Использование особенностей гриба Macrophomina phaseolina (Tassi) Goid. в селекции сои на толерантность к пепельной гнили (сообщение II)
 Goid. в селекции сои на толерантность к пепельной гнили (сообщение II)")
Автор: Саенко Г.М., Зеленцов С.В., Пивень В.Т.
Рубрика: Селекция и семеноводство
Статья в выпуске: 2 (155-156), 2013 года.
Бесплатный доступ
Показано, что микросклероции возбудителя пепельной гнили Macrophomina phaseolina (Tassi) Goid. могут формироваться не только из сплетения нескольких гиф, но и из отдельных гиф, что повышает риск трахеомикозного увядания растений сои. Вредоносность пепельной гнили проявляется в условиях водного стресса на фоне повышения концентрации клеточного сока в тканях растения-хозяина. При этом в тканях растений сои массово образуются микросклероции, полностью закрывающие просветы сосудов проводящей системы сои. Рост гриба прекращается при достижении изотонического уровня осмотического давления питательной среды около 930 кПа. Дальнейшее увеличение осмотического давления питательной среды вызывает массовое образование микросклероциев. Разработан метод отбора толерантного к болезни исходного материала для селекции сои на основе выделения форм растений с осмотическим давлением клеточного сока, не превышающим 930 кПа в фазы бобообразования и налива семян.
Соя, селекция на толерантность, микоз, пепельная гниль, микросклероции, осмотическое давление клеточного сока
Короткий адрес: https://sciup.org/142151147
IDR: 142151147 | УДК: 632.9:633.853.52
Using of fungi Macrophomina phaseolina (Tassi) Goid features in soybean breeding for tolerance to charcoal rot (report II)
The report shows that microslerotia of charcoal rot Macrophomina phaseolina (Tassi) Goid. may be formed not only from a plexus of hyphae, but also from separate hyphae, which increases the risk of fungal wilting of soybean plants. The harmfulness of charcoal rot is developing in conditions of water stress against the background of a higher concentration of the cell juice in the tissues of a host plant. In which case microslerotia begin mass formation in the plant tissues, completely closing the openings of vessels of soybean conducting system. Fungal growth stops when the isotonic level of osmotic pressure of nutrient medium is about 930 kPa. A further increase in the osmotic pressure of the nutrient medium causes mass formation of microsclerotia. There was developed the method of selection of breeding material tolerant to the disease for breeding soybean on the basis of plant sampling with the osmotic pressure of the cell juice of not more than 930 kPa in a phase of pod formation and seed ripening.
Текст научной статьи Использование особенностей гриба Macrophomina phaseolina (Tassi) Goid. в селекции сои на толерантность к пепельной гнили (сообщение II)
Введение. В европейской части России лидирующим по производству сои регионом является Краснодарский край, территориально входящий в зону Западного Предкавказья. В 2012 г. на площади 172 тыс. га при урожайности 18,5 ц/га в крае был получен валовой сбор сои более 300 тыс. т.
В то же время рост площадей и производства сои на юге России сопровождается увеличением распространения опасных микозов, способных нанести значительный ущерб урожаю и качеству семян. Одним из распространённых и вредоносных заболеваний сои является пепельная гниль, возбудитель которой – гриб Mac-rophomina phaseolina (Tassi) Goid.
Пепельная гниль в посевах сои наиболее часто проявляется при жаркой сухой погоде, когда неблагоприятные условия внешней среды вызывают стресс у растений. Участившиеся в последние десятилетия на Северном Кавказе, включая Западное Предкавказье, продолжительные и интенсивные летние засухи увеличивают вероятность образования благоприятных условий для развития М. phaseolina в промышленных посевах сои [3]. При сохранении тенденций изменения климата в Западном Предкавказье, с учётом дальнейшего увеличения посевных площадей под сою в среднесрочной перспективе, предполагается нарастание эпифитотийного распространения и развития болезни в регионе. Складывающаяся ситуация обусловила необходимость уточнения биологии возбудителя пепельной гнили – гриба M. phaseolina и создание сортов сои с повышенной устойчивостью к патогену.
Несмотря на распространённость возбудителя пепельной гнили, в мире до настоящего времени у сои так и не обнаружена генетическая устойчивость к этому патогену и практически отсутствуют иммунные к М. phaseolina сорта сои [13; 9; 11;14].
