Исследование антигенотоксического действия криофракций селезенки и плаценты крупного рогатого скота в условиях действия циклофосфамида

Автор: Востроилова Г.А., Шабанов Д.И., Корчагина А.А., Хохлова Н.А., Некрасов А.В.
Журнал: Сельскохозяйственная биология @agrobiology
Рубрика: Тканевые препараты
Статья в выпуске: 2 т.61, 2026 года.
Бесплатный доступ
Сохранение стабильности генома сельскохозяйственных животных связано с целостностью ДНК их клеток в условиях экологического неблагополучия. В настоящей работе было впервые установлено антигенотоксическое влияние гидрофильной криофракции селезенки крупного рогатого скота (ГКСК) в печени и костном мозге, а также антиапоптотическое действие ГКСК, липофильной криофракции селезенки крупного рогатого скота (ЛКСК) и равной смеси липофильной криофракции селезенки и плаценты крупного рогатого скота (ЛКСПК) в костном мозге мышей в условиях индуцированной циклофосфамидом (ЦФ) генотоксичности. Целью работы было определение влияния гидрофильной криофракции селезенки, липофильной криофракций селезенки и смеси липофильной криофракции селезенки и плаценты крупного рогатого скота на генотоксическое действие циклофосфамида у мышей. Исследование проводили во Всероссийском научно-исследовательском ветеринарном институте патологии, фармакологии и терапии (ФГБНУ ВНИВИПФиТ) в 2025 году. Использовали фармацевтические субстанции, содержащие ГКСК, ЛКСК и ЛКСПК (ФГБНУ ВНИВИПФиТ). Экспериментальным генотоксикантом служил Эндоксан® («Baxter Oncology GmbH», Германия), содержащий в качестве действующего вещества циклофосфамид моногидрат (ЦФ). Были сформированы 11 групп лабораторных беспородных мышей-самцов (Mus albus officinarum, n = 126) с массой тела 26,0±2,0 г. В I группе (негативный контроль) животные получали однократную внутримышечную и внутрибрюшинную инъекцию изотонического раствора хлорида натрия в объеме соответственно 0,1 и 0,5 мл (n = 6). Мыши во II, III и IV группах получали однократную внутримышечную инъекцию соответственно ГКСК, ЛКСК и ЛКСПК в дозе 0,5 мл/кг и объеме 0,1 мл (n = 12 в каждой группе). Животные в V группе выступали в качестве позитивного контроля, им однократно внутрибрюшинно вводили ЦФ в дозе 40 мг/кг и объеме 0,5 мл (n = 12). Животным в VI группе однократно внутримышечно вводили ГКСК в дозе 0,5 мл/кг и объеме 0,1 мл перед внутрибрюшинной инъекцией ЦФ в дозе 40 мг/кг и объеме 0,5 мл (n = 12). Мышам из VII и VIII групп однократно вводили соответственно ЛКСК и ЛКСПК перед введением ЦФ в дозах, аналогичных таковым в VI группе (n = 12). Животным из IX, X и XI групп трехкратно с интервалом 24 ч вводили соответственно ГКСК, ЛКСК и ЛКСПК в дозе 0,5 мл/кг и объеме 0,1 мл (n = 12 в каждой группе). Совместно с последней инъекцией исследуемой субстанции мыши однократно внутрибрюшинно получали ЦФ в дозе 40 мг/кг и объеме 0,5 мл. Животных выводили из эксперимента посредством передозировки углекислого газа в специальной камере через 6 и 24 ч после введения ЦФ. Из бедренных костей получали образцы костного мозга, также отбирали образцы печени. Ткани гомогенизировали и использовали для проведения щелочного электрофореза единичных клеток (метод ДНК-комет). Определяли содержание ДНК в «хвосте кометы» (TDNA, %) через 6 и 24 ч после инъекции ЦФ. С помощью коммерческого набора Annexin V-IF488/PI Cell Apoptosis Detection Kit («Servicebio», Китай) методом флуоресцентной микроскопии определяли долю апоптотических клеток в костном мозге через 24 ч после инъекции ЦФ. Результаты представляли в виде медианы и межквартильного интервала — Me (IQR). Однократное введение ГКСК снижало TDNA на 30,3 % до 18,7 (9,92) % (p < 0,05) и на 27,9 % до 28,5 (19,48) % (p < 0,05) в костном мозге через 6 и 24 ч после введения ЦФ относительно группы позитивного контроля — 26,8 (5,39) и 39,5 (7,11) %. Трехкратная инъекция ГКСК приводила к уменьшению TDNA на 57,3 % до 15,8 (20,29) % (p < 0,05) и 35,5 % до 35,4 (1,57) % (p < 0,05) в печени через 6 и 24 ч относительно группы позитивного контроля — соответственно 37,0 (5,28) и 55,0 (4,69) %. Курсовое введение ГКСК, ЛКСК, ЛКСПК перед ЦФ вызывало значимое снижение доли апоптотических клеток соответственно на 34,3; 45,7; 34,3 % (p < 0,05) относительно группы позитивного контроля — 17,5 (4,00) %. Полученные данные свидетельствуют об антигенотоксическом действии ГКСК и антиапоптотическом влиянии исследуемых субстанций в условиях индуцированной ЦФ цитотоксичности, которые, вероятно, проявляются благодаря антиоксидантному и иммуномодулирующему действию.
