Изменение показателей генетической структуры популяции сортов овса, возделываемых в Тюменской области, за 90-летний период

Автор: Любимова Анна Валерьевна, Еремин Дмитрий Иванович, Кочнева Дарья Алексеевна
Журнал: Вестник Красноярского государственного аграрного университета @vestnik-kgau
Рубрика: Агрономия
Статья в выпуске: 12, 2022 года.
Бесплатный доступ
Изучение показателей генетической структуры популяции и их изменений с течением времени важно для повышения эффективности селекционного процесса и грамотного управления развитием популяций сортов. Высокую эффективность в таких исследованиях показали проламин-кодирущие локусы. Целью исследований была оценка изменений во времени показателей генетической структуры популяций сортов овса посевного, включенных в Государственный реестр селекционных достижений по Тюменской области, с 1929 по 2021 г. с использованием множественных аллелей авенин-кодирующих локусов. Материалом для исследования послужили 19 сортов овса посевного. По локусу Avn A выявлено 8 различных аллелей, по локусу Avn B - 7 и по локусу Avn C - 6. В исследованных группах сортов по локусу Avn A максимальную частоту встречаемости имели аллели A2 и A11, по локусу Avn B - B8 и B4, а по локусу Avn C - C2 и Cnew. Аллельные варианты A1, A5, Anew, B1, Bnew1, Bnew2 и C1 имели низкую частоту встречаемости и элиминировались из популяции в течение нескольких десятков лет. Наиболее стабильные во времени значения внутрипопуляционного разнообразия и доли редких морф характерны для локуса Avn A. Значение внутрипопуляционного разнообразия по локусам Avn B и Avn C волнообразно изменялось, при этом периоды снижения величины данного показателя по одному из локусов сопровождались его повышением по другому. Внутрипопуляционное разнообразие сортов овса в настоящее время характеризуется как высокое. Показатель доли редких морф имеет низкое значение (0,04-0,09), что указывает на стабильность анализируемой популяции. Аллели локусов Avn B и Avn C перспективны для изучения в качестве маркеров ассоциаций генов, определяющих адаптивно-значимые признаки, а локуса Avn B - хозяйственно ценные признаки в условиях Тюменской области.
Овес, электрофорез, авенин, проламин-кодирующие локусы, генетическое разнообразие, доля редких морф, генетическая структура популяции, генетическая эрозия, маркер-ориентированная селекция
Короткий адрес: https://sciup.org/140297197
IDR: 140297197 | УДК: 581.19:631.52:633.112.1 | DOI: 10.36718/1819-4036-2022-12-32-41
Changes in indicators of the oat varieties population genetic structure cultivated in the Tyumen region over a 90-year period
The study of indicators of the genetic structure of the population and their changes over time is important for improving the efficiency of the breeding process and competent management of the development of populations of varieties. High efficiency in such studies was shown by prolamin-coding loci. The aim of research was to assess changes over time in the indicators of the genetic structure of the populations of oat varieties included in the State Register of Breeding Achievements in the Tyumen Region from 1929 to 2021 using multiple alleles of avenin-coding loci. The material for the study was 19 varieties of oats. Eight different alleles were identified for the Avn A locus, 7 for the Avn B locus, and 6 for the Avn C locus. C2 and Cnew. Allelic variants A1, A5, Anew, B1, Bnew1, Bnew2 and C1 had a low frequency of occurrence and were eliminated from the population within several decades. The most stable values of intrapopulation diversity and the proportion of rare morphs over time are typical for the Avn A locus. The value of intrapopulation diversity at the Avn B and Avn C loci changed in waves, while periods of a decrease in the value of this indicator for one of the loci were accompanied by its increase for another. The intrapopulation diversity of oat varieties is currently characterized as high. The indicator of the proportion of rare morphs has a low value (0.04-0.09), which indicates the stability of the analyzed population. The alleles of the Avn B and Avn C loci are promising for study as markers of gene associations that determine adaptively significant traits, and the Avn B locus is economically valuable traits in the conditions of the Tyumen Region.
Текст научной статьи Изменение показателей генетической структуры популяции сортов овса, возделываемых в Тюменской области, за 90-летний период
Введение. Овес ( Avena sativa L.) – культура азиатского происхождения, используемая для производства продуктов питания человека, а также на корм животным [1, 2]. В отличие от других зерновых культур овес не требователен к почвенному плодородию и способен формировать высокие урожаи даже в условиях пониженных температур. Это делает его культурой, наиболее перспективной для возделывания в условиях Западной Сибири. С начала XX в. в Государственный реестр селекционных достижений по Тюменской области было включено 19 сортов посевного овса, из них 6 – сорта селекции НИИСХ Северного Зауралья (филиал ТюмНЦ СО РАН). Селекционная работа с этой культурой в регионе направлена на создание
сортов интенсивного типа, характеризующихся высокой урожайностью, качеством зерна, скороспелостью и устойчивостью к неблагоприятным факторам среды.
