Изменчивость репродуктивных признаков у генотипов подсолнечника с различным содержанием жирных кислот и токоферолов в семенах

Автор: Демурин Я.Н., Рубанова О.А., Чебанова Ю.В., Земцева Т.А., Епишкина А.В.
Рубрика: Селекция, семеноводство и биотехнология сельскохозяйственных растений
Статья в выпуске: 4 (192), 2022 года.
Бесплатный доступ
Целью работы было изучение репродуктивных признаков у генотипов подсолнечника с наследственно измененным составом жирных кислот и токоферолов в семенах. Исследования проводили в 2017, 2020 и 2022 гг. в ФГБ-НУ ФНЦ ВНИИМК, г. Краснодар. Материалом был высокоолеиновый гибрид подсолнечника Окси с измененным составом токоферолов в масле семян, а также его материнская (ВК 876) и отцовская (ВК 195) родительские линии. Изучены также 19 образцов генетической коллекции. Оценка пчёлопосещаемости показала значительную наследственную изменчивость - от 98 до 0,5 особи/корзинку/час. При этом у гибрида Окси уровень пчёлопосещаемости был низким в среднем 14 особей/корзинку/час. Линии с максимальными значениями пчёлопосещаемости МВГ-8 и К 2479 имели большее количество нектара и высокий уровень его сахаристости, чем генотипы с минимальными значениями пчёлопосещаемости. Полученные данные подтверждают главную роль нектара трубчатых цветков в аттрактивности соцветия к опылителям. Гибрид Окси обладал низким значением диаметра пыльцевых зёрен (29 мкм), высокой долей дефектных пыльцевых зёрен (37 %), низким уровнем автофертильности (19 %) и завязываемости семянок (47 %). Передача этих негативных признаков гибриду происходит от материнской линии ВК 876, а не от отцовской формы ВК 195, которая характеризуется нормальными репродуктивными признаками. Содержание олеиновой кислоты достоверно не коррелировало ни с одним из семи репродуктивных признаков. Установлены достоверные коэффициенты корреляции: количество нектара -пчёлопосещаемость (0,57), количество нектара -сахаристость нектара (0,55), количество нектара -количество дефектных пыльцевых зёрен (-0,55), количество дефектных пыльцевых зёрен -автофертильность (-0,72), количество дефектных пыльцевых зёрен - завязываемость (-0,47) и автофертильность - завязываемость (0,60). Мутации высокоолеиновости Ol и состава токоферолов tphl и tph2 не обладали плейотропным действием на репродуктивные признаки у образцов генетической коллекции.
Подсолнечник, пчёлопосещаемость, олеиновая кислота, токоферолы, фертильность пыльцы, завязываемость семян
Короткий адрес: https://sciup.org/142236130
IDR: 142236130 | УДК: 633.854.78:575 | DOI: 10.25230/2412-608X-2022-4-192-3-11
Variability of reproductive traits in sunflower genotypes with different content of fatty acids and tocopherols in seeds
The aim of the work was to study the reproductive characteristics of sunflower genotypes with altered composition of fatty acids and tocopherols in seeds. Studies were conducted in 2017, 2020 and 2022 at the VNIIMK, Krasnodar. The material was a high oleic hybrid of sunflower Oxy with a modified composition of tocopherols, as well as its maternal VK 876 and male VK 195 parent lines. Moreover 19 samples of the genetic collection were studied. The assessment of bee attendance showed significant hereditary variability from 98 to 0.5 bee/plant/hour. The Oxy confirmed a low level of bee attendance, on average, 14 bee/plant/hour. The lines with the maximum values of bee attendance MVG-8 and K 2479 showed a greater amount of nectar and a high level of its sugar content than the genotypes with the minimum values of bee attendance. The data obtained confirm the main role of the nectar of tubular flowers in the pollinator attractiveness. The Oxy had a low pollen grain diameter of 29 microns, a high proportion of defective pollen grains of 37%, a low level of self-fertility of 19% and the seed setting of 47%. The transmission of these negative traits to the hybrid comes from the maternal line of VK 876, and not from the male form of VK 195, which is characterized by normal reproductive traits. The content of oleic acid did not significantly correlate with any of the seven reproductive traits. Correlation coefficients were established: the amount of nectar - bee attendance (0.57), the amount of nectar - sugar content of nectar (0.55), the amount of nectar - the number of defective pollen grains (-0.55), the number of defective pollen grains - self-fertility (-0.72), the number of defective pollen grains - seed settig (-0.47) and a self-fertility -seed setting (0.60). Mutations of high oleic Ol and the composition of tocopherols tphl and tph2 did not have a pleiotropic effect on reproductive traits in the lines of the genetic collection.
