Использование принципа фоновых признаков при разработке критериев отличимости, однородности и стабильности для описания селекционных достижений

Автор: Дьяков А.Б., Гронин В.В.
Статья в выпуске: 2 (135), 2006 года.
Бесплатный доступ
Описаны различия фоновых признаков, предназначенных для идентификации генотипов отдельных растений по их фенотипам и для оценки селекционной ценности растений при отборах из популяций. Показано, что фоновые признаки с нулевой генотипической дисперсией не пригодны для селекционной идентификации генотипов, но могут эффективно обеспечивать отличимость растений инбредных линий, гибридов, сортов с целью их патентной защиты. Установлено, что метод индексов не уступает по эффективности методу фоновых признаков только при условии, если соотношение двух признаков описывается прямой линией, проходящей через начало координат.
Короткий адрес: https://sciup.org/142150775
IDR: 142150775 | УДК: 633.854.78:517:631.52
Use of background traits in elaboration of distinctness, uniformity and stability criteria for breeding achievements description
«OIL CROPS» Scientific bulletin, VNIIMK, 2006, is 2 (135). Differences between background traits for individual plant genotype identification by their phenotypes and for plant breeding value estimation in selection from populations are described. It is shown that background traits with zero genotypic variation are not suitable for breeding genotypes identification but may effectively ensure distinctness of inbred lines, hybrids and varieties plants for their patent protection. It is determined that index method efficiency may be equal to background traits method efficiency only in condition when relation of two characters is described by straight line equitation with zero free term.
Текст научной статьи Использование принципа фоновых признаков при разработке критериев отличимости, однородности и стабильности для описания селекционных достижений
Обеспечение охраноспособности селекционных достижений, поддержание сортовой чистоты и идентичности требуют разработки для каждого сорта, гибрида, родительской линии критериев отличимости, однородности и стабильности. Базирующееся на таких критериях описание может быть эффективным лишь при условии, что оно обеспечивает надежное опознание (идентификацию) каждого растения конкретного сорта вследствие его отличия по комплексу морфологических признаков от растений других сортов (гибридов, линий) данной культуры. В странах Западной Европы определены центры, в которых производятся испытания селекционных достижений на отличимость, однородность и стабильность (Гончаров, 1996). В России принятый 16.08.93 г. закон «О селекционных достижениях» также требует проведения таких испытаний. Оценка отличимости, однородности и стабильности (ООС) селекционных достижений стала особенно актуальной после 24.04.98 г., когда Российская Федерация стала членом Международного Союза UPOV. В стране выделено 38 госсортучастков и госсортстанций, проводящих оценку на ООС селекционных достижений, поступивших не только на выдачу патента, но и на допуск к использованию. Очень сложна также проблема идентификации образцов коллекций генетических ресурсов, насчитывающих тысячи номеров (Porter, Smith, 1982).
Для того чтобы идентификация сортов была точной, свободной от возможных ошибок, она должна быть основана на комплексе признаков (Baum, 1981). Используемые при этом морфологические признаки необходимо изучить на стабильность фенотипического проявления в разных условиях внешней среды (Smith, Smith, 1989). В результате таких испытаний установлено, что даже часть морфологических признаков разных культур, которые в системе UPOV чаще всего используются в качестве апробационных, характеризуются высокой вариабельностью в зависимости от условий внешней среды (Жученко, 2001, с. 895). Значительная средовая изменчивость ряда признаков подсолнечника, используемых в определителе UPOV, обнаружена и нами (Гронин, 2003). Поэтому важно разработать способы обеспечения достаточно точной отличимости растений разных генотипов по морфометрическим признакам, для величин которых характерна высокая средовая вариабельность.
На основе сопоставления разных способов элиминации средовой изменчивости количественных признаков с целью повышения надежности идентификации генотипов отдельных растений Э. Х. Гинзбург (1984, с. 33-38) заключил, что наиболее перспективны для этого методы, основанные на учете фоновых признаков. Наши исследования показали высокую эффективность использования таких признаков при разработке методик на отличимость, однородность и стабильность. В данной статье это демонстрируется на примере отличимости растений двух инбредных родительских линий гибридов подсолнечника ВК 499 и ВК 678, различающихся в среднем по включенному в определитель UPOV признаку “размер листьев”. Для оценки степени нестабильности показателей длины и ширины листьев и для усиления их модификационной изменчивости растения этих линий выращивали в разные по погодным условиям 2001 и 2002 годы при разной интенсивности конкуренции в посевах различной густоты – от 2 до 7 растений на 1 м2. В начале фазы бутонизации измеряли длину и ширину пластинки самого крупного листа каждого изучаемого растения. Усредненные за оба года по всем густотам данные измерений представлены в табл. 1.
