибридов и сортов подсолнечника в условиях эпифитотийного развития болезни. Опыты проводились в 4-кратной повторности на делянках с защитными рядами и учетной площадью 23,5 мт Оказалось, что основной причиной варьирования оценок урожайности в этих опытах были наследственные различия но степени поражения фомопсисом, поскольку коэффициенты корреляции составили г- -0.843 по данным испытаний на фоне оптимальной густоты посевов 40 тыс. раст./га (переходит 0,1 %-ный уровень существенности). Результаты этого испытания представлены на графике (рис. 2). Их регрессионный анализ показал, что зависимость оценок урожайности (у, т/га) от степей и поражения растений фомопсисом (х, %) описывается уравнением:
у - 3,85 - 0,025х,
где 3,85 т/га - оценка урожайности в отсутствии поражения; 0,025 - средняя степень снижения оценок урожайности в т/га при увеличении степени поражения посевов на 1 %. В процентах к оценке урожайности непораженного посева этот коэффициент регрессии равен 0,65 % снижения урожайности при повышении поражения посева на 1 %.
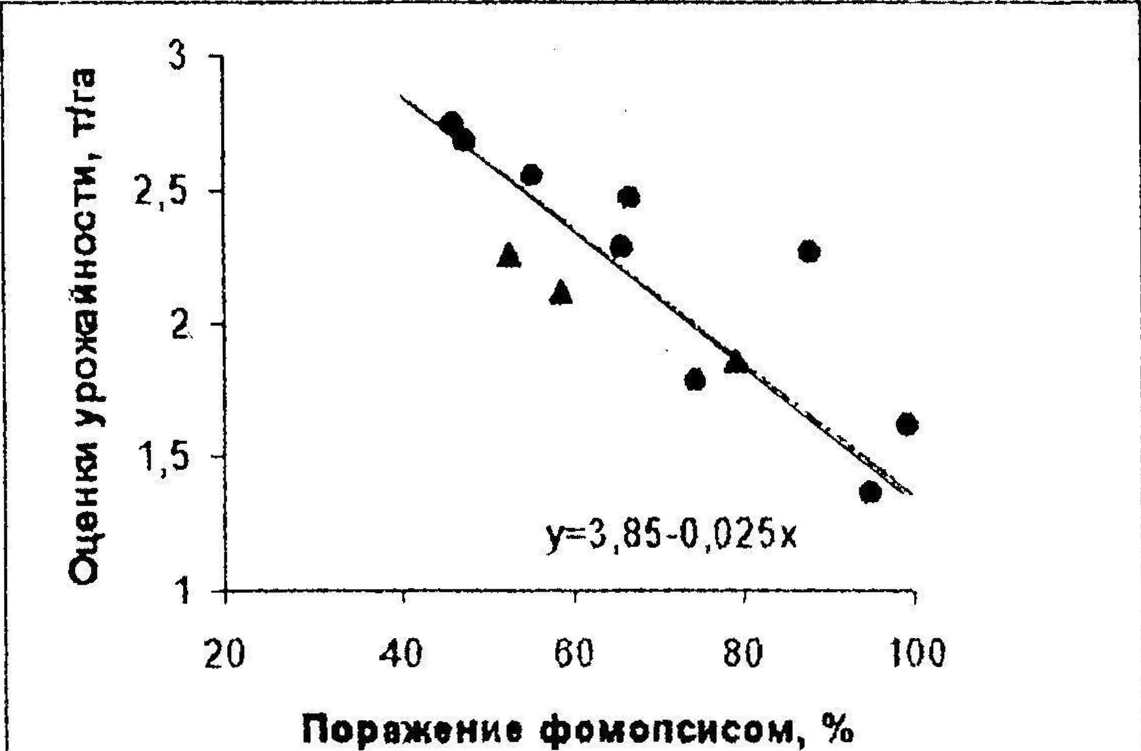
• Гибриды а Сорта---у^3.85-0,025х
Рисунок 2. Зависимость оценок урожайности гибридов и сортов подсолнечника от степени поражения фомопсисом при густоте посевов 40 тыс. раст./га.