Количество исследований, посвященных биологии, распространённости и вредоносности гриба М. phaseolina на сое, особенно в зоне Западного Предкавказья и Северного Кавказа в целом, крайне ограничено. Опубликованные сведения имеют фрагментарный и незавершённый характер. В литературе практически отсутствуют данные о наличии генотипов сои с вертикальной или горизонтальной устойчивостью к этому патогену. Слабо изучена биология гриба и практически не изучены условия образования микроскле-роциев в системе патоген–растение-хозяин в стрессовых условиях острой засухи.
В связи с возрастающей вероятностью наступления в летний период продолжительных и интенсивных засушливых периодов, изучение распространённости и развития пепельной гнили на сое, биологических особенностей гриба М. phaseolina, а также поиск эффективных мероприятий по снижению вредоносности пепельной гнили, включая создание исходного материала для селекции сортов сои с повышенной устойчивостью к М. phaseolina , являются актуальными.
Поэтому целью настоящих исследований явилось изучение биологии гриба М. phaseolina в онтогенезе сои, выделение толерантных к возбудителю пепельной гнили форм, а также создание на их основе исходного материала для селекции сортов сои в условиях Западного Предкавказья.
Материал и методы. Основные учёты и исследования посевов сои и возбудителя пепельной гнили Macrophomina phaseolina (Tassi) Goid. проводили в период 2007–2010 гг. на центральной экспериментальной базе Всероссийского научно-исследовательского института масличных культур имени В.С. Пусто-войта (ВНИИМК), г. Краснодар, Российская Федерация.
При выполнении полевых и лабораторных исследований наблюдения за растениями, их измерения и учёты проводили в соответствии с методическими указаниями по селекции сои, принятыми во ВНИИМК [4].
Массовые учёты распространённости и развития пепельной гнили проводили на селекционном участке отдела сои ВНИИМК. Материалом для исследований на устойчивость к пепельной гнили были районированные и перспективные сорта сои, селекционный материал лаборатории селекции сои ВНИИМК, образцы мировой коллекции ВИР.
Объектом изучения в данной работе являлся поражающий сою гриб Mac-rophomina phaseolina (Tassi) Goid. Растения с признаками проявления болезни отбирали в селекционных питомниках и на семеноводческих посевах. Изоляцию патогена в чистую культуру проводили с использованием методик выделения фи-топатогенных грибов из различных органов растений [8; 5; 1]. Для выделения патогена в чистую культуру использовали методики И.Я. Жербеле и С.Ф. Сидорова [2; 6].
Культивировали гриб на картофельноглюкозном агаре (КГА) и среде Чапека. Оптимальные температуры определяли при инкубации изолятов пепельной гнили на КГА. Необходимые температурные режимы поддерживали при помощи термостата.
Для учёта распространённости болезни проводили маршрутные обследования посевов сои диагональным методом, путём подсчёта числа растений на 1 погонном метре в 10–20 местах. Учёты по развитию болезни на мелкоделяночных опытах проводили по двум средним учётным рядам в делянке.
Для исследования динамики осмотического давления клеточного сока (ОДКС) в тканях сои в течение онтогенеза использовали рефрактометры ИРФ-22 и PR-101 a по методике Н.Н. Третьякова [7]. Особенности развития гриба в условиях водного стресса оценивали путём выращивания мицелия гриба на жидкой питательной среде Чапека с различными концентрациями высокомолекулярного нейтрального осмотического вещества полиэтилен-оксида (ПЭГ-6000) на основе методики M.L. Ramirez и S.N. Chulze [12].
Результаты и обсуждение. При изучении симптомов проявления пепельной гнили на сое в 2007–2010 гг. было установлено, что на фоне относительно благоприятных для роста и развития сои погодных условий первые симптомы про- явления болезни появились в фазы бобо-образования и налива семян.
В период физиологического созревания растений сои симптомы развития болезни наблюдали на поверхности прикорневых частей стеблей и корнях. При анатомическом исследовании растений микросклероции М. phaseolina обнаруживали во всех тканях стебля (рис. 1).
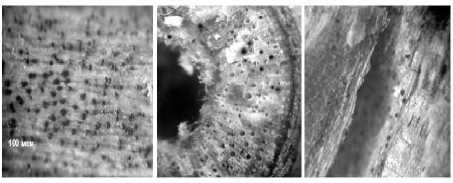
Рисунок 1 – Локализация микросклероциев в тканях стебля сои (ориг.): а – микросклероции в коре; б – в древесине (отдельные микросклероции закупорили сосуды ксилемы); в – на границе между древесиной и сердцевиной (ткани сердцевины полностью разрушены грибом)
Большое их количество обнаруживали во внутренних тканях коры и древесины. Наличие микросклероциев на внутренней поверхности древесины, примыкающей к сердцевине, свидетельствует о развитии мицелия гриба и в этой части стебля. Во всех тканях размеры микросклероциев были одинаковыми и в среднем составляли 50–75 мкм. Наиболее вредоносным представляется образование микроскле-роциев в древесине, которое вызывает закупорку проводящей сосудистой системы растений и приводит к нарушению водоснабжения, питания и к преждевременной гибели растений сои.