Таблица 1 – Усредненные оценки длины и ширины (см) листовых пластинок и показатели их вариабельности у растений инбредных линий подсолнечника, выращенных в 2001 и 2002 гг. при густотах посевов от 2до 7 растений/м2
|
Промеры листовой пластинки |
Инбред-ная линия |
Число растений, шт. |
Дисперсия |
Стандартное отклонение |
Коэффициент вариации, % |
Средняя величина, см |
99%-ный доверительный интервал средней |
|
Длина |
ВК 499 |
103 |
18,75 |
4,33 |
22,6 |
19,15 |
±1,10 |
|
Длина |
ВК 678 |
109 |
6,58 |
2,57 |
20,2 |
12,73 |
±0,63 |
|
Ширина |
ВК 499 |
103 |
26,82 |
5,18 |
27,7 |
18,70 |
±1,31 |
|
Ширина |
ВК 678 |
109 |
13,30 |
3,65 |
24,1 |
15,12 |
±0,90 |
Анализ полученных данных показал, что в среднем у растений линии ВК 499 размеры листьев больше, чем у линии ВК 678 как по длине, так и по ширине. Оказалось также, что у линии ВК 499 оценки длины и ширины пластины листьев отличаются более высокими показателями не только абсолютной, но и относительной вариабельности по сравнению с линией ВК 678. Обнаружилась также тенденция меньшей изменчивости длины листьев, чем их ширины. Поэтому доверительные интервалы средних величин для длины пластинок листьев оказались более узкими, чем для их ширины. А поскольку изучаемые линии различаются в большей степени также по длине пластинок листьев, их идентификация по этому показателю более надежна (19,15-1,10>12,73+0,63), чем по ширине листьев (18,70-1,31>15,12+0,90). Однако и по усредненным из 100 измерений оценкам ширины листьев различимость этих двух линий также достаточно надежна, поскольку приведенные в табл. 1 двусторонние доверительные интервалы вычислены для уровня вероятности 99%, а при парных сравнениях применим односторонний критерий, поэтому в данном случае линии ВК 499 и ВК 678 различаются по усредненным оценкам ширины листьев с вероятностью ошибки меньшей, чем 0,5%.
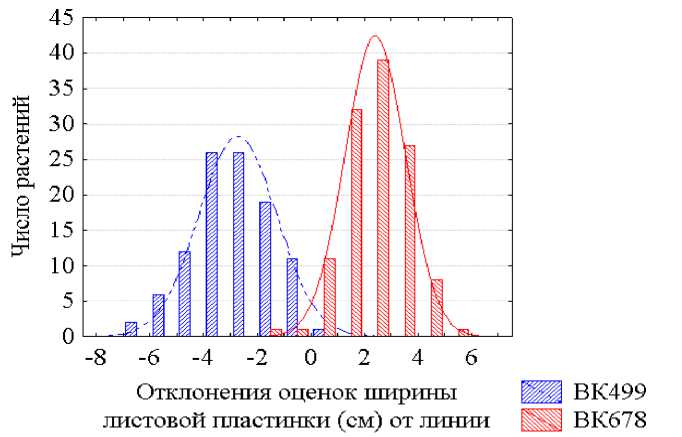
Более детальным анализом полученных данных установлено, что при сравнении растений, выращенных при их равномерном размещении и одинаковой площади питания в посеве, линии ВК 499 и ВК 678 достаточно надежно различаются по средним оценкам из 20 измерений длины листовых пластинок, но для их отличимости по ширине листьев такой численности выборок недостаточно и при таких условиях. По характеристикам же отдельных растений эти линии неотличимы ни по длине, ни по ширине листа вследствие большой модификационной изменчивости величин этих признаков и значительных трансгрессий распределений численностей растений линий ВК 499 и ВК 678 как по оценкам длины листовых пластинок (рис. 1), так и, особенно, их ширины (рис. 2.). Такие захождения одного распределения в пределы другого свидетельствуют о невозможности идентификации растений этих линий по таким размерам листьев. В. Н. Исаков с соавторами (1982) также пришли к выводу о непригодности оценок длины и ширины листьев для распознавания принадлежности растения даже к тому или иному виду или разновидности. Они установили, что при сравнении шести пар видов кленов коэффициенты дискриминации варьировали лишь от K =0,61 до K =1,28 для длины листовых пластинок и от K =0,00 до K =2,42 для их ширины. Н. И. Вавилов (1966, с. 364) также писал о крайней ненадежности использования в качестве критерия классификации таких сильно варьирующих признаков, как длина листа.
Вследствие широкой модификационной изменчивости, показанной на графиках (рис. 1 и 2), использовать признаки длина и ширина листьев при разработке методик на отличимость, однородность и стабильность инбредных линий подсолнечника можно было лишь при условии коррекции оценок этих признаков с целью элиминации ненаследственной их вариации. Поскольку для обеспечения такой коррекции разработано два варианта принципов использования фоновых признаков, надо было вначале выбрать наиболее приемлемый для данного случая вариант.