Известно, что при поражении подсолнечника фомопсисом потери урожая избыточно загущенных посевов выше, чем при оптимальной густоте (Maric et aL, 1982; Игнатов, 1984; Iliescu, Baicti, 1984). Для выяснения того, как увеличенная густота посевов влияет на вредоносность фомопсиса, тот же набор гибридов и сортов был одновременно испытан при тех же условиях на фоне густоты посевов 80 тыс. раст./га. Тесная отрицательная зависимость между степенью поражения посевов фомопсисом и оценками урожайности проявилась и в этом опыте (г= -0,858, переходит 0,1 %-ный уровень существенности). Регресс ионный анализ результатов этого опыта, представленных на графике (рис. 3), показал, что в условиях избыточной густоты посевов снижение оценок урожайности (у, т/га) с увеличением степени поражения фомопсисом (х, %) не выше, чем при оптимальной густоте посевов. Если для данных испытания на фоне 80 тыс. раст./га принять, такой же коэффициент регрессии 0,025 т/га на 1%, как полученный для опыта при 40 тыс. раст./га, то эта зависимость для избыточной густоты описывается уравнением:
у - 3,5 -0,025х [9]
Сравнение графиков (рис. 2 и 3) и уравнений [8] и [9] свидетельствует о том, что избыточное загущение посевов снизило оценки урожайности всех генотипов независимо от их восприимчивости к фомопсису в среднем на 3,85 - 3,5^0,35 т/га, но вредоносность этого патогена в условиях загущенных посевов оказалась не выше, чем при густоте
40 тыс. раст./га. Кажущееся несоответствие этого вывода литературным данным объясняется тем, что уборка урожая в наших опытах производилась вручную со сбором корзинок растений со сломленным стеблем. Такие корзинки теряются при комбайновой уборке, а растений с переломленным стеблем было гем больше, чем сильнее было поражение фо-мопсисом, особенно при избыточной густоте посевов. Это дает основание полагать, что повышенная вредоносность фомопсиса при избыточном загущении посевов подсолнечника обусловлена более высокими потерями урожая при комбайновой уборке густых посевов, пораженных этой болезнью.
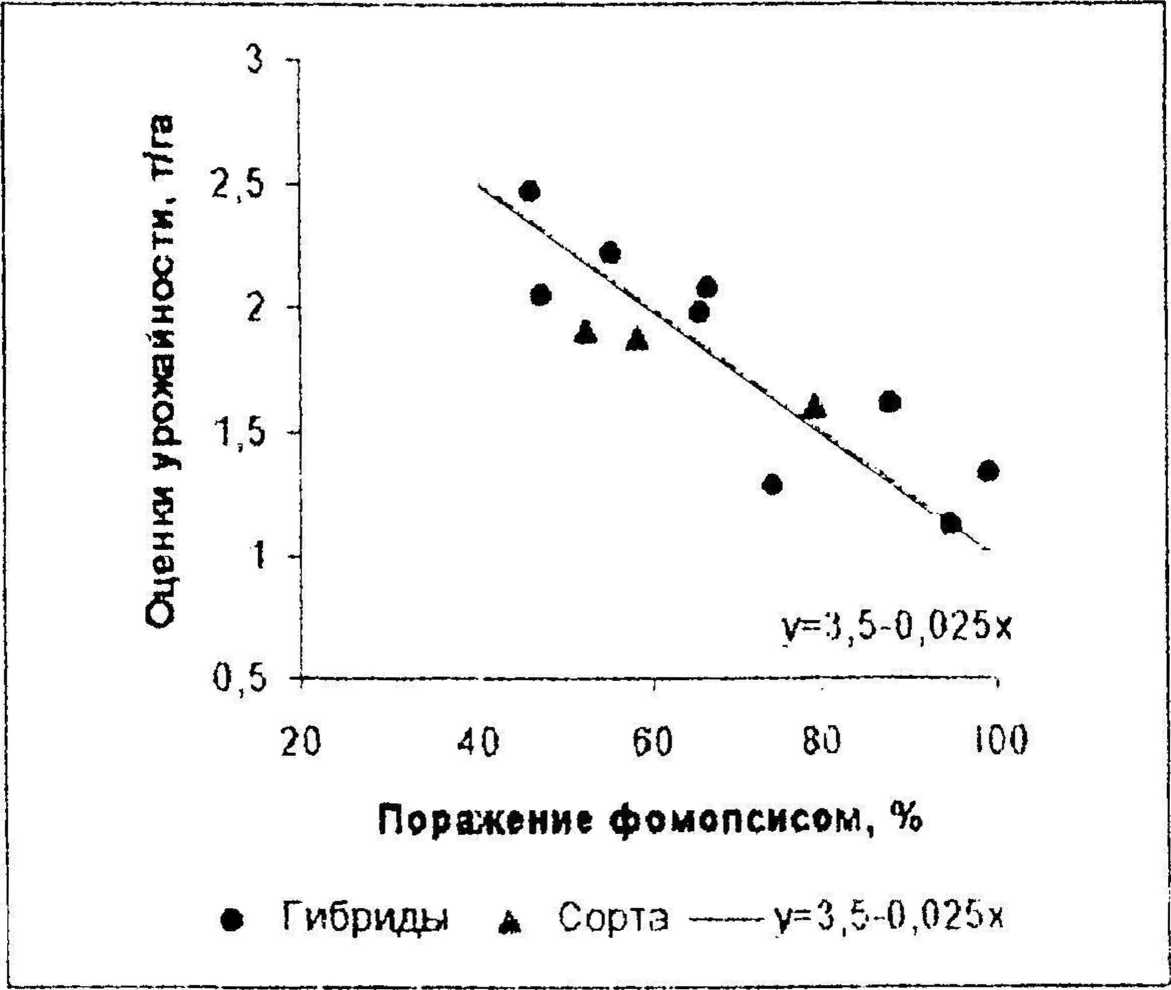
Рисунок 3. Зависимость оценок урожайности гибридов и сортов подсолнечника от степени поражения фомопсисом при густоте посевов 80 тыс. раст./га.