Крупный рогатый скот, криофракция селезенки, криофракция плаценты, циклофосфамид, метод ДНК-комет, повреждение ДНК, апоптоз, печень, костный мозг, лабораторные мыши
Короткий адрес: https://sciup.org/142247695
IDR: 142247695 | УДК: 619:615.36:575.224.46 | DOI: 10.15389/agrobiology.2026.2.371rus
Study of the antigenotoxic effect of the bovine cryofractions of spleen and placenta under the action of cyclophosphamide
Maintaining the stability of the genome of farm animals is associated with the integrity of their cell DNA under adverse environmental conditions. This work us the first to establish the antigenotoxic effect of hydrophilic cryofraction of bovine spleen (HCBS) in the liver and bone marrow, as well as the antiapoptotic effect of HCBS, lipophilic cryofraction of bovine spleen (LCBS) and an equal mixture of lipophilic cryofraction of bovine spleen and placenta (LCBSP) in the bone marrow of mice under conditions of cyclophosphamide (CP)-induced genotoxicity. The aim of the study was to determine the effect of hydrophilic cryofraction of bovine spleen, lipophilic cryofraction of bovine spleen and a mixture of lipophilic cryofraction of bovine spleen and placenta on the genotoxic effect of cyclophosphamide in mice. The study was conducted at the All-Russian Research Veterinary Institute of Pathology, Pharmacology and Therapy (FSBSI ARVRIPP&T) in 2025. Pharmaceutical substances containing HCBS, LCBS and LCBSP, obtained at FSBSI “ARVRIPP&T”, were used. The experimental genotoxicant was Endoxan® (Baxter Oncology GmbH, Germany), which contains cyclophosphamide monohydrate (CP) as the active ingredient. Eleven experimental groups of laboratory outbred mice — males (Mus albus officinarum, n = 126) with a body weight of 26.0±2.0 g were formed. In group I (negative control), animals received a single intramuscular and intraperitoneal injection of isotonic sodium chloride solution in a volume of 0.1 and 0.5 ml, respectively (n = 6). Mice of groups II, III and IV received HCBS, LCBS and LCBSP, respectively, once intramuscularly at a dose of 0.5 ml/kg. Animals in group V served as a positive control and were given a single intraperitoneal injection of CF at a dose of 40 mg/kg and a volume of 0.5 ml (n = 12). Animals in group VI were given a single intramuscular injection of HCBS at a dose of 0.5 ml/kg and a volume of 0.1 ml before an intraperitoneal injection of CP at a dose of 40 mg/kg and a volume of 0.5 ml (n = 12). Mice from groups VII and VIII were given a single injection of LCBS and LCBSP, respectively, before the administration of CF in doses similar to those in group VI (n = 12 in each group). Animals from groups IX, X, and XI were administered HCBS, LCBS and LCBSP three times at 24-hour intervals, respectively, at a dose of 0.5 ml/kg and a volume of 0.1 ml (n = 12 in each group). Along with the final injection of the test substance, mice received a single intraperitoneal injection of CF at a dose of 40 mg/kg and a volume of 0.5 ml. Animals were sacrificed by carbon dioxide overdose in a special chamber 6 and 24 hours after CF administration. Bone marrow samples were obtained from the femurs, and liver samples were also collected. The tissues were homogenized and used for single-cell alkaline electrophoresis (the comet assay). The DNA content in the comet tail (TDNA, %) was determined 6 and 24 h after CP injection. The proportion of apoptotic cells in the bone marrow was determined 24 h after CP injection using the commercial Annexin V-IF488/PI Cell Apoptosis Detection Kit (Servicebio, China) by fluorescence microscopy. The results of the studies were presented as median and interquartile range (Me(IQR)). A single administration of HCBS reduced TDNA by 30.3 — 18.7 (9.92) % (p < 0.05) and 27.9 % — 28.5 (19.48) %, (p < 0.05) in the bone marrow 6 and 24 hours after CP administration relative to the positive control group — 26.8 (5.39) and 39.5 (7.11) %, respectively. Three-fold injection of HCBS resulted in a decrease in TDNA by 57.3 — 15.8 (20.29) (p < 0.05) and 35.5 % 35.4 (1.57) % (p < 0.05) in the liver in 6 and 24 hours vs the positive control group — 37.0 (5.28) and 55.0 (4.69) %, respectively. A course of administration of HCBS, LCBS and LCBSP before CP caused a significant decrease in the proportion of apoptotic cells by 34.3; 45.7; 34.3 % (p < 0.05), respectively, to the positive control group — 17.5 (4.00) %. The obtained data indicate the antigenotoxic effect of HCBS and the antiapoptotic effect of the studied substances under conditions of CP-induced cytotoxicity, which are probably manifested due to the antioxidant and immunomodulatory effects.