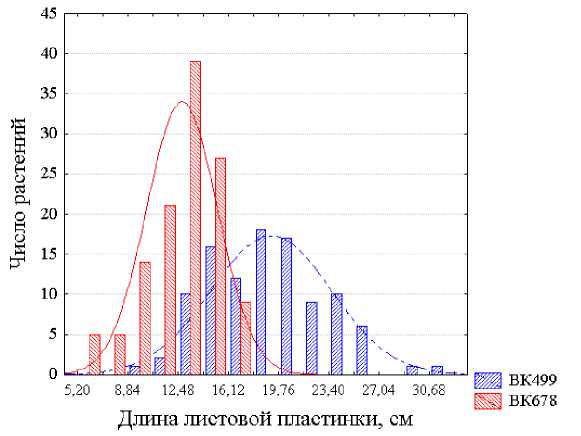
Рисунок 1 – Распределения растений линий подсолнечника ВК 499 и ВК 678 по оценкам признака длина листовой пластинки
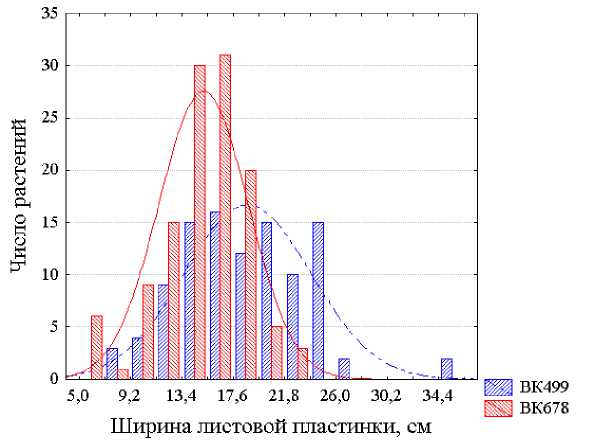
Рисунок 2 – Распределения растений линий подсолнечника ВК 499 и ВК 678 по оценкам признака ширина листовой пластинки
Разрабатывая принцип фоновых признаков, В. А. Драгавцев (1963, 1966, 1974) изначально ставил задачу найти способ разграничения генотипической и экологической изменчивости количественных признаков растений, позволяющий на этой основе оценивать генотип каждой особи популяции по ее фенотипическим характеристикам. В этих исследованиях был использован подход, аналогичный принципу контрольных делянок в полевых опытах по испытанию урожайности генотипов. Поскольку на всех таких делянках опыта высеваются растения одного генотипа, получаемые на них оценки урожайности характеризуют экологическую изменчивость, а наличие экологической корреляции между показателями соседних делянок позволяет расчленять дисперсию оценок урожайности испытуемых генотипов, выделив из нее обусловленный пестротой плодородия почвы компонент, и на этой основе откорректировать оценки урожайности отдельных генотипов.
Изучая изменчивость растений в популяциях, В. А. Драгавцев (1961) пришел к выводу, что о наследственных различиях особей можно судить не только по устойчивым к внешним воздействиям признакам, но и по таким вариабельным признакам, величины которых обнаруживают устойчивые сдвиги относительно других признаков, в такой же степени зависимых от условий внешней среды. В соответствии с этим для регистрации такого устойчивого сдвига признака В. А. Драгавцев (1963) на следующем этапе исследований предложил только один критерий выбора фонового признака - наличие экологической корреляции между ним и селекционным признаком. Он писал: «Единственное требование, предъявляемое к фону сравнения - это необходимость связи фона с оцениваемым признаком. Существует неограниченное количество фонов сравнения для оценки сдвигов подвижных признаков. Фоном сравнения может служить любой показатель, связанный с оцениваемым» (Драгавцев, 1963). Требование к фоновым признакам, которые автор назвал еще директивными, он сформулировал позже несколько иначе: «Лучшим фоном сравнения будет признак, в сильной степени зависящий от условий среды и в малой - от специфики расы... Эти признаки, названные нами директивными, легко обнаруживаются в популяции при изучении контрастных пар особей. Если значения какого-либо признака (коррелирующего с оцениваемым) у обеих особей контрастной пары близки, то он вполне пригоден для использования в качестве фона сравнения» (Драгав-цев, 1966). В результате дальнейших исследований В. А. Драгавцев (1969) установил, что в случаях, когда вариация фонового признака обусловлена не только изменчивостью условий внешней среды, но и наследственными различиями растений, выравнивание оценок селекционного признака по величинам фонового элиминирует не только средовую, но и часть генотипической компоненты вариации, причем эта часть будет пропорциональна генотипической дисперсии фонового признака. Это потребовало сформулировать более жесткие требования к фоновому признаку: в идеале он не должен иметь наследственной изменчивости, а коэффициент его средовой корреляции с селекционным признаком должен быть близким к единице.
Введение дополнительного требования к признакам, пригодным для использования в качестве фоновых, резко осложнило поиски таких признаков (Драгавцев, 1972; Гинзбург, 1984). Если по критерию наличия высокой средовой корреляции с селекционным признаком легко выявлялось много фоновых признаков (Драгавцев, 1963, 1966), то найти среди них признаки с нулевой или близкой к нулю наследственной изменчивостью оказалось очень трудно. По мнению Н. В. Тимофеева-Ресовского и В. И. Иванова (1966), а также К. Мазера и Дж. Джинкса (1985, с. 55) ненаследственных признаков вообще не существует. Даже у показателей водного режима тканей растений, которые были пригодны в качестве фоновых при изучении клонированных растений тополя (Ничипоренко, 1969), была обнаружена значительная собственная генотипическая дисперсия в исследованиях с гетерогенными популяциями пшениц (Герасименко и др., 1979). Детальный анализ структуры изменчивости признаков растений показал, что в полевых посевах любой пригодный для практического использования фоновый признак имеет собственную генотипическую дисперсию, обусловленную разной конкурентоспособностью растений (Дьяков, Драгавцев, Бехтер, 1976).
