Исследование кинетики аланинаминотрансферазной активности в картофеле

Автор: Бландов А.Н., Муста оглы Н.М.
Журнал: Вестник Воронежского государственного университета инженерных технологий @vestnik-vsuet
Рубрика: Фундаментальная и прикладная химия, химическая технология
Статья в выпуске: 4 (66), 2015 года.
Бесплатный доступ
Аминотрансферазы (трансаминазы) катализируют межмолекулярный перенос аминогруппы с аминокислот на кетокислоты, причем коферментом в этой реакции служит пиридоксальфосфат, который выполняет роль промежуточного акцептора аминогруппы. Например, аланинаминотрансфераза (АлАТ) катализирует реакцию переаминирования между аланином и α-кетоглутаровой кислотой. Определены кинетические параметры аланинаминотрансферазы картофеля методом Лайнуивера-Берка: Km = 0,298 ммоль/л, V max = 0,0151 мкмоль/мин. При больших концентрациях субстрата было обнаружено снижение скорости ферментативной реакции. Поскольку степень конверсии субстрата в продукт в условиях опыта не превышает 1 %, это говорит о явлении субстратного ингибирования. Используя предположение об образовании неактивного двухсубстратного комплекса, проанализирована соответствующая кинетическая схема для бесконкурентного ингибирования, где роль ингибитора играет субстрат. Выведено уравнение Михаэлиса-Ментен и Лайнуивера-Берка для этого случая и, путем дифференцирования, определено положение точки экстремума на графике Лайнуивера-Берка: Ki = [ S ]2min/ Km. Это позволило рассчитать константу ингибирования субстратом, которая составила Ki = 0,428 ммоль/л.
Аланинаминотрансфераза (алат), субстратное ингибирование, константа михаэлиса, максимальная скорость реакции, константа ингибирования
Короткий адрес: https://sciup.org/14043236
IDR: 14043236 | УДК: 547.772
Research of kinetics alaninaminotransferasе activities in potatoes
Aminotransferases (transaminases) catalyze intermolecular transfer of an amino group from amino acids on ketoacids, and as a coenzyme pyridoxal phosphate which carries out a role of an intermediate acceptor of an amino group serves in this reaction. For example, an alaninaminotransferase (AlAT) catalyzes reaction of transamination between alanine and α-ketoglutaric acid. Kinetic parameters of an alaninaminotransferase of potatoes are determined by Lineweaver-Burk plot: Km = 0.298 mmol/l, V max = 0.0151 µmol/min. The reduction of enzyme reaction rate under high concentration of a substrate was revealed. As extent of conversion of a substrate in a product doesn't exceed in our conditions 1%, it speaks about the phenomenon of inhibition by substrate. Using the assumption of formation of an inactive bisubstrate complex, we analysed the corresponding kinetic scheme for uncompetitive inhibition where the role of inhibitor plays a substratum, removed the equation Miсhaelis-Menten and Linewiever-Burk for this case and by differentiation defined the provision of a point of an extremum on Linewiever-Burk plot: : Ki = [ S ]2min / Km. It allowed to calculate an inhibition constant a substratum which is Ki = 0.428 mmol/l.
Текст научной статьи Исследование кинетики аланинаминотрансферазной активности в картофеле
Аминотрансферазы являются одними из ключевых ферментов обмена аминокислот и кетокислот. Они представлены как в тканях человека и животных, так и растений и микроорганизмов. По международной классификации их относят к классу трансфераз, подкласс переносящие азотистые группы, подподкласс аминотрансферазы. Наиболее известными их представителями являются аланин- и аспартатаминотрансфераза (КФ 2.6.1.2 и 2.6.1.1), катализирующие следующие реакции трансаминирования (рисунок 1)[1, 2].