Текст научной статьи Изменчивость репродуктивных признаков у генотипов подсолнечника с различным содержанием жирных кислот и токоферолов в семенах
Acknowledgements. The research was conducted with the financial support of the Kuban Science Foundation as part of the scientific project № N-21.1/8.
Введение. Современная мировая тенденция в селекции подсолнечника как факультативно перекрестноопыляемого энтомофильного растения связана с необходимостью повышения завязываемости семян в условиях значительного снижения по техногенным причинам количества насекомых-опылителей. Для этого следует генетически повышать как автофертильность [1], так и аттрактивность цветущих растений к опылителям [2].
Сравнительная автофертильность сортов, инбредных линий и гибридов подсолнечника оценивается в 7, 37 и 43 % соответственно [3]. По другим данным, сорта показывают размах автофертильности 25–64 %, а гибриды – 53–97 % [4]. Несмотря на высокий процент автофертильности современных линий и гибридов, насекомые-опылители играют незаменимую роль на участках размножения материнских линий и участках гибридизации, где происходит перенос пыльцы от фертильного опылителя к материнской стерильной форме. Кроме того, при достаточном опылении насекомые не только повышают урожайность подсолнечника, но и улучшают качество семян [5; 6].
Нектар является материальной основой экологического мутуализма, при котором опылители, потребляя этот продукт, совершают перенос пыльцы и опыление [7]. Интенсивность посещения пчёлами корзинок подсолнечника напрямую зависит от нектаропродуктивности растений [8], которая определяется количеством нектара в трубчатых цветках. Этот признак обладает широкими пределами изменчивости. Количество нектара, выделяемое одним цветком подсолнечника, изменяется от 0,11 до 0,25 мг/цветок [9]. Пчёлы предпочитают собирать нектар с концентрацией сахара не менее 40–50 % [10]. Признак нектаропродуктивности подсолнечника характеризуется высокой наследуемостью и может быть улучшен путем селекции [11].
Важным селекционным признаком является мужская фертильность растений подсолнечника. Соцветие принято считать фертильным, если пыльники трубчатых цветков нормально развиты и в них находится пыльца [12]. Однако были по- лучены результаты, доказывающие, что растения подсолнечника могут иметь в развитых пыльниках до 50 % ненормальных, со значительными отклонениями по размерам пыльцевых зёрен [13]. Генотипы подсолнечника, имеющие морфологическую гетерогенность пыльцы, могут обладать низкой завязываемостью семянок. Поэтому в селекции подсолнечника необходимо проводить направленный отбор в сторону улучшения признака мужской фертильности.
С другой стороны, перспективным направлением в генетике и селекции подсолнечника является изучение признака высокоолеиновости и создание гибридов с данным типом масла, которое по жирнокислотному составу аналогично оливковому. Такое масло целесообразно использовать в пищевых целях для снижения крайне токсичных продуктов окисления липидов. Кроме того, для повышения окислительной стабильности масла используется также увеличение содержания в нем сильных антиоксидантов, например γ- и δ-токоферолов. Испанские учёные установили с использованием почти-изогенных гибридов подсолнечника негативное влияние мутации высокоолеино-вости Ol на автофертильность растений в некоторых генотипических средах [14].