Изучение высоко- и низкомасличных популяций подсолнечника А. Б. Дьяков проводил с целью физиолого-генетического анализа достижений и методов работы академика В. С. Пустовойта. При этом с самого начала было ясно, что способ расчленения фенотипических дисперсий оценок селекционных признаков на генетический и экологический компоненты не может обеспечить селекционную идентификацию желательных генотипов. Во-первых, значительная изменчивость урожайности и масличности семян подсолнечника за счет различий генотипов по длительности вегетации так же бесполезна для селекционера, как и экологическая вариация. Во-вторых, было показано (Прокофьев, Дьяков, 1961), что на изменчивость этих и других признаков очень большое влияние оказывает конкуренция в посевах, причем эффекты разной конкурентоспособности растений тоже селекционно бесполезны, даже если они наследуются. Наряду с этими компонентами вариации при испытаниях созданных В. С. Пустовойтом высокомасличных сортов подсолнечника выявляется их значительное превосходство над старыми низкомасличными сортами, не только по масличности, но и по урожаю семян («ядер» семянок) при равной длительности вегетации (Дьяков, 1964, 1965). Важно было изучить физиолого-генетическую природу этих различий между сортами, так как они являются проявлением селекционно значимого компонента изменчивости этих признаков. Поэтому способ идентификации элитных растений в популяциях должен выявлять генотипы с высокой степенью выраженности такого типа отклонений, несмотря на фенотипическое проявление суммы экологических и генотипических компонент селекционно бесполезной изменчивости.
На основе этого исходного анализа поставленной задачи был разработан принципиально новый физиолого-генетический подход к расчленению экологической и генотипической дисперсий количественных признаков растений. С первых этапов работы было начато изучение эффектов двух компонент экологической дисперсии σ2 e количественных признаков подсолнечника: изменчивости, обусловленной вариацией плодородия почвы σ2 eπ , а также ненаследуемой конкурентной изменчивостью σ2 ecom (Прокофьев, Дьяков, 1961; Дьяков, 1964). Изучение наследственной изменчивости того же комплекса признаков планировалось на основе представления о том, что в генотипическую дисперсию включены компоненты, обусловленные разной длительностью вегетации генотипов σ2 gd , их различий по конкурентоспособности в посевах σ2 gcom , а также селекционно полезная изменчивость σ2 gu , причиной которой являются различия генотипов по эффективности использования растениями ресурсов внешней среды. Объектами изучения физиолого-генетической природы селекционно полезной изменчивости σ2 gu служили как разные по уровню маслонакопления сорта-популяции подсолнечника, так и отдельные растения этих популяций. Для исключения эффектов σ2 gd сравнивались пары сортов с равной скороспелостью, а проявления эффектов σ2 eπ , σ2 ecom и σ2 gcom сводили к минимуму или устраняли в вегетационных опытах выращиванием при равных условиях по одному растению в сосуде, а в полевых – использованием защитных рядов и способами коррекции данных. При изучении эффектов σ2 gd сравнивались сорта разной длительности вегетации, эффекты σ2 eπ моделировались внесением удобрений, эффекты σ2 ecom – всеванием конкурентов в разные сроки, проявления σ2 gcom – смешанными посевами разных генотипов. Важной деталью всех этих исследований было то, что впервые изучалась изменчивость урожаев не только плодов (семянок), но и собственно семян («ядер» семянок).
С целью выяснения физиолого-генетической природы селекционно полезной компоненты ( σ2 gu ) изменчивости растений подсолнечника вначале были проверены предложенные разными авторами гипотезы для объяснения причин наследственных различий по уровню жиронакопления в семенах. В результате было установлено, что у растений высокомасличных сортов по сравнению с низкомасличными площадь листьев и продолжительность их работы даже уменьшены, но ассимилятов образуется больше, чем могут использовать на синтез жира семена, поэтому различия по интенсивности фотосинтеза являются не причиной, а следствием разного уровня маслообразования (Прокофьев, Дьяков, 1961; Дьяков, 1966в). Была показана ошибочность гипотезы «микрораспределений пластики» (Дьяков, 1966б), согласно которой урожаи семян и масла высокомасличных сортов повышены по причине уменьшения расходования ресурсов на построение плодовых оболочек семянок. Разными экспериментами была опровергнута также теория антагонизма между процессами биосинтезов жира и белка (Дьяков, 1964). Сторонники этой теории считают, что маслонакопление может усиливаться лишь на столько, насколько сократится биосинтез белка. Показано также (Дьяков, 1966а), что увеличение масличности семян в процессе селекции подсолнечника обусловлено не уменьшением поглощения азота, а повышением эффективности его использования.