Оптимальной температурой для роста и развития мицелия гриба M. phaseolina, является интервал 25–30 °С, а для интенсивного образования микросклероциев – 27–35 °С. Оптимальная кислотность среды, при которой наиболее быстро развивается мицелий и формируются микросклероции составляет: на КГА – 4,5–6,0 рН, на среде Чапека – 5,5–6,0 рН.
При исследовании механизма образования в тканях сои микросклероциев гриба M. phaseolina в качестве рабочей гипотезы нами было принято их образование из отдельных гиф на любом пригодном для гриба питательном субстрате. Для подтверждения гипотезы были проведены более детальные исследования формирования микросклероциев в динамике (рис. 2).
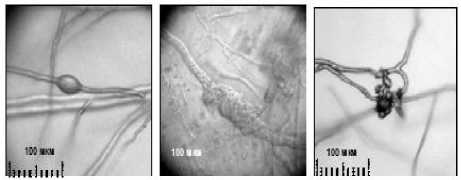
а б в
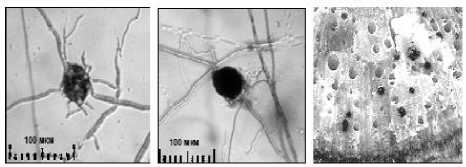
г д е
Рисунок 2 – Этапы формирования микросклероциев М. phaseolina из отдельных гиф, х 200, (ориг.): а – первичная протосклероциевая капсула;
б – септирование протосклероциевой капсулы;
в – начало агрегирования клеток капсулы в микросклероций; г – «незрелый» микросклероций; д – полностью сформированный микросклероций; е – микросклероции в сосудах ксилемы
Начальным этапом формирования микросклероциев является образование на отдельных гифах сферических, недифференцированных утолщений – первичных протосклероциевых капсул (рис. 2а). Затем первичная капсула формирует перегородки (септы) с расчленением на отдельные клетки (рис. 2б). Септированные клетки капсулы продолжают делиться, постепенно агрегируются в молодые микросклероции, приобретая тёмную окраску из-за накопления меланина (рис. 2в). К моменту созревания микросклероциев они приобретают типичный для пепельной гнили серебристо-чёрный цвет. Причём меланиновая окраска равномерно распространяется по всей толще микросклероция (рис. 2г, д) протосклероциевые капсулы, а в последствии микросклероции могут формироваться в гифах как ин-теркалярно, так и терминально.
В целом, проведённые исследования показали, что для закупорки микроскле-роциями проводящей сосудистой системы растений достаточно наличия одной гифы в сосуде (рис. 2 е ).
При исследовании динамики образования и роста микросклероциев в пределах одних и тех же фиксированных гиф было установлено, что при оптимальной температуре активный рост мицелия патогена на среде Чапека начинается уже на 2-е сутки. По мере истощения среды и постепенного снижения её влажности, как правило, на 4-е сутки, начинается образование первичных капсул на отдельных гифах. На 6-е сутки из капсул формируются микросклероции, которые с возрастом увеличиваются до типичных размеров и темнеют от накапливающегося меланина.
В 2007–2009 гг. на центральной экспериментальной базе ВНИИМК было изучено влияние погодных условий на распространённость и развитие пепельной гнили на сое.
Трёхлетние обследования разнообразного генетического материала сои показали, что повышенные температуры воздуха перед массовым созреванием сои способствуют увеличению развития пепельной гнили на ней. Так, в 2007 г. развитие болезни составило 25–45 %; в 2008 г. – 18–41; в 2009 г. – 17–37 %. Распространённость составила 100, 90 и 70 % соответственно.
Анализ многочисленных литературных данных, а также собственных наблюдений за распространением и развитием пепельной гнили на сое показывает, что признаки болезни максимально проявля- ются на культуре в острозасушливые годы, когда в тканях растений из-за дефицита воды повышается концентрация клеточного сока.
На основании выявленных взаимосвязей между влажностью окружающей среды и образованием микросклероциев нами была сформулирована рабочая гипотеза осмотически зависимого образования микросклероциев у М. phaseolina , основанная на градиенте осмотического давления внутри и вне мицелия гриба.