В ходе наших предыдущих исследований обнаружен факт достоверного снижения значений ключевых репродуктивных признаков (количества и качества нектара, пчёлопосещаемости, завязывае-мости семян при свободном цветении, автофертильности, однородности и фертильности пыльцы) созданного впервые в мире во ВНИИМК высокоолеинового гибрида подсолнечника Окси с измененным составом токоферолов в масле семян (мутации tph1 и tph2 ) [15; 16; 17].
Для ответа на вопрос о возможной плейтропии вышеописанных мутаций на репродуктивные признаки следует провести исследование с использованием родительских линий гибрида Окси и линий генетической коллекции с различным со- четанием мутаций Ol, tph1 и tph2, включая почти-изогенные линии.
Целью данной работы было изучение комплекса репродуктивных признаков у генотипов подсолнечника с наследственно измененным составом жирных кислот и токоферолов в семенах для разработки генетических основ селекции на повышение завязываемости семян в условиях дефицита насекомых-опылителей.
Материалы и методы . Экспериментальные исследования проводили на опытных полях 1-го (2017 и 2020 гг.), а также 2-го (2022 г.) отделений ФГБНУ ФНЦ ВНИИМК, г. Краснодар. Материалом для изучения был высокоолеиновый гибрид подсолнечника Окси с измененным составом токоферолов в масле семян, а также его материнская (ВК 876) и отцовская (ВК 195) родительские линии. Для получения семян F 1 гибрида Окси были выполнены скрещивания родительских линий в осенне-зимний период 2021–2022 гг. в условиях фитотронно-тепличного комплекса ВНИИИМК. Кроме того, в работе изучены селекционные линии и образцы генетической коллекции подсолнечника, различающиеся по содержанию олеиновой кислоты и составу токоферолов в семенах. В частности, использовали селекционные линии: ВК 905 Ol , ВК 934 Ol , ВК1-клп Ol , ВК 101 Ol , tph2 , ВК 678 tph1 и ВК 101. Образцы генетической коллекции представлены линиями: ЛД 102, КГ 49, Л 2138, Л 7247, И 7 246, К 1587, МВГ-8, К 2479, ВК 416, ЛГ 26, RIL 39, RIL 200 и I 5 Крупняк.
Растения подсолнечника выращивали в 2017 г. на однорядных, а в 2020 и 2022 гг. на четырёхрядных делянках при схеме посева 70 × 35 см по 25 растений в ряду.
Определение жирно-кислотного состава проводили в средних пробах семян на хроматографе «Хроматэк-Кристалл 5000». Общий состав токоферолов определяли методом тонкослойной хроматографии в средней пробе.
В работе изучено семь репродуктивных признаков: пчёлопосещаемость, ко- личество нектара, сахаристость нектара, диаметр пыльцевых зёрен, количество дефектных пыльцевых зёрен, завязывае-мость семянок и автофертильность.
Для определения пчёлопосещаемости использовали маршрутный метод. Учёт проводили на растениях второго-пятого дня цветения. Опыт выполняли в трёхкратной повторности. На четырёхрядных делянках учёт проводили на 50 корзинках двух центральных рядов, затем делали перерасчёт количества особей на корзинке в час:
Х2 = Х1 × 0,6, где Х1 – общее число пчёл на 50 растениях за 2 минуты;
Х 2 – особь/корзинка/час.
Пчёлопосещаемость на однорядных делянках изучали на 10 растениях, затем делали перерасчёт признака:
Х2 = Х1 × 12, где Х1 – общее число пчёл на 10 растениях за 30 секунд;
Х 2 – особь/корзинка/час.
Для оценки нектаропродуктивности за день до отбора нектара изолировали по пять корзинок каждого генотипа третьего дня цветения индивидуальными изоляторами для предотвращения сбора нектара насекомыми, которые могут посещать растения подсолнечника в ранние утренние часы.