Проведенными исследованиями было установлено (Дьяков, 1964), что количество масла, накапливаемого в семенах подсолнечника за период налива, предопределяется до начала биосинтеза запасного жира, в период эмбрионального роста семян. Оказалось, что за период их налива синтезируется тем больше масла, чем больше азота поступит в растущие семена, и, соответственно, больше будет в них конституционных веществ, а азот, поступающий в наливающиеся семена, включается в запасные белки, не влияющие на синтез масла, но снижающие масличность семян (Дьяков, 1964). Соцветия высокомасличных растений подсолнечника перед цветением и в период роста семян оказывают более сильное аттрагирующее воздействие на вегетативные органы, подавляя их рост и усиливая отток пластических веществ (Дьяков, 1965). Особенно важно то, что вследствие генотипических различий по аттрагирующей способности соцветий в это время в семена высокомасличных растений поступает больше азота в период роста за- родышей и меньше – в период налива семян (Дьяков, 1964, 1966в). Хотя оказалось, что высоко- и низкомасличные генотипы в обычных условиях не различаются по поглощению азота из почвы (Дьяков, 1966а), разная динамика притока азота в семена является причиной различий по урожаю семян и масла (Дьяков, 1964, 1966в). Прямыми опытами установлено, что эти различия при равной длительности вегетации генотипов обусловлены только одной причиной – разной аттрагирующей способностью соцветий в период эмбрионального роста семян (Дьяков, 1964, 1966б). Это позволило считать, что селекционно полезный компонент σ2gu генотипической дисперсии уровней накопления в семенах обусловлен в популяциях подсолнечника изменчивостью генотипов по аттрагирующей способности соцветий, и его можно обозначить символом σ2gattr. Установлено, что этот компонент дисперсии является причиной вариации признака при соблюдении равенства длительности вегетации, условий внешней среды и конкурентного стресса (Дьяков, 1965).
В соответствии с полученными результатами эффективность идентификации желательных генотипов определяется тем, насколько надежно можно выявить элитные растения с максимальными наследственными отклонениями по аттрагирующей способности соцветий в период роста семян на фоне ряда селекционно бесполезных компонент экологической и генотипической изменчивости. Поэтому важно было установить комплекс скоррелированных признаков, фенотипическое проявление которых обусловлено генотипическими различиями по аттрагирующей способности, а также характер вариации и ковариации этих признаков под воздействием разных причин селекционно бесполезной изменчивости.
Полевыми и вегетационными опытами установлено, что вследствие усиления аттракции азота растущими семенами подсолнечника увеличивается масса запасающей жир ткани, возрастает доля конституционных белков в общей массе белка в семенах, вследствие чего повышается эффективность использования поглощенного азота (Дьяков, 1964, 1966б). Поэтому такие наследственные различия по аттрагирующей способности являются причиной высоких генотипических корреляций урожаев семян («ядер» семянок) с процентным содержанием белка в них до r g =-0,88, а также с масличностью семян до r g =0,91 вследствие отрицательной связи между процентами жира и белка до r g =-0,90 (Дьяков, 1964, 1965, 1966а). Еще более высокими оказались корреляции урожаев масла с масличностью семян, обычно около r g =0,94, а с урожаями семян связь даже почти функциональная, обычно около r g =0,99 как в вегетационных, так и в полевых опытах (Дьяков, 1965, 1966а, 1967). У генотипов с усиленной аттракцией интенсивнее уменьшается сухая масса вегетативных органов и цветоложа корзинки во второй половине вегетации, что обусловливает отрицательную генотипическую связь между урожаями семян и массой вегетативных органов (Прокофьев, Дьяков, 1961; Дьяков, 1965).
Иной характер зависимостей между теми же признаками обнаруживается в тех случаях, когда их ковариация была обусловлена разным плодородием почвы или конкуренцией между растениями. С повышением урожая семян под влиянием этих причин увеличиваются размеры и масса вегетативных органов, а масличность семян снижается (Прокофьев, Дьяков, 1961; Дьяков, 1964, 1966в). По данным опыта, в котором изучалась реакция растений одного сорта подсолнечника на разные уровни содержания азота в почве, были обнаружены высокие отрицательные экологические корреляции величин урожаев семян и их масличности r e =-0,961, а также урожаев масла и масличности семян r e =-0,908 (Дьяков, 1967). Также отрицательны экологические ковариации этих признаков при обусловленности их вариации конкуренцией между растениями в посевах (Прокофьев, Дьяков, 1961; Дьяков, 1967).