При этом в клетках происходит синтез необходимых аминокислот из кетокислот, являющихся метаболитами гликолиза и цикла Кребса, либо наоборот превращение аминокислот в кетокислоты с последующим их использованием на энергетические нужды. Учитывая обратимость этих реакций, равновесия могут смещаться в ту или другую сторону в зависимости от потребностей клеток.
|
COOH |
COOH |
|||||
|
CH 3 |
+ |
CO |
аланинамино- |
CH 3 |
CH NH 2 |
|
|
CH NH 2 COOH |
CH 2 |
CO COOH |
+ |
CH 2 CH 2 COOH |
||
|
CH 2 COOH |
трансфераза |
|||||
|
аланин |
α-кетоглутаровая кислота |
пировиноградная кислота |
глутаминовая кислота |
|||
|
б |
||||||
|
COOH CH 2 |
+ |
COOH CO |
аспартатамино- |
COOH CH 2 |
COOH CH NH 2 |
|
|
CH NH 2 COOH |
CH 2 |
CO COOH |
+ |
CH 2 CH 2 COOH |
||
|
CH 2 COOH |
трансфераза |
|||||
|
аспарагиновая кислота |
α-кетоглутаровая кислота |
щавелево-уксусная кислота |
глутаминовая кислота |
|||
|
Рисунок 1. Реакции трансаминирования: а |
– аланинаминотрансфераза; б – |
аспартатаминотрансфераза |
||||
Эти ферменты (сокращенно АЛТ, АСТ) широко используются в медицинской диагностике заболеваний, связанных с цитолизом – гепатиты, инфаркты, т. к. они являются в основном внутриклеточными ферментами и их уровень в крови резко повышается при массивном цитолизе (разрушении клеток). В связи с этим их называют индикаторными ферментами [3-7].
В растениях эти ферменты выполняют аналогичные биохимические функции, однако гораздо меньше изучались, чем в организме человека и животных. Так, в работе [8] была изучена динамика активности АЛТ и АСТ при прорастании семян сои и было показано, что активность АЛТ в семядолях значительно повышается на третьи сутки проращивания, а через пять суток резко снижается (в три раза по сравнению с первыми сутками). Это связано, по-видимому, с тем, что семядоли значительно атрофируются к этому моменту. Однако активность АЛТ в проростках повышается в три раза на пятые сутки роста. Это может быть связано, по мнению авторов, с участием пирувата в синтезе глюкозы по реакциям глюконеогенеза [8-12]. Активность АСТ в семядолях также значительно повышается на третьи сутки, а на пятые сутки возвращается к исходным значениям. В проростках она также значительно повышается на 3-5 сутки. Авторы объясняют это необходимостью синтеза α-кетоглутарата, который занимает централь- ное место в синтезе аминокислот и затем белков, в том числе ферментов [8-12].
Однако в литературе не изучался более широкий ряд растительных объектов, а также кинетические параметры рассматриваемых ферментов. Поэтому целью настоящего исследования стало изучение активности и кинетических параметров АЛТ в растительном сырье. В качестве объекта исследования нами был выбран картофель.
Между тем, ранее было показано, что активность АЛТ может служить показателем свежести рыбного сырья [13-15]. Поэтому, изучение активности АЛТ в растительном сырье может представлять интерес при анализе пищевого сырья и контроле качества готовых продуктов, о возможном влиянии химических добавок, в частности, консервантов и образующихся из них вторичных метаболитов на ферментативную активность пищевых систем, что делает энзиматические исследования актуальными и приложимыми для решения практических задач пищевой промышленности.
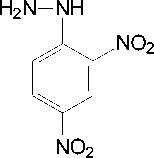
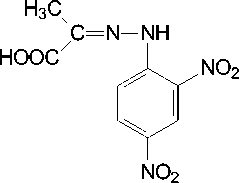
По количеству образовавшейся пировиноградной кислоты можно судить об активности фермента. Пировиноградную кислоту определяют колориметрически по цветной реакции с 2,4-динитрофенилгидразином, приводящей к образованию окрашенного 2,4-динитрофенилгидразона (рисунок 2).
CH 3
CO +
COOH пировиноградная кислота
- H 2 O
2,4-динитро-фенилгидразин
2,4-динитрофенилгидразон (красно-бурого цвета)
Рисунок 2. Цветная реакция пировиноградной кислоты с 2,4-динитрофенилгидразином
Субстратная смесь, приготовленная следующим образом: в 100 мл фосфатного буфера с pH 7,4 растворяют 1,78 г DL-аланина (или 0,89 г α-аланина) и 29,2 мг α-кетоглутаровой кислоты (смесь хранится в замороженном виде); 0,02 %-й раствор 2,4-динитрофенилгидразина в 1н HCl; 0,4н раствор NaOH. Фотоэлектрический колориметр ФЭК-Н-57 с зеленым светофильтром с λ = 540 нм и кюветами с рабочим расстоянием 10 мм; термостат на 38 °С; картофель.