Хотя зависимость урожайности гибридов и сортов подсолнечника от степени поражения посевов фомопсисом по данным наших опытов высока, но она не полная: коэффициенты детерминации оценок урожайности процентом пораженности посевов составили Cd^69,6 % и Cd^73,6 %. Остальная часть вариации величин урожайности обу- словлена частично неодинаковым потенциалом продуктивности генотипов, в том числе за счет разной продолжительности вегетации, в какой-то мере - экспериментальными погрешностями. В то же время при испытаниях этого набора гибридов и сортов в разные годы на различных фонах были обнаружены воспроизводимые отклонения генотипов от соответствующих линий регрессии. Это свидетельствует о наличии наследственных различий по оценкам урожайности при равной степени поражения фомопсисом за счет разной толерантности гибридов и сортов к воздействию патогена (Тарр, 1975, с. 510). Для оценки этого свойства у гибридов и сортов нами (Дьяков, Шарыгина, 1997) разработан способ вычисления индекса толерантности гибрида или сорта:
\00*Ywi*Yw.Г‘ YdMYd.
[Ю]
где Ті - индекс толерантности і-го гибрида или сорта к фомопсису, %; Ywi и Ydi - оценки урожайности і-го гибрида или сорта по данным иены ганий при влажных и сухих погодных условиях соответственно, т/га; Yw. и Yd. -- средние оценки урожайности всех гибридов и сортов тех же испытаний, т/га.
В условиях энифитогий фомопсиса выявляются большие различия между межлинейными гибридами подсолнечника по степени восприимчивости к этой болезни. Сорта-популяции различаются по этому признаку в меньшей степени. Например, наиболее восприимчивый гибрид в испытании 1995 г. (см. рис. 2) был поражен фомопсисом в 2,2 раза сильней, чем самый устойчивый, a copra различались в 1,5 раза. По оценкам урожайности такие гибриды различались в 2 раза, a copra - в 1,2 раза. В меньшей степени различались средние показа гели: средняя урожайность по группе гибридов 2,19 т/га, сортов -2,07 т/га; средняя степень поражения фомопсисом посевов гибридов 70,9 %, сортов -63 Л %. Эти данные свидетельствуют о том, что в генофонде популяций подсолнечника селекции ВНИИМК содержатся гены, оказывающие как положительное, так и отрицательное влияние на устойчивость к фомопсису. По этой причине при создании инбредных линий подсолнечника на базе таких популяций происходит значительная дифференциация генотипов по этому признаку. Это означает также, что рекуррентной селекцией популяций подсолнечника в этом направлении можно в значительной степени повысить концентрацию генов, обеспечивающих высокую степень устойчивости подсолнечника к фомопсису. Использование таких популяций в качестве исходного материала при селекции родительских линий с учетом этого признака должно обеспечить создание полностью устойчивых к фомопсису гибридов подсолнечника. Эффективность такой программы селекции зависит о г степени наследуемости этого признака, поскольку он находится под 11 о.11 и гс и и ы м ко 1 ггрол е м.
Поскольку оценки коэффициентов наследуемости, полученные корреляционным и регрессионным методами, не только не совпадают, но и содержат различающуюся информацию. мы использовали оба эти метода. Для повышения надежности оценок за счет выявления всего адди г и в но го (генетического) компонента генотипической вариации мы анализировали ковариацию значений степени устойчивости межлинейных гибридов F] с тем же признаком не одного родителя, а среднеродительскими значениями этого признака. В этом случае коэффициенты наследуемости в узком смысле (1г) равны неудвоенным величинам коэффициента корреляции и коэффициента регрессии (Плохинский, 1964, с. 27, 39; Рокицкий, 1978, с. 245, 294). Проведенный анализ показал, что зависимость восприимчивости к фомопсису гибридов Fi (у, %) подсолнечника от значений усредненной восприимчивости к этому патогену пар родительских линий (х, %) прямолинейная (рис. 4), описывается уравнением:
у - 37 । 0,56х, [11]
а теснота связи оценивается коэффициентом корреляции г-0,853 (переходит I %-ный уровень сущеетвенности). Поэтому и оцененный корреляционным методом коэффициент наследуемоети в узком смысле равен Ь^0,853. Из этого следует, что даже при неизбежных погрешностях оценок степени поражения патогеном родителей и потомков, доля аддитивной (генегической) изменчивости в общей фенотипической вариации очень велика по сравнению с наследуемостью других хозяйственно важных полигонных признаков, оцениваемой в системе родители/потомки. Это является предпосылкой высокой эффективности правильно спланированной селекционной программы отборов на такой признак (Рокицкий, 1978, с. 228, 241).