Было установлено, что при отборах только по урожаю семян из популяций выделяются в основном растения, причинами высокой продуктивности которых являются повышенное плодородие почвы и, в еще большей мере, освоение увеличенной площади питания в посеве вследствие наследственного или ненаследуемого конкурентного преимущества. Для устранения таких ошибок идентификации желательных генотипов разработан оригинальный вариант принципа фоновых признаков, основанный на явлении разнонаправленности ковариаций селекционного и фонового признаков, обусловленных селекционно полезной их изменчивостью и суммой бесполезных для селекционера компонент не только экологической, но и генотипической дисперсий (Дьяков, 1967). В идеа-43
ле на графике зависимости величин селекционного признака (СП) от значений фонового признака (ФП) точки особей популяции должны смещаться вдоль экологической линии регрессии в той мере, в какой они отражают сумму эффектов компонент селекционно бесполезной вариации. В то же время противоположная направленность селекционно полезной ковариации СП и ФП обусловливает отклонения точек от экологической регрессии на этом графике. Поэтому степень отклонения точки в нужном направлении от такой регрессии служит оценкой селекционной ценности соответствующего элитного растения.
Сопоставление описанных двух вариантов принципа фоновых признаков позволило выявить не только различия между ними (Дьяков, Драгавцев, 1975; Дьяков, Дра-гавцев, Бехтер, 1976; Драгавцев, Дьяков, 1982). Был сделан вывод, что у растений выращенной в сомкнутом агроценозе популяции не может быть найден фоновый признак, причиной варьирования которого была бы только пестрота плодородия почвы, и он не был бы чувствителен к проявлениям генотипических различий по конкурентоспособности. Поэтому реальные фоновые признаки не могут обеспечить идеальную идентификацию генотипов особи гетерогенной популяции, но позволяют вычислять, какими показателями обладало бы каждое конкретное растение, если бы не только уровень плодородия почвы, но и величина использованной площади питания в посеве не отличались от средних уровней для всей популяции. В отличие от такой генотипической идентификации предложенный А. Б. Дьяковым подход к селекционной идентификации желательных генотипов получил название принципа ортогональности селекционно полезной и бесполезных компонент ковариаций СП и ФП. При этом в идеале в качестве фонового подбирается признак, чувствительный к n факторам, сдвигающим точки особей вдоль экологической линии регрессии, а в качестве селекционного – признак, к изменчивости которого, обусловленной эффектами тех же n факторов, добавлена селекционно полезная вариация, приводящая к отклонениям точек на графике от экологической линии регрессии. О принципиальной возможности такого вычленения одного компонента вариации свидетельствуют также данные Ю. А. Филипченко (1979), который обнаружил, что соотношение таких полигенных признаков, как ширина колосковой чешуи и ее длина, контролируется только парой аллелей одного гена.
Методы селекционной идентификации, основанные на принципе ортогональности компонент ковариаций СП и ФП, позволили достаточно эффективно выделять лучшие по уровню маслообразования генотипы среди растений популяций подсолнечника, выращенных в полевых посевах даже при увеличенных различиях по плодородию почвы и конкуренции (Дьяков, 1967, 1969; Дьяков, Драгавцев, 1975;Дьяков, Драгавцев, Бехтер, 1976). Несмотря на проблемы поиска идеального фонового признака с σ2 g =0, в ряде опытов Драгавцева (1963, 1966, 1969) достигалась эффективная идентификация генотипов по отклонениям точек от линий экологических регрессий СП на ФП. Общей особенностью этих опытов было то, что сравнивались растения клонов двух генотипов тополя (Драгавцев, 1963, 1969), клонов двух типов подвоев яблони дусенов или двух сортов сливы, особи которых также являются клонами (Драгавцев, 1966). Поскольку в пределах клона практически отсутствует наследственная изменчивость, в том числе и по конкурентоспособности, в таких опытах соблюдается требование нулевой генотипической дисперсии у идеального ФП, что и явилось причиной эффективности генетической идентификации в таких опытах.
Объектами исследований по разработке критериев отличимости, однородности и стабильности также являются инбредные линии, первые поколения межлинейных гибридов или чистолинейные сорта культур-самоопылителей. В пределах каждого из таких объектов при соблюдении критерия их однородности растения должны быть генетически идентичными, поэтому у них можно легко найти фоновые признаки с нулевой генотипической дисперсией. Это дает основание считать, что разработанный В. А. Драгавцевым вариант принципа фоновых признаков наиболее пригоден для элиминации экологической изменчивости при использовании количественных признаков растений в качестве критериев отличимости, однородности и стабильности. Более того, для этой цели непригоден принцип ортогональности разных компонент ковариаций СП и ФП, специально разработанный для селекционной идентификации желательных генотипов при отборах из гетерогенных и гетерозиготных популяций.
Вторым требованием к идеальному фоновому признаку по В. А. Драгавцеву (1974) является наличие высокой экологической корреляции ФП с СП. В нашем примере коэффициенты таких корреляций между длиной и шириной листовой пластинки оказались очень высокими для растений как инбредной линии подсолнечника ВК 499 r e =0,97, так и для особей линии ВК 678 r e =0,95. Поскольку в среднем по длине листовых пластинок линия ВК 499 высокозначимо превосходит линию ВК 678 (см. табл. 1), соответствующие экологические регрессии должны различным образом располагаться на графике. Действительно, регрессия длины листовых пластинок на их ширину растений инбредной линии ВК 499 лежит выше такой же экологической регрессии растений линии ВК 678 (рис. 3).