Определение активности аланинаминотрансферазы
Навеска картофеля 2 г растиралась в ступке с 10 мл воды и полученный гомогенат фильтровался через бумажный фильтр. Затем к 0,2 мл фильтрата гомогената добавляли 0,5 мл субстратной смеси, выдерживали при температуре 38 ºС в течение 30 мин, после чего добавля- ли 0,5 мл раствора ДНФГ и через 15 мин 5 мл 0,4н NaOH. Определяли оптическую плотность раствора относительно холостой пробы, полученной при использовании вместо фильтрата гомогената 0,2 мл дистиллированной воды.
Формулы для расчетов
C = D мкмоль; (1)
E- 1 ,
V = C мкмоль/мин, (2)
t, где ε = 0,45; l = 1 см; t = 30 мин.
Коэффициент экстинкции ε определяется по ряду стандартных растворов, содержащих ДНФГ пировиноградной и α-кетоглутаровой кислоты. Результаты эксперимента приведены в таблице 1 и показаны на рисунке 3.
Т а б л и ц а 1
Численные результаты опытов
|
№ разведения |
[ S ], ммоль/л |
1/[ S ], л/ммоль |
D 1 |
D 2 |
D средн |
V средн, мкмоль/мин |
1/ V , мин/мкмоль |
|
1 |
1,429 |
0,7 |
0,165 |
0,150 |
0,1575 |
0,01167 |
85,71 |
|
2 |
0,714 |
1,4 |
0,140 |
0,150 |
0,145 |
0,01074 |
93,10 |
|
3 |
0,476 |
2,1 |
0,120 |
0,130 |
0,125 |
0,00926 |
108,00 |
|
4 |
0,357 |
2,8 |
0,125 |
0,100 |
0, 1125 |
0,00833 |
120,00 |
|
5 |
0,286 |
3,5 |
0,120 |
0,085 |
0,1025 |
0,00759 |
131,71 |
|
6 |
0,238 |
4,2 |
0,085 |
0,100 |
0,0925 |
0,00685 |
145,95 |
|
7 |
0,204 |
4,9 |
0,080 |
0,090 |
0,085 |
0,00630 |
158,82 |
|
8 |
0,179 |
5,6 |
0,080 |
0,075 |
0,0775 |
0,00574 |
174,19 |
Определены кинетические параметры аланинаминотрансферазы картофеля методом Лайнуивера-Берка: К m = 0,298 ммоль/л, V max = 0,0151 мкмоль/мин.
Из графика (рисунок 3) видно, что в области больших концентраций наблюдается небольшое отклонение графика вверх. Это может говорить о наличии субстратного ингибирования.
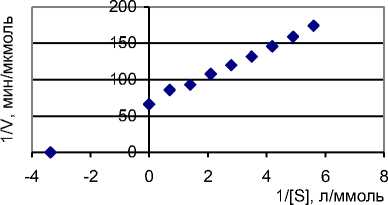
Рисунок 3. Определение активности аланинаминотрансферазы
Субстратное ингибирование (ингибирование избытком субстрата) можно рассматривать в качестве частного случая бесконкурентного ингибирования. Помимо изменения свойств среды при больших [ S ] причиной такого явления может быть образование неактивного комплекса с двумя молекулами субстрата ES 2 и тогда субстрат фактически играет роль бесконкурентного ингибитора, а кинетическая схема выглядит следующим образом:
S
V = V max ------ ■ .