Однако активная селекционная работа может привести к снижению генетического разнообразия вида [3, 4]. Для повышения эффективности селекции и грамотного управления развитием популяций сортов важно понимание процессов, протекающих в этих популяциях, что требует изучения их генетической структуры и ее изменений с течением времени. К наиболее часто анализируемым показателям относятся внутрипопуляционное разнообразие (µ), доля редких морф (h) и среднее генное разнообразие ( Н ). Вычисление этих показателей позволяет
Таблица 1
Проанализированные сорта овса посевного
ляционного разнообразия и процессов генетической эрозии вида [8].
Цель исследований – оценка изменений во времени показателей генетической структуры популяций сортов овса посевного, включенных в Государственный реестр селекционных достижений по Тюменской области, с использованием множественных аллелей авенин-кодирующих локусов с 1929 по 2021 г.
Материалы и методы. Исследования проводили на базе лаборатории геномных исследований в растениеводстве Тюменского научного центра СО РАН в 2022 году. Материалом для изучения послужили сорта овса посевного, включенные в Государственный реестр селекционных достижений по Тюменской области, в период с 1929 по 2021 гг. (табл. 1).
Биохимические формулы авенина, представленные в таблице 1, описаны нами в работе [8], уточнены и дополнены в процессе идентификации новых блоков компонентов проламинов [9]. Аллели авенин-кодирующих локусов определяли в соответствии с каталогом, разработанным В.А. Портянко [10], с дополнениями А.В. Любимовой [9]. В случае, если найденный вариант блока отсутствовал в каталоге, его обозначали пометкой «new».
Для того чтобы оценить динамику изменений показателей структуры популяции во времени, все сорта были разделены на группы. В одну группу объединяли сорта, возделываемые в один и тот же десятилетний период времени.
Генное разнообразие на локус (H) рассчитывали по формуле я ×(1-∑i=i Pi} , (1)
где pi – популяционная частота i -го аллеля; k – количество аллелей локуса; n – объем выборки [11]. Для расчета среднего генного разнообразия ( Н ) усредняли количество аллелей на локус по всем локусам.
Показатели внутрипопуляционного разнообразия (µ) и доли редких морф (h) рассчитывали по формулам:
µ=(р1+ р2+ р3 + ⋯ + рm)2 , (2)
һ =1- µ , (3)
где p 1 , p 2 , … p m – частоты аллелей в долях от единицы; m – число вариаций признака, выделенных в данной выборке [12].
Расчеты проводили с использованием программы Arlequin Ver 3.5.2.2 (Copyright 2015 L. Excoffier. CMPG, University of Berne) и в MS Excel.
Результаты и их обсуждение. В результате исследований установлено, что из 19 проанализированных сортов 9 были гетерогенными по компонентному составу авенина и состояли из двух биотипов: Победа, Золотой дождь, Орел, Северянин, Нарымский 943, Таежник, Сельма, Мегион и Отрада. При расчете показателей генетической структуры популяций каждый обнаруженный биотип рассматривался как отдель- ный образец. Таким образом, всего было проанализировано 28 образцов.
Ряд из исследованных образцов имели идентичные спектры авенина: Победа (I биотип) и Золотой дождь (II биотип); Победа (II биотип) и Орел (II биотип); Золотой дождь (I биотип), Нидар и Таежник; Северянин (II биотип) и Скороспелый; Нарымский 943 (II биотип), Астор и Новосибирский 88; Перона и Талисман (см. табл. 1). К совпадению электрофоретических спектров приводит множество факторов – от общности происхождения, как в случае с сортами Победа, Золотой дождь и Орел, выведенными из сорта овса Milton (=Propsteier), до отбора в процессе селекции генотипов, характеризующихся определенными адаптивными генными комплексами и маркирующими их аллелями авенин-кодирующих локусов [8].
Установлено, что по локусу Avn A в сортах овса посевного встречалось 8 различных аллелей, по локусу Avn B – 7 и по локусу Avn С – 6. Как видно из рисунка 1, частота встречаемости аллелей авенин-кодирующих локусов в популяциях сортов изменялась с течением времени. При этом за все годы по локусу Avn A максимальную частоту встречаемости имели аллели А2 и А11 , по локусу Avn B – В8 и В4, а по локусу Avn С – С2 и Сnew. По нашему мнению, такие аллели могут выступать в качестве маркеров ассоциаций генов, имеющих высокую хозяйственную или адаптивную ценность для данного региона. Кроме того, по каждому из локусов были выявлены аллели, имеющие относительно низкую частоту встречаемости и элиминировавшиеся из популяции в течение нескольких десятков лет. К таковым можно отнести варианты А1, А5, Аnew, В1, Вnew1, Вnew2 и С1 . В наших предыдущих исследованиях установлено, что смена одних аллелей АКЛ другими связана с процессом сортосмены и, в частности, замены сортов иностранного происхождения на сорта российской, а затем и региональной селекции [8]. При этом источником «новых» аллелей становится генетически отличающийся исходный материал, вовлекаемый в селекционный процесс.