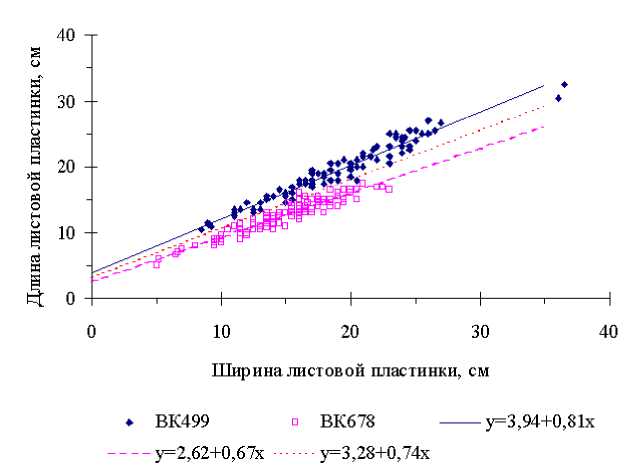
Рисунок 3 – Экологические регрессии величин длины листовой пластинки (СП) на ее ширину (ФП) растений линий подсолнечника ВК 499 и ВК 678
Обусловлено это различиями по значениям как свободных членов уравнений a , так и коэффициентов регрессии b . Построенная по среднеарифметическим значениям параметров a и b двух линий регрессии, третья линия оптимальным образом разделяет на графике точки, соответствующие характеристикам растений генотипов ВК 499 и ВК 678. Величины отклонений этих точек от такой линии практически независимы от пестроты плодородия почвы и средовых различий по конкурентоспособности растений потому, что эффекты этих компонент вариации элиминируются экологическими регрессиями величин одного признака на другой. В результате в вариацию величин отклонений включены в основном лишь эффекты погрешностей измерений. Поэтому значительно уменьшились дисперсии оценок этих признаков, например, для длины пластинок листьев линии ВК 499 в 16 раз – с 18,75 до 1,16, а для линии ВК 678 в 9,4 раза – с 6,58 до 0,70.
Судя по разбросу точек на графике (рис. 3), использование таких признаковых координат позволяет значительно повысить отличимость растений разных генотипов по количественным морфологическим признакам. Это наглядно иллюстрируется графиками распределения растений инбредных линий подсолнечника ВК 499 и ВК 678 по значениям отклонений от усредненной линии экологической регрессии СП на ФП величин как длины листовой пластинки (рис. 4), так и ее ширины (рис. 5). В обоих случаях имеют место только небольшие перекрытия площадей распределения частот растений двух сравниваемых генотипов. Сравнение графиков распределений растений двух генотипов по откорректированным оценкам длины и ширины листовых пластинок (рис. 4 и 5) с графиками распределений тех же растений по неоткорректированным величинам тех же признаков (см. рис. 1 и 2) свидетельствуют о том, что методика идеальной идентификации генотипического значения признака у любого, отдельно взятого растения с использова- нием фонового признака с нулевой генотипической дисперсией позволяет за счет резкого уменьшения трансгрессий признаков многократно снизить вероятность ошибок опознания (идентификации) каждого растения сравниваемых по количественным признакам генотипов.
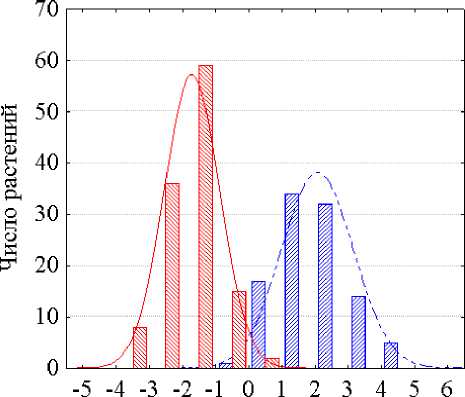
Отклонения оценок длины листовой пластинки (см) от линии регрессии
ВК499
ВК678
Рисунок 4 – Распределения растений линий подсолнечника ВК 499 и ВК 678 по величинам отклонений длины листовой пластинки от усредненной линии экологической регрессии СП на ФП
регрессии
Рисунок 5 – Распределения растений линий подсолнечника ВК 499 и ВК 678 по величинам отклонений ширины листовой пластинки от усредненной линии экологической регрессии СП на ФП
Ю. А. Филипченко (1979) такую задачу предлагает решать путем вычисления индексов, то есть отношений оценок одного количественного признака к величинам другого. В случае нашего примера отношение длины листовой пластинки к ее ширине является индексом формы листа. Судя по степени перекрытия площадей распределений растений сравниваемых линий по величинам такого индекса формы листа (рис. 6), надежность идентификации особей этих генотипов с использованием этого индекса выше, чем по абсолютным величинам длины и ширины листовых пластинок (см. рис. 1 и 2), но менее надежна, чем их распознание по отклонениям тех же величин от экологической линии регрессии СП на ФП (см. рис. 4 и 5).