Текст научной статьи Исследование антигенотоксического действия криофракций селезенки и плаценты крупного рогатого скота в условиях действия циклофосфамида
Исследование выполнено за счет гранта Российского научного фонда ¹ 24-26-00034,
Сельскохозяйственные животные в современных условиях содержания могут подвергаться воздействию разнообразных экзогенных факторов: лекарств, микотоксинов, пестицидов и других поллютантов, способствующих повреждению их генома. Высокая технологическая нагрузка, технологический стресс, окислительный стресс также приводят к активации эндогенных процессов, дестабилизирующих геном (1, 2).
Одним из основных источников нарушения генома животных выступает повреждение ДНК в клетках, которое рассматривается как предму-тационное событие, могущее привести к изменению закодированной в ДНК информации (3, 4).
В связи с этим поддержание целостности ДНК животных остается важной задачей, для решения которой используют вещества, способные уменьшить повреждение ДНК в клетках или стимулировать ее репарацию (5). Фармацевтические субстанции, содержащие ткани животных и человека, продемонстрировали различные биологические эффекты (6). Ранее нами было показано антикластогенное действие нескольких криофракций селезенки и плаценты крупного рогатого скота на клетки костного мозга мышей при индукции цитогенетической нестабильности препаратами ал-киляторами ДНК — циклофосфамидом и митомицином, однако их влияние на целостность ДНК остается не изученным (7-9).
Циклофосфамид (ЦФ) — один из наиболее часто используемых экспериментальных мутагенов в исследованиях, связанных с изучением генотоксичности веществ. ЦФ метаболизируется в печени в цитотоксические соединения, которые взаимодействуют с ДНК и образуют сшивки ДНК-ДНК или ДНК-белок (10). Кроме того, воздействие метаболитов ЦФ индуцирует окислительный стресс в организме и приводит к увеличению повреждения ДНК свободными радикалами. Эти процессы обусловливают нарушение синтеза ДНК и транскрипции, замедление клеточного цикла и гибель клеток. При этом наибольшему воздействию ЦФ подвергаются быст-ропролиферирующие ткани (костный мозг, печень, мочевой пузырь), а токсический эффект влияет на все фазы клеточного цикла (10-12).
В настоящей работе было впервые установлено антигенотоксическое влияние гидрофильной криофракции селезенки крупного рогатого скота (ГКСК) в печени и костном мозге, а также антиапоптотическое действие ГКСК, липофильной криофракции селезенки крупного рогатого скота (ЛКСК) и равной смеси липофильной криофракции селезенки и плаценты крупного рогатого скота (ЛКПК) в костном мозге мышей в условиях индуцированной ЦФ генотоксичности.
Целью работы было определение влияния гидрофильной криофракции селезенки, липофильной криофракций селезенки и смеси липофильной криофракции селезенки и плаценты крупного рогатого скота на генотоксическое действие циклофосфамида у мышей.
Методика. Исследование проводили во Всероссийском научно-исследовательском ветеринарном институте патологии, фармакологии и терапии (ФГБНУ ВНИВИПФиТ) в 2025 году в соответствии с требованиями действующих международных и российских законодательных актов, а также комиссии по биоэтике института.
Использовали фармацевтические субстанции, содержащие гидрофильную криофракцию селезенки крупного рогатого скота (ГКСК КРС), липофильную криофракцию селезенки КРС (ЛКСК) и смесь липофильной криофракции селезенки и плаценты КРС (ЛКСПК) в равном количестве, полученные в ФГБНУ ВНИВИПФиТ (13).
Экспериментальным генотоксикантом служил Эндоксан® («Baxter
Oncology GmbH», Германия), содержащий в качестве действующего вещества циклофосфамид моногидрат (ЦФ), который используют как противоопухолевый препарат алкилирующего действия (11-13).