Согласно этой гипотезе образование микросклероциев в значительной степени обусловлено гипертоническим ОДКС внутри мицелия гриба относительно ОДКС тканей растения-хозяина. При развитии водного стресса ОДКС тканей растения-хозяина увеличивается и достигает изотонических значений к ОДКС внутри мицелия. При изотонических значениях ОДКС мицелия патогена и тканей растений для гриба наступает «физиологическая засуха», провоцирующая массовое образование микросклероциев. Образовавшиеся микросклероции закупоривают сосуды ксилемы, последовательно вызывая угнетение, завядание и гибель растения-хозяина.
В естественных условиях микросклероции пепельной гнили обычно формируются на поздних этапах развития сои. В то же время в рамках осмотической гипотезы при наличии мицелия в тканях сои при критических значениях ОДКС микросклероции должны образовываться независимо от возраста растений, почти сразу после прорастания мицелия в растительных тканях (см. Сообщение I в настоящем журнале на стр. 27).
Проведённые эксперименты подтвердили наше предположение о потере тканями растений воды и возрастании ОДКС клеточного сока, как основной причине образования микросклероциев пепельной гнили.
Следовательно, в растениях сои, ОДКС которых не достигает изотонических значений с ОДКС гриба, даже при наличии мицелия в растительных тканях, не должны образовываться микросклероции, либо они будут формироваться уже на стадии физиологического созревания растений и не окажут негативного влияния на урожай семян.
Было выдвинуто предположение, что если в пределах доступного генофонда сои существует генетически детерминированный полиморфизм по ОДКС тканей сои, то разработка и осуществление программы отбора генотипов с гипотоническим ОДКС позволит повысить устойчивость сои к пепельной гнили.
Первым этапом при реализации этой программы стало определение осмотического давления мицелия гриба. Прямое определение ОДКС внутри гиф гриба представляет собой технически сложную задачу. Однако критическое для роста и развития патогена ОДКС можно определить через культивирование культуры гриба в жидких питательных средах с добавлением различного количества осмотически активных веществ типа полиэтиленгликоля ПЭГ-6000. При этом осмотическое давление (ОД) среды, при котором полностью прекратится рост мицелия патогена, будет указывать на изотонические уровни ОДКС внутри и вне мицелия.
Исследования показали, что ОД жидкой питательной среды Чапека составляет 220 кПа. В такой среде объём мицелия М. phaseolina на 4-е сутки культивирования в среднем достигал 520 см3. Дальнейшее увеличение концентрации ПЭГ-6000 в жидкой среде вызывало постепенное угнетение роста мицелия гриба (табл. 1).
При увеличении ОД среды до уровня 710–795 кПа скорость прироста мицелия гриба уменьшалась. Дальнейшее увеличение ОД среды до уровня 860 кПа приводило к 10-кратному сокращению прироста мицелия.
Критическим осмотическим давлением среды, при котором была зафиксирована остановка роста мицелия в эксперименте, оказался уровень 930 кПа, формируемый 9,5 %-ной концентрацией ПЭГ-6000.
Таблица 1
Зависимость роста мицелия в объёме жидкой среды Чапека от концентрации полиэтиленгликоля (ПЭГ-6000)
|
ВНИИМК, 2007–2008 гг. |
||||
|
Концентрация осмотического вещества, % |
Осмотическое давление раствора, кПа |
Рост зоны объёма мицелия, см3 |
||
|
2-е сутки |
3-и сутки |
4-е сутки |
||
|
Контроль (без ПЭГ-6000) |
220 |
33,5–113,0 |
113,0 |
520,3 |
|
2,0 |
490 |
33,5–113,0 |
113,0 |
523,3 |
|
7,5 |
710 |
33,5–113,0 |
113,0–179,5 |
267,9 |
|
8,0 |
795 |
4,0 |
107,3 |
214,6 |
|
8,5 |
840 |
3,5 |
33,5 |
33,5–65,4 |
|
9,0 |
860 |
3,5 |
33,5 |
33,5–65,4 |
|
9,5 |
930 |
Слабый рост мицелия |
Плотный мицелий на семянке |
Плотный мицелий на семянке |
|
10,0 |
1100 |
Нет роста на среде |
Нет роста на среде |
Нет роста на среде |
Дальнейшее увеличение ОД среды полностью блокировало рост мицелия и, следовательно, являлось гипертоническим к внутреннему ОДКС гриба.
Таким образом, осмотическое давление среды, равное 930 кПа, при котором полностью прекращался рост гриба M. phaseolina , было принято за изотоническое, при котором осмотическое давление становится равным внутри и вне мицелия.