На следующий день срезали подготовленные корзинки, ставили в ёмкость с водой и доставляли в лабораторию для дальнейшего исследования. Нектар отбирали с 40 трубчатых цветков в пыльниковой фазе каждой из пяти корзинок (200 цветков на генотип) одного дня цветения с помощью микрокапиллярных трубок с внутренним диаметром 0,25 мм и внешним – 0,50 мм.
После отбора нектара в микрокапил-лярную трубку его помещали на предварительно взвешенный диск фильтровальной бумаги и проводили повторное взвешивание. По разнице массы находили количество нектара. После высыхания диска, т.е. испарения водной части нектара, проводили повторное взвешивание. По разнице массы вычисляли долю сухого вещества (сахаристость) в нектаре.
Собранную пыльцу окрашивали ацето-орсеином в течение пяти минут, затем препарат микроскопировали с помощью цифрового микроскопа Биолаб TS-2000 LCD МС 20. Проводили измерение 200 пыльцевых зёрен и описывали морфологическую однородность пыльцы как показатель нормальной фертильности.
После созревания корзинки подсолнечника срезали и в лабораторных условиях индивидуально обмолачивали. Завязываемость семянок при свободном цветении и самоопылении (автофертильность) определяли как отношение выполненных семянок к общему числу семянок в корзинке (или количеству трубчатых цветков). Для подсчёта выполненных семянок использовали автоматический счётчик Automatic Seed Counter.
Результаты и обсуждение . Двухлетнее изучение пчёлопосещаемости 11 генотипов подсолнечника показало значительную наследственную изменчивость признака от 98 у К 2479 до 0,5 особи/корзинку/час у карликового И 7 246, т.е. почти в 200 раз (табл. 1). У гибрида Окси подтвердился низкий уровень пчёлопосещаемости – в среднем 14 особей/корзинку/час. Две другие вы-сокоолеиновые линии – ЛГ 26 и ВК 905 – также относились к этой низкозначимой категории. При этом ВК 905 обладает длинным венчиком трубчатых цветков, что, очевидно, может негативно влиять на доступность нектарников для опылителей.
Таблица 1
Пчёлопосещаемость растений подсолнечника (особь/корзинка/час)
ЦЭБ ВНИИМК, 1-е и 2-е отделение, г. Краснодар, 2017, 2022 гг.
|
Генотип |
Год |
Среднее за два года |
|
|
2017 |
2022 |
||
|
К 2479 |
160 |
36 |
98 |
|
МВГ-8 |
132 |
34 |
83 |
|
Л 2138 |
36 |
10 |
23 |
|
КГ 49 |
24 |
7 |
16 |
|
ВК 416 |
20 |
15 |
18 |
|
ЛД 102 |
20 |
15 |
18 |
|
ЛГ 26 Ol |
12 |
8 |
10 |
|
Окси Ol, tph1, tph2 |
10 |
18 |
14 |
|
ВК 905 Ol |
6 |
8 |
7 |
|
Л 7247 |
4 |
1 |
3 |
|
И 7 246 |
0 |
1 |
0,5 |
|
НСР 05 |
6 |
4 |
- |
Две отобранные линии с максимальными значениями пчёлопосещаемости – МВГ-8 и К 2479 – ожидаемо имели за два года большее количество нектара (0,27 и 0,24 мг/цветок) и высокий уровень его сахаристости (55 и 47 % соответственно), чем три линии с минимальными значениями пчёлопосещаемости – И 7 246, Л
7247 и Окси (табл. 2). При этом для изученных генотипов наблюдали положительную связь количества нектара и его сахаристости. Полученные данные подтверждают главную роль нектара трубчатых цветков в аттрактивности соцветия опылителям.