Были сформированы 11 экспериментальных групп лабораторных беспородных мышей-самцов ( Mus albus officinarum , n = 126) с массой тела 26,0±2,0 г, полученных в филиале «Андреевка» ФГБУН НЦБМТ ФМБА России (пос. Андреевка, Солнечногорский р-н Московской обл.). В I группе (негативный контроль) животные получали однократную внутримышечную и внутрибрюшинную инъекцию изотонического раствора хлорида натрия в объеме соответственно 0,1 и 0,5 мл ( n = 6). Мыши в II, III и IV группах получали однократную внутримышечную инъекцию соответственно ГКСК, ЛКСК и ЛКСПК в дозе 0,5 мл/кг и объеме 0,1 мл ( n = 12 в каждой группе). Животные в V группе выступали в качестве позитивного контроля, им однократно внутрибрюшинно вводили ЦФ в дозе 40 мг/кг и объеме 0,5 мл ( n = 12). Животным в VI группе однократно внутримышечно вводили ГКСК в дозе 0,5 мл/кг и объеме 0,1 мл перед внутрибрюшинной инъекцией ЦФ в дозе 40 мг/кг и объеме 0,5 мл ( n = 12). Мышам из VII и VIII групп однократно вводили соответственно ЛКСК и ЛКСПК перед введением ЦФ в дозах, аналогичных дозам в VI группе ( n = 12 в каждой группе). Животным из IX, X и XI групп трехкратно с интервалом 24 ч вводили соответственно ГКСК, ЛКСК и ЛКСПК в дозе 0,5 мл/кг и объеме 0,1 мл ( n = 12 в каждой группе). Совместно с последней инъекцией исследуемой субстанции мыши однократно внутрибрюшинно получали ЦФ в дозе 40 мг/кг и объеме 0,5 мл.
Животных выводили из эксперимента посредством передозировки углекислого газа в специальной камере через 6 и 24 ч после введения ЦФ, по 6 мышей из группы в соответствующее время. Мышей из группы негативного контроля выводили из эксперимента через 24 ч. Из бедренных костей получали образцы костного мозга, также отбирали образцы печени. Ткани гомогенизировали и использовали для проведения щелочного электрофореза единичных клеток (метод ДНК-комет) согласно опубликованной методике (14).
Для приготовления препаратов клетки заключали в агарозный гель, лизировали и проводили щелочной электрофорез в течение 30 мин при силе тока 300 мА и напряженности электрического поля 1 В/см2 с помощью источника питания Эльф-4 (ООО «ДНК-технология», Россия) и камеры для электрофореза SE-2 («Helicon», Россия). Препараты окрашивали флуоресцентным зондом dsGreen® (ООО «Люмипроб Рус», Россия), высокоселективно связывающимся с ДНК (15), затем микроскопировали при увеличении ½200, используя флуоресцентный микроскоп Микромед-3 ЛЮМ LED («Микромед», Китай), при максимуме длины волны возбуждения 488 нм. Учитывали не менее 100 клеток на препарат, за исключением клеток с сильно поврежденным нуклеоидом, с помощью цифрового видеоокуляра UA1600CA («ToupCam», Китай). Степень повреждения ДНК оценивали с помощью программы CASP 1.2.2 («Comet Assay Software Project», Польша). В качестве определяемого параметра использовали содержание ДНК в «хвосте кометы» (TDNA, %), который прямо зависит от величины повреждения ДНК (16).
Величину апоптотической гибели клеток исследовали в костном мозге лабораторных мышей через 24 ч после введения ЦФ по их окрашиванию аннексином V и йодистым пропидием с помощью коммерческого набора Annexin V-IF488/PI Cell Apoptosis Detection Kit («Servicebio», Китай). Согласно инструкции производителя изготавливали препараты костного мозга, окрашенные флуоресцентными красителями, и микроскопировали при увеличении ½400, используя флуоресцентный микроскоп Микромед-3 ЛЮМ LED («Микромед», Китай) при максимуме длины волны возбуждения 488 нм. Учитывали не менее 200 клеток для одного животного и определяли процент апоптотических клеток (17).
Статистическую обработку данных проводили с помощью пакетов программ STATISTICA 10 («StatSoft, Inc.», США). Нормальность распределения количественных параметров проверяли с помощью критериев Колмогорова-Смирнова и Шапиро-Уилка. Выборки сравнивали с использованием U-критерия Манна-Уитни (18). Различия в выборках считали статистически значимыми при p < 0,05. Полученные результаты представляли как Me (IQR) — медиану (межквартильный размах между первым и третьим квартилем).
Результаты. Нами были получены микрофотографии «ДНК-комет» клеток печени и костного мозга, исследуемых мышей (рис. 1).