В связи с суточной динамикой температур и влажности воздуха и явной зависимостью от них ОДКС растений сои была поставлена задача разработки полевого метода определения этого параметра и установления оптимального периода суток для его определения.
За основу полевого экспресс-метода был взят рефрактометрический анализ, позволяющий определить ОДКС через концентрацию растворённых в клеточном соке веществ.
При помощи ручного пресса получали клеточный сок из каждого образца. На рефрактометре определяли показатель его преломления с учётом температурного коэффициента. Затем на основе справочных таблиц Н.Н. Третьякова [7] находили величину осмотического давления, соот- ветствующую найденной концентрации клеточного сока.
Технология экспресс-метода определения ОДКС в тканях стеблей сои включала следующие этапы:
– отбор 3–5 типичных растений на каждой делянке на расстоянии не менее 0,5 м от края делянки;
– измельчение секатором стеблей отобранных растений до полного заполнения рабочей камеры пресса;
– отжим ручным прессом 1–3 мл клеточного сока из измельчённых стеблей сои;
– определение концентрации клеточного сока, помещённого на призму рефрактометра ИРФ-22 или в рабочую камеру рефрактометра PR-101 a;
– расчёт ОДКС всех образцов в лабораторных условиях.
Хронометраж времени показал, что на выполнение первых четырёх этапов при определении концентрации клеточного сока в одном сортообразце требуется не более 3–5 мин. Таким образом, используемый экспресс-метод позволяет за 4 часа в поле определить ОДКС 40–50 сортооб-разцов.
Для определения оптимального периода в течение полных суток с интервалом в три часа оценивали ОДКС в тканях нижнего, среднего и верхнего ярусов растений сои (рис. 3).
Растения сорта сои Лира на дату определения суточной динамики ОДКС (16.07.2007 г.) полностью завершили свой рост и находились в критической по вла-гопотреблению фазе налива семян.
Среднесуточное значение ОДКС у сорта Лира составило 700 кПа. Этот уровень оказался гипотоническим к ОДКС возбудителя пепельной гнили (930 кПа) и не провоцировал формирование микроскле-роциев.
Наиболее близкий к среднесуточному уровень ОДКС стабильно поддерживали ткани нижнего яруса растений в период с 9 ч утра до 21 ч вечера, а также ткани среднего и верхнего ярусов – в ночные и утренние часы.
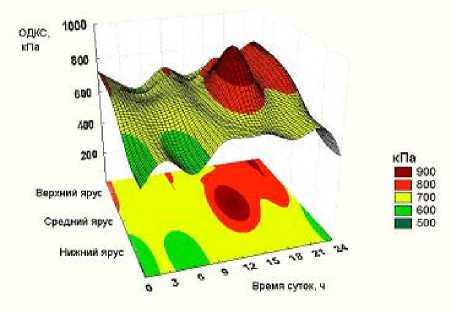
Рисунок 3 – Суточный ритм ОДКС растений сои сорта Лира в фазе налива семян, кПа
С учётом установленных периодов и ярусов растений, позволяющих наиболее объективно оценить ОДКС на различных сортообразцах, в течение 3 лет (2007– 2009 гг.) оценивали этот параметр в фазы начала цветения, формирования бобов, налива семян и физиологического созревания путём отжима клеточного сока из измельчённых тканей прикорневой части главных побегов 3–5 типичных растений каждого сортообразца.
Проведённые исследования показали, что ОДКС тканей растений в начале цветения заметно варьировало у разных сортообразцов. На примере сортов питомника экологического сортоиспытания показано, что минимальным оно было в фазе цветения, варьируя по годам от 513 до 609 кПа (табл. 2).
К фазе формирования бобов ОДКС в среднем по опыту увеличилось до 646 кПа с варьированием этого показателя от 554 до 758 кПа. В период налива семян среднее за три года ОДКС достигло уровня 729 кПа с варьированием признака от 550 до 989 кПа. В фазе физиологического созревания отдельные сорта в 2007 и 2008 гг. достигали субкритических и критических значений (более 1300 кПа) ОДКС и находились в стадии завядания.
Выявленные различия по ОДКС, а также ранги между сортами в целом сохранялись во все годы исследований, свидетельствуя о наличии наследования разных уровней ОДКС.
Проведённые исследования позволили выделить три основных типа динамики ОДКС в онтогенезе, условно названных как типы А, Б и В (табл. 3).
Тип А характеризуется достижением растениями сои гипертонических значений ОДКС по отношению к возбудителю пепельной гнили в тканях в фазы бобооб-разования и налива семян.