Оцененный регрессионным методом коэффициент реализуемой наследуемости признака устойчивости подсолнечника к фомопсису оказался также относительно высоким, 1г Ьн/р-0,56. Это свидетельствует как о достаточной генетической изменчивости этого признака в популяции родителей (Плохинский, 1964, с. 38), гак и о том, что селек- ционный сдвиг в желаемом направлении даже при однократном отборе может составлять более половины селекционного дифференциала по этому признаку (Рокицкий, 1978, с. 296-297), а также о том, что отборы на устойчивость к фо мопс и су при селекции родительских линий должны эффективно обеспечивать создание устойчивых гибридов.
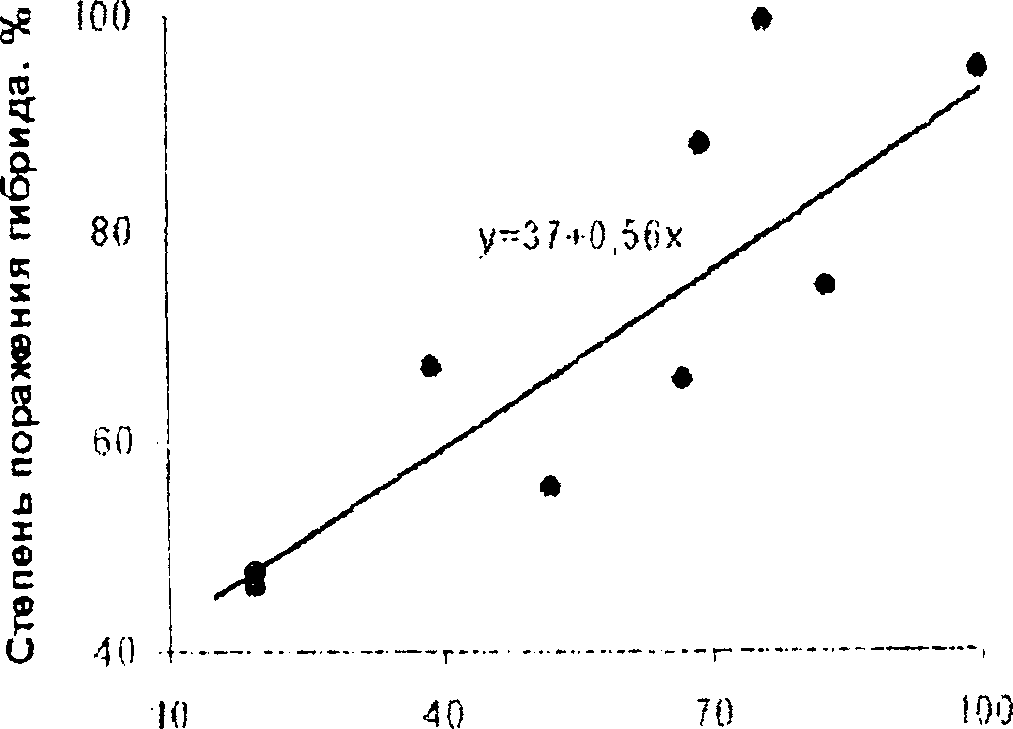
Среднеродительская степень поражения, %
Рисунок 4. Регрессионная оценка реализуемой наследуемости степени восприимчивости подсолнечника к поражению фомопсисом.
Гаким образом, количественные подходы с использованием математических методов позволяют повышать эффективность исследований взаимоотношений возделываемых растений с патогенными организмами. На ряде конкретных примеров показано, что математическое моделирование позволяет получать результаты, которые не могут быть получены на основе использования качественных или полуколичественных методов исследования. Продемонстрированы возможности математического описания развития процессов поражения растений патогенами, прогноза эпифитотий. зависимостей потерь урожая от степени поражения посевов в разных агроэкологических условиях, что необходимо для вычисления порогов вредоносности в различных экономических ситуациях. Кроме гого показана возможность и степень эффективности селекции на полигенно контролируемую устойчивость к патогену, предлагается способ оценки толерантности генотипов к поражению болезнью.