K m + [ S ] + [ S L
K i
Тогда уравнение Лайнуивера-Берка будет иметь следующий вид:
1 1 fl K [ S ] )
= 1 + -m +^
V V max V [ S ] K ; J
E + S _ w ES -----► E + P k-i
* c [ES] [S]
+ S Ki= ------ t [ES2]
ES 2
,(3)
здесь K i – константа ингибирования субстратом, т.е. константа диссоциации двойного фер-мент-субстратного комплекса. Проведем для этого случая вывод уравнения Михаэлиса-Ментен:
V = k 2 [ ES ] ; V max = k 2 [ E ] , ;
k 1 [ E ][ S ] = k -1 [ ES ] + k 2 [ ES ] ; (4)
[ E H k _ , + k 2
[ ES ] k , m .
Запишем уравнение материального баланса по ферменту, в котором в данном случае будет три слагаемых:
[ E ] o = [ E ] + [ ES ] + [ ES 2 ] = [ E ] + [ ES ] f 1 +[ S 1 '■
V K i 2 7
Выразим отсюда [ E ] и подставим в выражение константы Михаэлиса:
При этом на графике Лайнуивера-Берка в области больших концентраций субстрата наблюдается отклонение от линейности, и на графике, как в нашем случае, будет наблюдаться минимум (см. рисунок 3).
Для того чтобы найти [ S ], соответствующую минимуму, продифференцируем обратную скорость (обозначим ее y ) по обратной концентрации (обозначим ее x ). В точке экстремума производная обращается в ноль. Исходя из этого, находим [ S min ]:
_ = y;
V [ S ]
У = — 1 + K m x +— ; (8)
Vmax V dy = _LfK + * 1=JLfK -[St' dx Vmax V m X2 Ki J V,,, V m Ki
[ s min ] = K K Ki^ ;
K i =
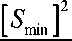
= 0;
[ E ] , - [ ES ] ' ' +K
[ ES ]
[ S ]
—= K m ; (5)
[ s ][ E ]0- = k + [ S ] K L±I S 1
L J [ ES ] m L J K i
.
Теперь выразим отсюда [ ES ] и преобразуем выражение в форму, похожую на уравнение Михаэлиса-Ментен:
[ ES ] = [ E ]>
[ S ]
K m + [ S ] [ 1 + [ - ]
V K J
Непосредственно по графику рассчитать K i невозможно, т.к. на первом участке кривой зависимость не является линейной. Однако в точке минимума на графике Лайнуивера-Берка производная d(1/ V )/d(1/[ S ]) обращается в ноль. Отсюда следует, что K i = [ S min ]2/ K m . Это дает возможность рассчитать константу ингибирования исходя из точки экстремума на графике.
Кинетические параметры реакции определяются экстраполяцией линейного участка графика в области малых концентраций до пересечения с осями, как и в обычном случае.
Для проверки предположения об ингибировании субстратом мы провели опыт, взяв меньшее количество фермента (1,5 г картофеля в том же объеме воды) и увидели явное отклонение вверх при больших концентрациях субстрата, что подтверждает предположение о субстратном ингибировании. Результаты эксперимента представлены в таблице 2 и показаны на рисунке 4.
Т а б л и ц а 2
Численные результаты опытов
|
№ разведения |
[ S ], ммоль/л |
1/[ S ], л/ммоль |
D 1 |
D 2 |
D средн |
V средн, мкмоль/мин |
1/ V , мин/мкмоль |
|
1 |
1,429 |
0,7 |
0,165 |
0,150 |
0,1575 |
0,01167 |
85,71 |
|
2 |
0,714 |
1,4 |
0,140 |
0,150 |
0,145 |
0,01074 |
93,10 |
|
3 |
0,476 |
2,1 |
0,120 |
0,130 |
0,125 |
0,00926 |
108,00 |
|
4 |
0,357 |
2,8 |
0,125 |
0,100 |
0, 1125 |
0,00833 |
120,00 |
|
5 |
0,286 |
3,5 |
0,120 |
0,085 |
0,1025 |
0,00759 |
131,71 |
|
6 |
0,238 |
4,2 |
0,085 |
0,100 |
0,0925 |
0,00685 |
145,95 |
|
7 |
0,204 |
4,9 |
0,080 |
0,090 |
0,085 |
0,00630 |
158,82 |
|
8 |
0,179 |
5,6 |
0,080 |
0,075 |
0,0775 |
0,00574 |
174,19 |
|
300 250 |
♦ |
|
200 |
|
|
150 |
|
|
100 |
|
|
50 |
|
|
—♦ 0 |
-5 0 5 10
1/[S], л/ммоль
Рисунок 4. Определение активности аланинаминотрансферазы
Из графика, показанного на рисунке 4, видно, что минимум на графике Лайнуивера-Берка соответствует четвертому разведению, т.е. [S]min = 0,357 ммоль/л. Это позволяет вычислить константу ингибирования субстратом по формуле Ki = [S]2min /KM = 0,428 ммоль/л. Константа Михаэлиса здесь определяется экстраполяцией линейной части графика в области малых концентраций субстрата до пересечения с осями и она численно совпадает с определенной в первом опыте. Однако Vmax = 0,0128 мкмоль/мин, что меньше, чем в первом опыте, т.к. использовалась меньшая масса картофеля и соответственно меньшее количество фермента.