Таблица 2
Характеристика нектара трубчатых цветков у генотипов подсолнечника
ЦЭБ ВНИИМК, 1-е и 2-е отделение, г. Краснодар, 2020, 2022 гг.
|
Генотип |
Количество нектара, мг/цветок |
Сахаристость нектара, % |
||||
|
2020 г. |
2022 г. |
среднее за два года |
2020 г. |
2022 г. |
среднее за два года |
|
|
МВГ-8 |
0,32 |
0,22 |
0,27 |
53 |
56 |
55 |
|
К 2479 |
0,25 |
0,23 |
0,24 |
40 |
53 |
47 |
|
И 7 246 |
0,22 |
0,09 |
0,16 |
46 |
45 |
46 |
|
Л 7247 |
0,15 |
0,12 |
0,14 |
40 |
24 |
32 |
|
Окси Ol, tph1, tph2 |
0,14 |
0,17 |
0,16 |
21 |
33 |
27 |
|
НСР 05 |
0,03 |
0,04 |
- |
6 |
7 |
- |
В 2022 г. 22 генотипа распределились по трём группам на основе содержания олеиновой кислоты в масле семян: высо- кое (80–90 %), среднее (50–63 %) и низкое (27–39 %) (рис. 1, табл. 3). Высоко-олеиновый фенотипический класс вызывается гомозиготным состоянием мутации Ol, за исключением низкоолеи-новой линии RIL 200, у которой мутация Ol находится под эпистатическим действием супрессора Sup и фенопически не проявляется (табл. 3).
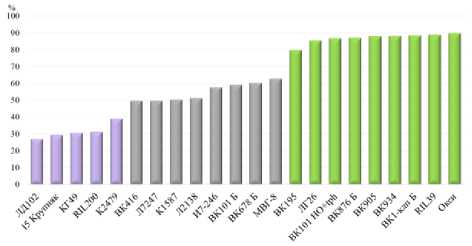
Рисунок 1 – Содержание олеиновой кислоты в масле семян подсолнечника (ЦЭБ ВНИИМК, 2-е отделение, г. Краснодар, 2022 г.)
Пять генотипов коллекции обладали рецессивными мутациями tph1 и tph2 состава токоферолов в семенах (табл. 3, рис. 2). Для гибрида Окси, его родительских линий ВК 876 и ВК 195 характерно гомозиготное состояние по двум мутациям, приводящим к высокому содержанию γ- и δ-токоферолов. Мутация tph1 приводит к соразмерному накоплению α- и β-форм, а у мутации tph2 синтезируется преимущественно γ-форма. Дикий тип линии ВК 101 связан с присутствием в фенотипе только α-токоферола.
Таблица 3
Состав токоферолов в семенах у генотипов подсолнечника
ЦЭБ ВНИИМК, 2-е отделение, г. Краснодар, 2022 г.
|
Генотип |
Состав токо |
еролов, % |
||
|
α |
β |
γ |
δ |
|
|
Окси Ol, tph1, tph2 |
0 |
0 |
70,5 |
29,5 |
|
ВК 876 Ol, tph1, tph2 |
0 |
0 |
66,0 |
33,0 |
|
ВК 195 Ol, tph1, tph2 |
0 |
0 |
63,1 |
36,9 |
|
ВК 101 Ol, tph2 |
12,6 |
0 |
83,3 |
0 |
|
ВК 678 tph1 |
45,3 |
54,7 |
0 |
0 |
|
ВК 101 |
100 |
0 |
0 |
0 |
|
НСР 05 |
4,5 |
4,0 |
4,2 |
4,1 |