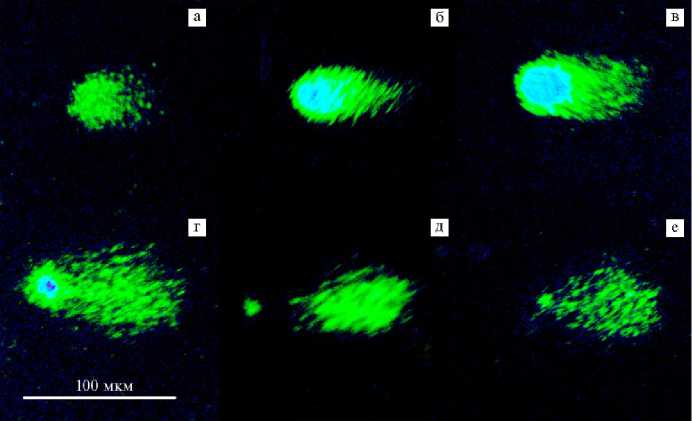
Рис. 1. Микрофотографии ДНК клеток беспородных мышей ( Mus albus officinarum ) с различной степенью повреждения : a, б, в, г — ДНК клеток с увеличивающейся степенью повреждения; д, е — ДНК клеток, исключенных из анализа (сильное повреждение нуклеоида) (окрашивание dsGreen®, увеличение ½200, микроскоп Микромед-3 ЛЮМ LED, «Микромед», Китай) (Всероссийский научно-исследовательский ветеринарный институт патологии, фармакологии и терапии, 2025 год).
Результаты исследования электрофоретической подвижности ДНК единичных клеток печени и костного мозга мышей продемонстрировали различия в группах (рис. 2).
Показатель TDNA в негативном контроле (I группа) составил 13,0 (6,52) % в клетках печени и 5,7 (8,74) % в костном мозге. Однократное введение ГКСК, ЛКСК и ЛКСПК не приводило к значимому изменению TDNA через 6 и 24 ч ни в клетках печени, ни в костном мозге мышей (II, III и IV группы). Эти данные могут свидетельствовать об отсутствии у исследуемых фармацевтических субстанций генотоксической активности. Через 6 ч после введения ЦФ в дозе 40 мг/кг (V группа) величина TDNA статистически значимо увеличилась в 2,8 и 4,7 раза (p < 0,05) соответственно в клетках печени и костного мозга относительно I группы и составила 37,0 (5,28) и 26,8 (5,39) %. Через 24 ч после введения ЦФ TDNA в клетках печени мышей значимо возрастало до 55,0 (4,69), а в костном мозге до 39,5 (7,11) % (p < 0,05). Таким образом, количество повреждений ДНК увеличивалось в 4,2 и 6,9 раза в клетках печени и костного мозга мышей в группе позитивного контроля относительно I группы.
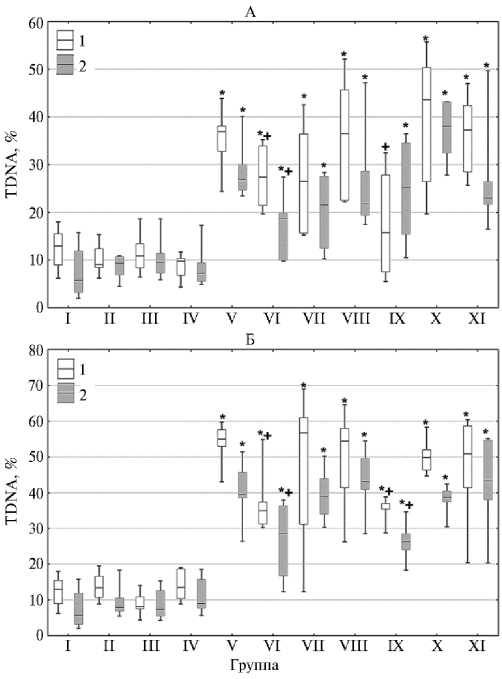
Рис. 2. Содержание ДНК в «хвосте кометы» (TDNA) в клетках печени (1) и костного мозга (2) у беспородных мышей ( Mus albus officinarum ) под влиянием гидрофильной криофракции селезенки крупного рогатого скота (ГКСК), липофильной криофракции селезенки КРС (ЛКСК) и смеси липофильной криофракции селезенки и плаценты КРС (ЛКСПК) через 6 ч (А) и 24 ч (Б) после инъекции циклофосфамида: I-XI — исследуемые группы животных (описание в см. в разделе «Методика») . Бокс — диапазон между первым и третьим квартилем выборки, горизонтальная черта в боксе — медиана, вертикальные отрезки — разброс данных (Всероссийский научноисследовательский ветеринарный институт патологии, фармакологии и терапии, 2025 год).
* Различие с негативным контролем (I группа) статистически значимо при p < 0,05; + различие с позитивным контролем (V группа) статистически значимо при p < 0,05.