Тип Б характеризуется достижением растениями сои изотонических по отношению к грибу значений ОДКС в эти же фазы онтогенеза.
Тип В представляют собой сортообраз-цы, сохраняющие гипотонические для пепельной гнили значения ОДКС вплоть до начала физиологического созревания растений.
Кроме основных, были выделены промежуточные типы динамики ОДКС. В острозасушливый 2007 г. некоторые сортообразцы в фазы бобообразования и налива семян достигали уровня 930 кПа, в наиболее благоприятном по увлажнению 2009 г. – имели значения несколько ниже критического уровня. Такие сорта были отнесены к промежуточному типу АБ.
ОДКС ещё одной группы сортов в период бобообразования и налива семян варьировало в пределах 650–690 кПа. Группа таких сортов была отнесена к промежуточному типу БВ.
В целом, совокупность полученных в течение трёх лет экспериментальных данных позволила сформулировать основное условие отбора исходного материала для селекции сои на устойчивость к возбудителю пепельной гнили (рис. 4).
Динамика ОДКС в тканях нижнего яруса растений у генотипов сои в питомнике экологического сортоиспытания, кПа
ЦЭБ ВНИИМК, 2007–2009 гг.
|
Сорто-образец |
Дата и онтогенетическая фаза на момент определения ОДКС, день, месяц, год |
|||||||||||
|
цветение |
бобообразование |
налив семян |
физиологическое созревание |
|||||||||
|
14.06. 2007 г. |
16.06. 2008 г. |
17.06. 2009 г. |
04.07. 2007 г. |
01.07. 2008 г. |
01.07. 2009 г. |
25.07. 2007 г. |
28.07. 2008 г. |
31.07. 2009 г. |
30.08 2007 г. |
01.09 2008 г. |
20.08. 2009 г. |
|
|
Ланцетная |
620 |
520 |
420 |
700 |
590 |
520 |
1180 |
700 |
550 |
- |
- |
- |
|
Свапа |
540 |
440 |
440 |
660 |
490 |
470 |
950 |
870 |
550 |
- |
- |
- |
|
Лира |
480 |
480 |
440 |
620 |
630 |
480 |
710 |
640 |
570 |
- |
- |
- |
|
Белогорская 143 |
590 |
610 |
450 |
730 |
650 |
540 |
920 |
650 |
530 |
- |
- |
650 |
|
Воронежская 31 |
550 |
580 |
470 |
730 |
650 |
430 |
990 |
630 |
600 |
- |
- |
- |
|
Брянская Мия |
600 |
580 |
400 |
730 |
620 |
650 |
1050 |
540 |
610 |
- |
- |
- |
|
Славия |
540 |
370 |
540 |
780 |
560 |
660 |
1060 |
700 |
610 |
- |
- |
780 |
|
Амфор |
650 |
460 |
450 |
800 |
660 |
650 |
910 |
670 |
510 |
- |
1340 |
490 |
|
Альба |
600 |
510 |
510 |
660 |
650 |
450 |
950 |
740 |
540 |
- |
630 |
800 |
|
Селекта 201 |
670 |
530 |
460 |
770 |
680 |
820 |
950 |
730 |
540 |
- |
910 |
620 |
|
Парма |
680 |
540 |
535 |
690 |
710 |
544 |
1080 |
580 |
544 |
1320 |
1010 |
556 |
|
Рента |
670 |
600 |
530 |
680 |
620 |
460 |
850 |
640 |
560 |
1010 |
720 |
710 |
|
Вилана |
590 |
510 |
450 |
820 |
650 |
550 |
800 |
690 |
420 |
690 |
740 |
620 |
|
Среднее |
609 |
513 |
468 |
758 |
626 |
554 |
989 |
647 |
550 |
1090 |
881 |
655 |
Таблица 3
Осмотическое давление клеточного сока тканей некоторых сортов сои в онтогенезе
ЦЭБ ВНИИМК, 2007–2009 гг.