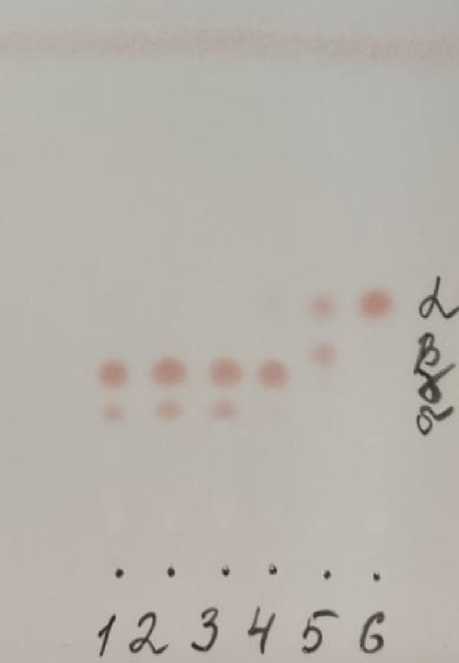
Рисунок 2 – Хроматограмма состава токоферолов в масле семян подсолнечника (ВНИИМК, г. Краснодар, 2022 г.):
1 – Окси Ol, tph1, tph2; 2 – ВК 876 Ol, tph1, tph2; 3 – ВК 195 Ol, tph1, tph2;
4 – ВК 101 Ol, tph2; 5 – ВК 678 tph1;
6 – ВК 101 дикий тип
Гибрид Окси обладал самым низким значением диаметра пыльцевых зёрен (29 мкм), одним из высоких значений доли дефектных пыльцевых зёрен (37 %), одним из низких значений автофертильности (19 %) и завязываемости семянок при свободном цветении (47 %) (табл. 4). При этом очевидна передача этих негативных признаков от материнской линии ВК 876 Ol, tph1, tph2 , а не от отцовской формы ВК 195 Ol, tph1, tph2 , которая характеризуется нормальными репродуктивными признаками при наличии всех 8
трёх мутаций высокоолеиновости и состава токоферолов в семенах.
Содержание олеиновой кислоты достоверно не коррелировало ни с одним из семи репродуктивных признаков (табл. 5). Однако установлены достоверные коэффициенты корреляции для шести пар репродуктивных признаков между собой: количество нектара – пчё-лопосещаемость (0,57), количество нектара – сахаристость нектара (0,55), количество нектара – количество дефектных пыльцевых зёрен (-0,55), количество дефектных пыльцевых зёрен – автофертильность (-0,72), количество дефектных пыльцевых зёрен – завязываемость (-0,47) и автофертильность – завязываемость (0,60).
Заключение. Изучен комплекс репродуктивных признаков у генотипов подсолнечника с наследственно измененным составом жирных кислот и токоферолов в семенах с целью обоснования селекции на повышение завязываемости семян при потенциальной нехватке насекомых-опылителей. Мутации качества масла семян подсолнечника: высокоолеиновости Ol и состава токоферолов tph1 и tph2 не обладали плейотропным действием на семь изученных репродуктивных признаков у образцов генетической коллекции. Этот факт позволяет прогнозировать эффективную селекционную работу по созданию продуктивных гибридов подсолнечника с улучшенным качеством масла.
Таблица 4
Характеристика репродуктивных признаков у генотипов подсолнечника
ЦЭБ ВНИИМК, 2-е отделение, г. Краснодар, 2022 г.