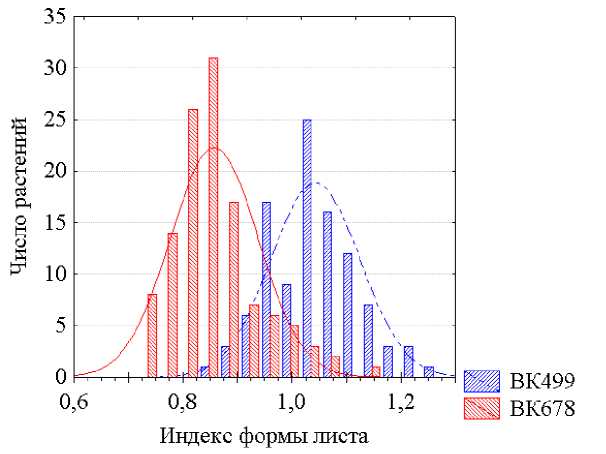
Рисунок 6 – Распределения растений линий подсолнечника ВК 499 и ВК 678 по величинам индекса формы листа (отношения длины листовой пластинки к ее ширине).
Эффективность использования трех способов идентификации отдельных растений двух генотипов подсолнечника оценили также путем вычисления значений коэффициента дискриминации, предложенного А. А. Любищевым (1959). Этот коэффициент является отношением квадрата разности средних величин признака двух генотипов к сумме дисперсий того же признака тех же выборок особей этих генотипов. По значениям этого коэффициента вычисляются показатели вероятности ошибок идентификации генотипов ( p ) и числа оцененных растений в расчете на одно ошибочное определение (О). Результаты вычислений этих показателей представлены в табл. 2.
Таблица 2 – Величины коэффициентов дискриминации (K), вероятности ошибочных определений (p) и числа оцененных особей на одну ошибку (О) при идентификации растений линий подсолнечника ВК 499 и ВК 678 по признакам ширина и длина листовых пластинок, их соотношению (индексу формы) и откорректированным значениям с использованием регрессии СП на ФП
|
Способ идентификации |
Признак листовой пластинки |
K |
p |
О |
|
Без коррекции |
ширина |
0,3 |
0,87 |
1:2,7 |
|
Без коррекции |
длина |
1,6 |
0,15 |
1:6 |
|
По индексу формы |
длина / ширина |
2,6 |
0,13 |
1:8 |
|
С коррекцией по ФП |
ширина |
4,8 |
0,06 |
1:18 |
|
С коррекцией по ФП |
длина |
7,7 |
0,02 |
1:44 |
Судя по данным табл. 2, для идентификации генотипов отдельных растений линий подсолнечника ВК 499 и ВК 678 более пригоден признак длина листовой пластинки, чем ее ширина, поэтому в качестве СП лучше использовать оценки длины, а более чувствительные к пестроте среды величины ширины могут служить в качестве ФП. Однако при идентификации по неоткорректированным оценкам даже длины листа остается высокой вероятность ошибочных определений. Оценки по отношению длины листо- вой пластинки к ее ширине, то есть по индексу формы листа, в некоторой степени повышают отличимость отдельных растений тех же генотипов, но надежность идентификации повышается лишь в небольшой степени и остается слишком низкой. Элиминация экологической изменчивости на основе использования фонового признака в 16 раз увеличивает коэффициент дискриминации при идентификации по ширине листовой пластинки и в 4,8 раза – по ее длине. Самой эффективной оказалась идентификация сравниваемых в опыте линий подсолнечника по значениям отклонений оценок длины листовых пластинок от экологической линии регрессии СП на ФП. Хотя коэффициент дискриминации при этом не поднялся до величины K=18, обеспечивающей полную идентификацию, при K=7,7 вероятность ошибок меньше 2%.
Хотя Ю. А. Филипченко (1979) смог почти полностью элиминировать экологическую изменчивость показателей размеров колосковой чешуи пшеницы путем вычисления индекса ее формы как отношения ширины к длине, сам он предупреждает, что это очень редкий случай эффективного использования метода индексов. По его наблюдениям успех при этом может быть обеспечен при соблюдении такого эмпирически найденного правила: «Меньшую изменчивость индекса по сравнению с абсолютными величинами можно ожидать лишь в тех случаях, когда обе сравниваемые величины связаны довольно большой корреляцией и их изменчивость под влиянием внешних условий имеет приблизительно одинаковый характер». С позиций принципа фоновых признаков мы можем уточнить это правило: метод индексов может быть не менее эффективным, чем использование ФП, при условии, если экологическая регрессия одной величины на другую является прямой линией, проходящей через начало координат. В нашем случае неэффективность метода индексов объясняется тем, что экологическая вариабельность ФП (ширины листовой пластинки) больше, чем дисперсия СП (длины листовой пластинки), поэтому большие оси эллипсов рассеивания точек на графике зависимости СП от ФП проходят соответственно выше начала координат (см. рис. 3). Вследствие с уменьшением значений ФП возрастают величины отношения СП/ФП, то есть сохраняется значительная часть экологической изменчивости. В таких случаях использование принципа фоновых признаков безусловно обеспечивает более высокую эффективность идентификации генотипов, чем метод индексов.