В VI-XI группах, за исключением IX группы, введение ЦФ индуцировало значимое увеличение TDNA относительно I группы в обеих исследуемых тканях через 6 и 24 ч. Так, при однократном введении фармацевтической субстанции вместе с ЦФ через 6 ч TDNA в печени составило 27,4 (12,48), 26,5 (20,76) и 36,5 (23,17) % соответственно в VI, VII и VIII группах. Величина TDNA в клетках костного мозга через 6 ч в этих группах была равной 18,7 (9,92), 21,6 (15,00) и 21,8 (9,19) %. В VI группе через 24 ч TDNA увеличивалось до 34,9 (6,14) % в гепатоцитах и до 28,5 (19,48) % в клетках костного мозга относительно группы негативного контроля. Таким образом, при однократном введении ГКСК с ЦФ (VI группа) было обнаружено снижение повреждения ДНК, которое определялось статистически значимым уменьшением TDNA на 25,8 и 36,5 % (p < 0,05) в печени, а также на 30,3 и 27,9 % (p < 0,05) в костном мозге соответственно через 6 и 24 ч относительно группы, получавшей только ЦФ (V группа). TDNA в VII группе через 24 ч после введения ЦФ составляло 56,7 (29,75) и 39,0 (9,89) % соответ- ственно для клеток печени и костного мозга, а в VIII группе — 54,4 (16,56) % для клеток печени и 43,2 (8,64) % для костного мозга. Хотя эти значения в некоторых случаях и были несколько ниже таковых в группе позитивного контроля, мы не выявили статистически значимого снижения TDNA при введении ЛКСК и ЛКСПК.
Сходные тенденции были обнаружены при изучении влияния трехкратного введения исследуемых фармацевтических субстанций на индуцированное ЦФ генотоксическое воздействие. Как и при однократном введении ГКСК, курсовое применение ГКСК перед введением ЦФ (IX группа) через 6 ч приводило к статистически значимому снижению TDNA в клетках печени на 57,3 % относительно группы позитивного контроля (V группа). При этом показатель TDNA был равен 15,8 (20,29) % и не имел статистически значимых отличий от группы негативного контроля (I группа). В клетках костного мозга из мышей IX группы через 6 ч после введения ЦФ TDNA равнялось 25,3 (19,07) %, однако этот показатель не имел значимых отличий от V группы. Через 24 ч после введения ЦФ и в клетках печени, и в клетках костного мозга животных из IX группы было отмечено значимое снижение TDNA на 35,5 и 33,2 % (p < 0,05) относительно V группы. Значение TDNA при этом было равным 35,4 (1,57) и 26,4 (4,34) % соответственно в клетках печени и костного мозга.
Курсовое введение ЛКСК (X группа) и ЛКСПК (XI группа) перед инъекцией ЦФ не вызвало значимого изменения TDNA относительно V группы, как и при однократном применении липофильных криофракций. В клетках печени мышей из X группы TDNA составило 43,6 (23,88) и 49,8 (5,72) % через 6 ч и 24 ч после введения ЦФ. В клетках костного мозга этот показатель был равен 38,1 (10,69) и 38,6 (2,96) % через 6 ч и 24 ч после введения ЦФ. В XI группе через 6 ч после введения ЦФ TDNA составлял 37,3 (13,97) и 23,0 (4,75) %, а через 24 ч после введения ЦФ — 50,9 (17,21) и 43,3 (16,57) % в клетках печени и костного мозга, соответственно. Следует также отметить, что мы не выявили значимых различий в повреждении ДНК в VI и IX группах. Таким образом, однократное и курсовое введение ГКСК снижало количество повреждения ДНК, индуцированного введением ЦФ.
Поскольку вызванное метаболитами ЦФ повреждение ДНК и цитотоксическое действие способно индуцировать гибель клеток посредством апоптоза, влияние на который рассматривается как один из механизмов антигенотоксичности, мы определили содержание апоптотических клеток в костном мозге мышей исследуемых групп через 24 ч после финального введения фармацевтических субстанций и ЦФ (11, 19). Так, в костном мозге у мышей в негативном контроле содержание аннексин V-позитивных клеток составило 8,0 (3,50) %. Применение ГКСК, ЛКСК и ЛКСПК (II, III и IV группы) не приводило к значимому изменению содержания апоптотических клеток (показатель колебался в пределах 5,0-7,5 %) в костном мозге относительно I группы (рис. 3.). Введение ЦФ (V группа) индуцировало значимое увеличение доли апоптотических клеток до 17,5 (4,00) % (p < 0,05) в костном мозге мышей. Однократное введение ГКСК, ЛКСК и ЛКСПК вместе с ЦФ (VI, VII и VIII группы) приводило к снижению содержания апоптотических клеток соответственно до 11,0 (4,00); 10,0 (8,00) и 12,5 (6,00) %. Однако только при введении ЛКСК (VII группа) наблюдалось значимое снижение доли апоптотических клеток на 42,9 % (p < 0,05) относительно группы позитивного контроля.