|
Генотип |
Содержание олеиновой кислоты, % |
Пчёлопосе-щаемость, особь/корзинка/час |
Количество нектара, мкг/цве-ток |
Сахаристость, % |
Диаметр пыльцевых зёрен, мкм |
Количество дефектных пыль-це-вых зёрен, % |
Автофертильность, % |
Завязывае-мость, % |
|
Окси Ol, tph1, tph2 |
89,8 |
18 |
0,17 |
33 |
29 |
37 |
19 |
47 |
|
RIL 39 Ol |
88,8 |
24 |
0,22 |
86 |
33 |
18 |
40 |
51 |
|
ВК1-клп Ol |
88,4 |
12 |
0,13 |
39 |
32 |
14 |
61 |
74 |
|
ВК 934 Ol |
88,1 |
10 |
0,25 |
68 |
32 |
10 |
40 |
58 |
|
ВК 905 Ol |
88,0 |
8 |
0,19 |
73 |
33 |
6 |
50 |
64 |
|
ВК 876 Ol, tph1, tph2 |
87,0 |
8 |
0,05 |
27 |
31 |
30 |
24 |
40 |
|
ВК 101 Ol, tph2 |
86,9 |
9 |
0,20 |
37 |
31 |
15 |
42 |
62 |
|
ЛГ 26 Ol |
85,4 |
8 |
0,12 |
51 |
32 |
16 |
33 |
70 |
|
ВК 195 Ol, tph1, tph2 |
79,6 |
7 |
0,14 |
59 |
32 |
6 |
56 |
63 |
|
МВГ-8 |
62,7 |
34 |
0,22 |
56 |
31 |
9 |
59 |
72 |
|
ВК 678 tph1 |
60,1 |
19 |
0,16 |
47 |
31 |
29 |
38 |
66 |
|
ВК 101 |
59,1 |
22 |
0,18 |
31 |
32 |
12 |
33 |
64 |
|
И 7 -246 |
57,6 |
1 |
0,09 |
45 |
31 |
17 |
18 |
61 |
|
Л2 138 |
51,1 |
10 |
0,19 |
15 |
32 |
26 |
5 |
44 |
|
К 1587 |
50,3 |
20 |
0,21 |
71 |
32 |
8 |
30 |
64 |
|
Л 7247 |
49,7 |
1 |
0,12 |
24 |
33 |
20 |
29 |
49 |
|
ВК 416 |
49,6 |
15 |
0,12 |
59 |
32 |
45 |
8 |
61 |
|
К 2479 |
38,9 |
36 |
0,23 |
53 |
33 |
9 |
36 |
64 |
|
RIL 200 Ol Sup |
31,0 |
13 |
0,15 |
66 |
31 |
11 |
46 |
52 |
|
КГ 49 |
30,6 |
7 |
0,10 |
36 |
30 |
45 |
13 |
48 |
|
I 5 Крупняк |
29,2 |
14 |
0,07 |
- |
31 |
24 |
38 |
68 |
|
ЛД 102 |
26,9 |
15 |
0,19 |
64 |
33 |
14 |
31 |
58 |
|
НСР 05 |
4 |
0,04 |
7 |
1 |
4 |
8 |
9 |
Таблица 5
Корреляция содержания олеиновой кислоты в масле семян с репродуктивными признаками подсолнечника (n=22)
ЦЭБ ВНИИМК, 2-е отделение, г. Краснодар, 2022 г.
|
Показатель |
1 |
2 |
3 |
4 |
5 |
6 |
7 |
|
Содержание олеиновой кислоты (1) |
|||||||
|
Пчёлопосещаемость (2) |
-0,14 |
||||||
|
Количество нектара (3) |
0,15 |
0,57* |
|||||
|
Сахаристость нектара (4) |
0,05 |
0,30 |
0,55* |
||||
|
Диаметр пыльцевых зёрен (5) |
-0,03 |
0,09 |
0,35 |
0,40 |
|||
|
Количество дефектных ПЗ (6) |
-0,17 |
-0,18 |
-0,50* |
-0,41 |
0,18 |
||
|
Автофертильность (7) |
0,32 |
0,25 |
0,29 |
0,34 |
0,23 |
-0,72* |
|
|
Завязываемость (8) |
0,05 |
0,31 |
0,17 |
0,18 |
0,20 |
-0,47* |
0,60* |
* – достоверно на 5%-ном уровне значимости
Список литературы Изменчивость репродуктивных признаков у генотипов подсолнечника с различным содержанием жирных кислот и токоферолов в семенах
- Skoric D. Sunflower breeding // Sunflower Genetics and Breeding. International monograph. -Serbian Academy of Science and Arts, Branch in Novi Sad, 2012. - 520 p.
- Miklw V., Sakaс Z., Dusanw N., Atlagic J., Joksimovic J., Vasic D. Effects of genotype, growing conditions, and several parameters of sunflower attractiveness for bee visitation // Proc. of the 16th International Sunflower Conference, Fargo, ND USA. - 2014. - P. 871-876.