|
Сорт |
Тип онтогенетической динамики ОДКС * |
ОДКС в основные фазы развития растений, кПа |
||||
|
1-й настоящий лист |
полное цветение |
бобообразо-вание |
налив семян |
физиологическое созревание |
||
|
Амфор |
Тип А |
560 |
760 |
1170 |
1100 |
700 |
|
Армавирская 2 |
530 |
740 |
1220 |
1090 |
660 |
|
|
Славия |
Тип АБ |
490 |
660 |
870 |
930 |
590 |
|
Изидор |
510 |
690 |
820 |
920 |
620 |
|
|
Вилана |
Тип Б |
540 |
650 |
800 |
830 |
540 |
|
Рента |
510 |
600 |
820 |
860 |
520 |
|
|
Альба |
Тип БВ |
420 |
510 |
650 |
670 |
430 |
|
Валента |
Тип В |
380 |
460 |
650 |
630 |
400 |
|
Белгородская 6 |
450 |
410 |
650 |
600 |
420 |
|
*– тип динамики ОДКС устанавливается в критические по влагообеспеченности периоды плодообразования и налива семян
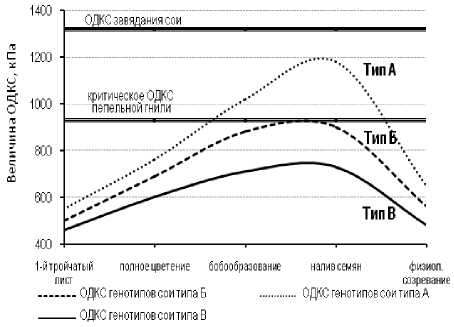
Рисунок 4 – Особенности онтогенетической динамики осмотического давления клеточного сока в тканях растений различных типов сои
Это условие заключается в том, что в фазы бобообразования и налива семян выделяют формы растений с осмотическим давлением клеточного сока не более 930 кПа, который является критическим для возбудителя пепельной гнили и приводит к массовому формированию мик-росклероциев этого гриба.
В 2007 г. при первичной оценке ОДКС сортообразцов в питомнике экологического сортоиспытания было обнаружено, что среднеспелый сорт Парма отличался от всех остальных сортов высокой нестабильностью оценок ОДКС, достигающего 80–100 кПа между отдельными растениями, при 10–20 кПа во всех остальных случаях.
Поскольку сорт Парма фенотипически выровнен, то было предположено, что повышенная нестабильность оценок ОДКС могла обуславливаться гетерогенностью по признаку величины ОДКС.
В семеноводческих посевах сорта Парма были отобраны две группы по 100 растений без видимых признаков поражения пепельной гнилью и с максимальным поражением патогеном не только в прикорневом, но и в среднем ярусе. Внутригрупповые ОДКС также отличались друг от друга на 80–100 кПа. Из потомств этих групп растений были выделены толерантные и восприимчивые сублинии.
С целью подтверждения установленной разницы между ОДКС здоровых и поражённых пепельной гнилью растений были проведены более детальные исследования ОДКС на верхнем и нижнем (прикорневом) ярусах толерантной и восприимчивой к пепельной гнили сублиний.
Было установлено, что ОДКС нижнего и верхнего яруса растений толерантной сублинии в среднем составляло 652 и 672 кПа соответственно, при этом разница между ярусами была всего 3 %. ОДКС тканей прикорневого яруса восприимчивой к патогену сублинии составило 705 кПа, что было на 8 % выше аналогичного показателя у устойчивой изолинии. ОДКС тканей верхнего яруса восприимчивой сублинии достигло 792 кПа, превысив показатель ОДКС нижнего яруса этой же сублинии на 12,3 %.
В 2008–2009 гг. на растениях толерантной и восприимчивой к пепельной гнили сублиний сорта Парма было проведено определение вредоносности пепельной гнили у сои по элементам структуры урожая (табл. 4).
Таблица 4
Элементы структуры урожая растений толерантной и восприимчивой к пепельной гнили сублиний сорта Парма
ВНИИМК, 2008–2009 гг.
|
Вариант |
Количество с одного растения, шт. |
Масса 1000 семян, г |
||||
|
бобов |
семян |
|||||
|
2008 г. |
2009 г. |
2008 г. |
2009 г. |
2008 г. |
2009 г. |
|
|
Парма-T (толерантная сублиния) |
94,9± 8,5 |
90,1± 9,6 |
171,7± 15,1 |
204,5± 20,7 |
161,9± 10,2 |
174,2± 15,3 |
|
Парма-S (восприим чивая сублиния) |
73,4± 6,8 |
59,8± 8,6 |
149,5± 14,6 |
141,8± 23,0 |
134,4± 23,1 |
181,4± 25,5 |
Было установлено, что у растений Парма-S восприимчивой сублинии количество бобов и семян по сравнению с растениями толерантной сублинии Парма-Т было меньше. Уменьшение массы 1000 семян у растений восприимчивой сублинии Парма-S отмечено только в более засушливом 2008 г. В 2009 г. различия по этому показателю были в пределах ошибки.