Вместе с тем при курсовом применении исследуемых фармацевтических субстанций перед введением ЦФ (IX, X и XI группы) мы обнару-376
жили статистически значимое снижение содержания апоптотических клеток в костном мозге относительно V группы на 34,3; 45,7 и 34,3 % (p < 0,05) при введении соответственно ГКСК, ЛКСК и ЛКСПК — доля таких клеток равнялась 11,5 (8,00); 9,5 (4,00) и 11,5 (9,00) %. При этом как однократное, так и курсовое применение гидрофильных и липофильных криофракций с ЦФ приводило к снижению содержания апоптотических клеток до такой величины, которая достоверно не отличалась от значений негативного контроля. Таким образом, курсовое применение ГКСК, ЛКСК и ЛКСПК оказывало антиапоптотическое действие на клетки костного мозга в условиях индуцированной ЦФ цитотоксичности.
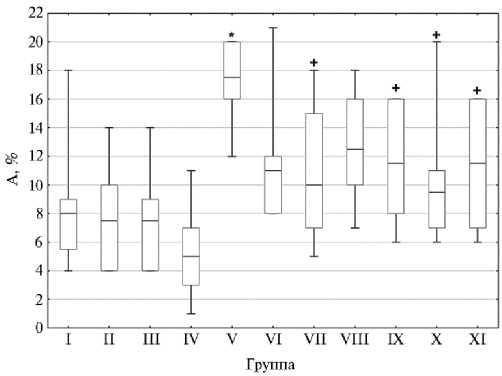
Рис. 3. Доля апоптотических клеток (А) в костном мозге у беспородных мышей ( Mus albus officinarum ) под влиянием гидрофильной криофракции селезенки крупного рогатого скота (ГКСК), липофильной криофракции селезенки КРС (ЛКСК) и смеси липофильной криофракции селезенки и плаценты КРС (ЛКСПК) через 24 ч после инъекции циклофосфамида: I-XI — исследуемые группы животных (описание в см. в разделе «Методика») . Бокс — диапазон между первым и третьим квартилем выборки, горизонтальная черта в боксе — медиана, вертикальные отрезки — разброс данных (Всероссийский научно-исследовательский ветеринарный институт патологии, фармакологии и терапии, 2025 год).
* Различие с негативным контролем (I группа) статистически значимо при p < 0,05; + различие с позитивным контролем (V группа) статистически значимо при p < 0,05.
Электрофоретическая подвижность ДНК единичных клеток при воздействии ЦФ может зависеть от дозы генотоксиканта, исследуемой ткани и времени, которое прошло после введения ЦФ, поскольку соединение требует биоактивации в печени (20, 21, 22). Так, образование сшивок ДНК под действием ЦФ может снизить подвижность ДНК в клетках печени и костного мозга при щелочном электрофорезе после инъекции препарата алки-лятора (23). Исследования показывают индуцированное ЦФ увеличение повреждения ДНК клеток, связанного с возрастанием подвижности ДНК через 4 и 24 ч в лейкоцитах крови и через 24 ч в клетках печени и костного мозга мышей относительно негативного контроля (24-26). Эти данные соответствуют обнаруженному нами значимому увеличению повреждения ДНК через 24 ч на 48,7 % в печени (p < 0,05) и на 32,1 % костном мозге (p < 0,05) относительно 6 ч после введения ЦФ в дозе 40 мг/кг.
Выявленное нами ДНК-защитное действие ГКСК подтверждается уменьшением повреждения ДНК лимфоцитов у мышей после воздействия ионизирующей радиации при использовании экстракта плаценты (27). Снижение повреждения ДНК под действием ГКСК может происходить в связи с активацией антиоксидантной системы мышей. Ранее было показано сни- жение содержания малонового диальдегида, увеличение концентрации витаминов А и Е и активности каталазы и глутатионпероксидазы при введении ГКСК и содержащих ее препаратов мышам или свиньям (9, 28). В других исследованиях также продемонстрировано антиоксидантное действие экстрактов различных тканей КРС и свиней, которые содержали витамины, аминокислоты и другие молекулы с антиоксидантным действием (29, 30).