- Бочковой А.Д., Камардин В.А., Назаров Д.А. О перспективах отбора самофертильных биотипов в звеньях первичного семеноводства сортов подсолнечника // Масличные культуры. - 2020. - Вып. 1 (181). - С. 3-11.
- Горьковая Е.Г. Влияние автофертильности исходного материала на завязываемость в первых поколениях инбредных линий // Материалы VI международной конференции молодых учёных и специалистов «Инновационные направления исследований в селекции и технологии возделывания масличных культур», 24-25 февраля 2011 г. - Краснодар, 2011. - С. 61-65.
- Wali M., Shafqat S., Ammad A., Ishfaq M.Y., Anjum M.M. Pollinator community of sunflower (Helianthus annus L.) and its role in crop reproductive success // Asian J. Agric. Biol. - 2021. - Vol 2. - 202007398. DOI: 10.35495/ajab.2020.07.398.
- Сазоненко М.М., Баладин В.С., Нагоев А.Ш., Казакова В.В. Влияние опыления пчёл на урожайность подсолнечника гибрида Натали // Научные исследования XXI века: -Научно-издательский центр «Мир науки», 2019. - № 2. - С. 143-145.
- Bergonzoli S., Romano E., Beni C., Latterini F., Lo Scalzo R., Scarfone A. Nectar dynamics and pollinators preference in sunflower // Insects. - 2022. - No 13 (8). 717. DOI: 10.3390/ insects13080717.
- Голиков В.И., Морева Л.Я. Пчелиные -опылители сортов, линий и гибридов подсолнечника // Пчеловодство. - 2019. - № 7. - С. 26-27.
- Rinku Chaudhary O.P., Kaushik H.D. Variations in morphological and phonological traits of selected sunflower populations and hybrids reveal their relative preference to honey bees // Indian Journal of Ecology. - 2017. - Vol. 44 (Special, 5). - P. 536-542.
- Пономарева Е.Г. Кормовая база пчеловодства и опыление сельскохозяйственных растений. 2-е изд., перераб. и доп. - М.: Колос, 1973. - 256 с.
- Sammataro D., Erickson E.N., Garmend M.B. Ultrastructure of the sunflower nectary // Journal of Apicultural Research. - 1985 . - Vol. 24 (3). - P. 150-160.
- Воронова О.Н., Гаврилова В.А. Количественный и качественный анализ пыльцы подсолнечника (Helianthus L.) и его использование в селекционной работе // Труды по прикладной ботанике, генетик и селекции. - 2019. - Вып. 180 (1). - С. 95-104.
- Воронова О.Н., Толстая Т.Т., Рожкова В.Т., Гаврилова В.А. Определение фертильности пыльцы у ряда диких многолетних видов и образцов подсолнечника коллекции, произрастающей на Кубанской станции ВИР // Труды по прикладной ботанике, генетике и селекции. - 2011. - Т. 167. - С. 145-158.
- Fernandez-Martinez J., Munoz J., Gomez-Arnau J. Performance of near-isogenic high and low oleic acid hybrids of sunflower // Crop Science. - 1993. - Vol. 33 (6). - P. 1158-1163.
- Рубанова О.А., Демурин Я.Н. Влияние нектаропродуктивности на посещаемость пчелами гибридов подсолнечника // Пчеловодство. - 2021. - № 5. - С. 24-26.
- Демурин Я.Н., Рубанова О.А. Завязываемость семян у гибридов подсолнечника в конкурсном сортоиспытании // Труды КубГАУ. - 2019. - № 5 (80). - С. 109113. DOI: 10.21515/1999-1703-80-109-113.
- Демурин Я.Н., Рубанова О.А. Пыльцевой анализ растений различных генотипов подсолнечника // Масличные культуры. - 2021. - Вып. 2 (186). - С. 10-17. DOI: 10.25230/2412-608X-2021-2-186-10-17.