Аналогичная ситуация наблюдалась в работе [16] для АЛТ чеснока, для которой были найдены K M = 0,0206 ммоль/л и K i =0,0777 ммоль/л. Отсюда видно, что сродство к субстрату для АЛТ чеснока гораздо больше, чем для картофеля.
Список литературы Исследование кинетики аланинаминотрансферазной активности в картофеле
- Комов В.П., Шведова В.Н. Биохимия. 3-е изд. М.: Дрофа, 2008. 638 с.
- Смирнов В.А., Климочкин Ю.Н. Ферменты. Классификация и номенклатура. Самара, 2008. 42 с.
- Николаев А.Я. Биологическая химия. 3-е изд. М.: Медицинское информационное агентство. 2004. 566 с.
- Северин Е.С. Биохимия: учебник. 4-е изд. М.: ГЭОТАР-Медиа, 2006. 784 с.
- Северин Е.С., Николаев А.Я. Биохимия. Краткий курс с упражнениями и задачами. М.: ГЭОТАР-МЕД, 2001. 448 с.
- Щербак И.Г. Биологическая химия. СПб.: Издательство СПбГМУ, 2005. 480 с.
- Березов Т.Т., Коровкин Б. Ф. Биологическая химия: учебник. 3-е изд. М.: Медицина, 1998, 704 с.
- Бездудная О.Ф., Калиман П.А. Изменение содержание белка и трансаминирования аминокислот при прорастании семян сои (GLICINE MAX L.)//Вiсник Харкiвського нацiонального унiверситету iменi В.Н.Каразiна. Серiя: бiологiя. 2006. Вып. 4. № 748. С. 17-20.
- Плакунов В.К. Основы энзимологии. М.: Логос, 2002. 128 с.
- Lancien M., Hodges M. Enzyme redundancy and the importance of 2-oxoglutarate in higher plant ammonia assimilation//Plant Physiol. 2000. V. 123. P. 817-824.
- Жмурко В.В., Джамаев В.Ю. Дегидрогеназная активность и содержание растворимого белка в корнях проростков сои (Glicine max L.) при оптимальной и пониженной температурах//Физиология и биохимия культурных растений. 1999. Т. 31. № 4. С. 308-313.
- Жмурко В.В., Джамеев В.Ю. Рiст, завиток та фiзiолого-бiохiмiчнi процеси у теплолюбивих культур при адаптацїї до пониженої температури//Фiзiологiя рослин в Українi на межi тисячолiть. 2001. Т. 2. С. 182-189.
- Шлейкин А.Г., Данилов Н.П., Шарапова Т.А. Технологические и медико-биологические аспекты действия трансглутаминазы//Известия СПбГУНиПТ. 2009. № 3/4. С. 47-49.
- Shleikin A.G., Danilov N.P. Evolutionary-biological peculiarities of transglutaminase. Structure, physiological functions, application//Journal of Evolutionary Biochemistry and Physiology. 2010. V. 47. № 1. P. 1-14.
- Shleikin A.G., Danilov N.P., Ternovskoy G.V. Modification of food products properties by use of transglutaminase. Original Research//Procedia Food Science. 2011. V. 1. P. 1568-1572.
- Маджидов Ш.Б. Изучение аминотрансферазной активности растительного сырья. СПб.: Университет ИТМО. 2014. 76 с.