Интересно, что обнаруженное нами ДНК-защитное действие ГКСК при ЦФ индуцированной генотоксичности в клетках печени и костного мозга проявлялось как при однократном введении гидрофильной криофракции селезенки, так и при курсовом применении этой фармацевтической субстанции. При этом полученные нами ранее данные по частоте полихроматофильных эритроцитов с микроядрами в костном мозге мышей при введении животным ЦФ или другого препарата алкилятора ДНК, митомицина, демонстрировали статистически значимое снижение кластогенного эффекта этих веществ только при курсовом применении ГКСК (7, 8, 31). Кроме того, снижение повреждения митохондриальной ДНК митомицином в клетках печени под действием ГКСК также было выявлено только при курсовом применении этой фармакологической субстанции (8). Показано снижение содержания провоспалительных IL-2, TNF α и изменение концентрации других цитокинов (повышение IFN- α и IFN- γ , обладающих антимутагенным действием) в сыворотке крови мышей после введения ЦФ, вызванное применением экстракта селезенки телят, которое сопровождалось уменьшением экспрессии ядерного транскрибционного фактора Nf-Kb в спленоцитах (32). Nf-Kb играет ключевую роль в развитии воспалительных реакций в иммунных клетках (33). Таким образом, уменьшение повреждения ДНК в печени и костном мозге и снижение содержания апопто-тических клеток в костном мозге мышей, вероятно, может быть обусловлено иммуномодулирующим действием ГКСК и других исследуемых фармацевтических субстанций. Оно проявляется в противовоспалительном эффекте на организм в условиях воздействия генотоксиканта, поскольку ГКСК и препараты, содержащие эту фармацевтическую субстанцию, проявляли свою иммуномодулирующую активность при курсовом применении (28, 34). При этом противовоспалительная активность веществ может быть одним из механизмов их антигенотоксических свойств (23). Кроме того, экстракт плаценты также оказывал противовоспалительное действие по отношению к клеткам костного мозга у мышей (29).
Следует отметить, что при введении ЦФ антиапоптотическое действие ГКСК и ЛКСПК проявилось только при курсовом применении субстанций, а при использовании ЛКСПК однократное и трехкратное введение значимо снижало долю апоптотических клеток в костном мозге. При этом ДНК-защитное действие ГКСП проявлялось и при однократном, и при курсовом применении и не было выявлено при введении липофильных криофракций. Можно предположить, что обнаруженное нами снижение ЦФ-индуцированного апоптоза в меньшей мере было связано с уменьшением повреждения ДНК клеток метаболитами ЦФ, но в большей степени могло быть обусловлено влиянием исследуемых фармацевтических субстанций на процессы внутриклеточной регуляции. Это подтверждают данные о том, что ЦФ-индуцированная цитотоксичность проявляется в синергетическом воздействии его метаболитов: горчичного фосфорамида, алкилирующего ДНК, и проапоптотического альдегида — гидроксипропаналя, который вызывает снижение синтеза антиапоптотических белков Bcl-2 и Bcl-xL, активирует MAPC пути клеточной регуляции, что приводит к остановке клеточного цикла и апоптозу (35, 36). В пользу этого также свидетельствуют данные об активации пролиферативных регуляторных факторов (RSK1p90, ELK1, c-Myc, GATA-1, GATA-2), увеличении экспрессии различных цитокинов и особенно G-CSF, который приводил к активации JAK2/STAT3 пути в клетках костного мозга мышей в условиях индуцированной ЦФ иммуносупрессии под действием экстрактов селезенки телят (32, 37). При этом активация JAK2/STAT3 пути в клетках мозга такими индукторами гемопоза, как эритропоэтин, индуцировала увеличение экспрессии антиапоптотических белков Bcl-2 и Bcl-x и уменьшение апоптоза (38). Кроме того, было показано ан-тиапоптотическое действие экстракта печени свиньи (39).
Итак, внутримышечное введение гидрофильной криофракции селезенки крупного рогатого скота индуцировало значимое снижение количества повреждений ДНК в клетках печени и костного мозга мышей через 6 и 24 ч после введения генотоксиканта — циклофосфамида. Эти данные свидетельствуют о наличии у гидрофильной криофракции селезенки КРС ДНК-защитных свойств, которые проявляются, вероятно, благодаря антиоксидантной, иммуномодулирующей активности гидрофильной криофракции. Гидрофильная криофракция селезенки КРС, липофильная криофракция селезенки КРС, а также смесь липофильной криофракции селезенки и плаценты КРС при внутримышечном введении проявляли антиапоптотиче-ское действие по отношению к клеткам костного мозга в условиях цитотоксического влияния циклофосфамида. Этот эффект, возможно, был связан с противовоспалительным иммуномодулирующим влиянием исследуемых фармацевтических субстанций и проявлялся преимущественно при курсовом применении.
ФГБНУ Всероссийский научно-исследовательский ветеринарный институт патологии, фармакологии и терапии,