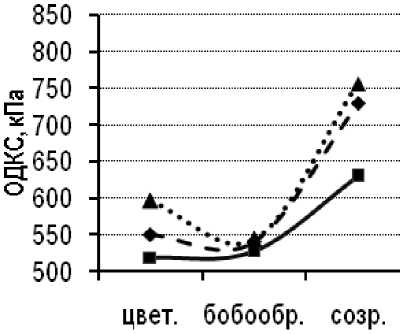
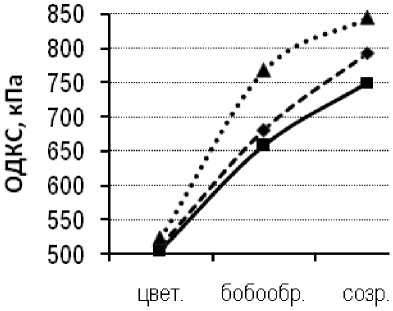
Сублинии сорта Парма с разной степенью устойчивости к пепельной гнили испытывали в 2009–2010 гг. В оба года исследований минимальный уровень ОДКС от цветения до физиологического созревания был зафиксирован у растений толерантной сублинии Парма-Т, соответственно максимальный уровень ОДКС – у восприимчивой сублинии Парма-S (рис. 5).
2009 г.
—*-- Парма (исходный сорт)
—■— Парма-Т (толерантная сублиния)
.....-К-— Парма-S(восприимчиваясублиния)
2010 г.
—*— Парма (исходный сорт)
—■— Парма-Т(толерантная сублиния)
.....А - Парма-S(восприимчиваясублиния)
Рисунок 5 – Онтогенетическая динамика ОДКС толерантной и восприимчивой к пепельной гнили сублиний и исходного сорта Парма, 2009–2010 гг.
Учёт болезни также показал, что развитие пепельной гнили на растениях толерантной сублинии Парта-Т начиналось на более позднем этапе и в целом было ниже по сравнению с исходным сортом и восприимчивой сублинией (табл. 5).
Таблица 5
Урожайность сублиний сорта Парма с различной устойчивостьюк пепельной гнили, т/га
ВНИИМК, 2009–2010 гг.
|
Вариант |
Развитие болезни на дату учёта, % |
Урожайность, т/га |
||||
|
2009 г. |
2010 г. |
|||||
|
налив семян (18.09) |
полная спелость (28.09) |
налив семян (8.09) |
полная спелость (20.09) |
2009 г. |
2010 г. |
|
|
Парма (исходный сорт) |
10,8 |
31,2 |
5,3 |
16,0 |
3,19 |
1,88 |
|
Парма-Т (толерантная сублиния) |
6,9 |
29,7 |
2,6 |
15,4 |
3,30 |
1,93 |
|
Парма-S (восприимчивая сублиния) |
14,1 |
34,8 |
8,8 |
15,7 |
3,10 |
1,81 |
|
НСР 05 |
- |
- |
- |
- |
0,14 |
0,09 |
Кроме этого, сублиния Парма-Т в оба года исследований обеспечивала достоверные прибавки урожая семян по сравнению с восприимчивой к болезни изолинии.
Таким образом, полученные экспериментальные результаты позволяют рекомендовать отбор сортообразцов на пониженный уровень ОДКС в критические по водопотреблению фазы развития растений при создании исходного материала для селекции сортов сои, толерантных к пепельной гнили.
Выводы. Микросклероции возбудителя пепельной гнили могут формироваться не только из сплетения нескольких гиф, но и интеркалярно или терминально из отдельных гиф, находящихся в сосудах, что повышает риск трахеомикозного увядания растений сои.
Вредоносность пепельной гнили проявляется в условиях водного стресса в период засухи и вызванного дефицитом воды повышения концентрации клеточ- ного сока в тканях растения-хозяина. При этом в тканях растений сои массово образуются микросклероции, полностью закрывающие просветы сосудов проводящей системы сои.
Рост гриба прекращается при достижении изотонического уровня осмотического давления питательной среды 930 кПа. Дальнейшее увеличение осмотического давления питательной среды является гипертоническим для гриба, что способствует массовому образованию микро-склероциев.
Разработан метод отбора толерантного к болезни исходного материала для селекции сои. Метод предусматривает выделение форм растений с осмотическим давлением клеточного сока, не превышающим 930 кПа в критические по водопо-треблению фазы бобообразования и налива семян. Гипертонические (более 930 кПа) значения ОДКС тканей сои приводят к массовому формированию микро-склероциев гриба.
Выделен исходный материал с пониженным уровнем ОДКС для создания толерантных к пепельной гнили сортов сои: Парма-Т, Вилана, Альба, Славия, Фора, КСХИ-709, Т-102.