От частоты встречаемости и разнообразия аллелей зависят такие показатели структуры популяции, как внутрипопуляционное разнообразие (µ) и доля редких морф (h) (табл. 2).
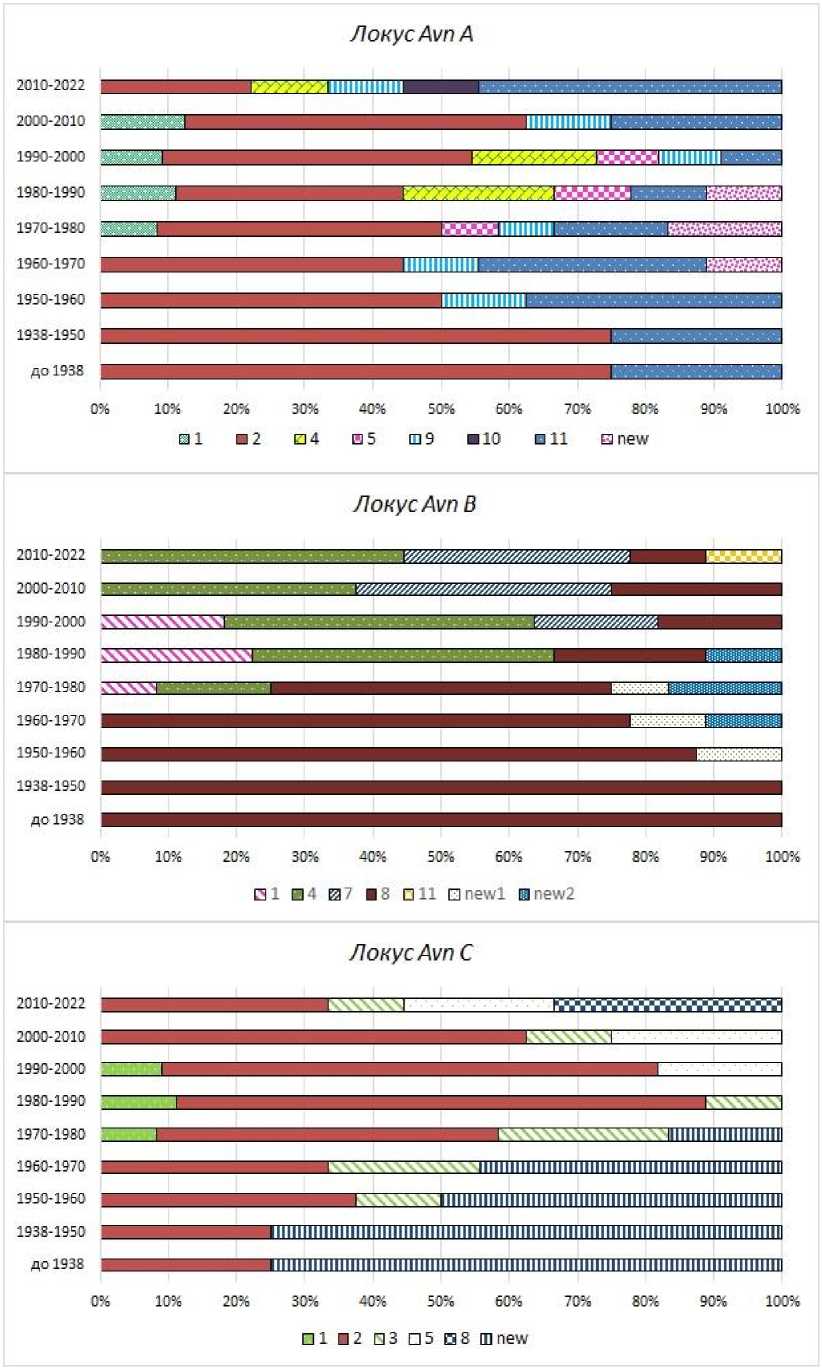
Рис. 1. Частота встречаемости аллелей авенин-кодирующих локусов у сортов овса посевного, возделываемых в Тюменской области, в период с 1929 по 2021 г., %
Таблица 2
Генное разнообразие (Н), внутрипопуляционное разнообразие (µ) и доля редких морф (h) в популяциях исследованных сортов овса посевного
|
Cl О ф “ c £ ф cE co О co |
ZE Ф Ф co о I VO ф о co 05 |
|
Ф о x Z h £ 05 i X4 CD |
X ci ^ ^§ к о с; s о d |
О g = ф с £ ф со О со |
ZE Ф Ф со о то х ю ф О 1__ о со 05 |
05 VO 05 “ m m О 05 О 05 т 1. |
ф о h £ 05 i § СП |
X ^§ СК о о d |
|
Локус Avn A |
Локус Avn B |
||||||||
|
До 1938 |
0,50 |
2 |
1,87±0,250 |
0,07 |
До 1938 |
0,00 |
1 |
1,00±0,000 |
0,00 |
|
1938–1950 |
0,50 |
2 |
1,87±0,250 |
0,07 |
1938–1950 |
0,00 |
1 |
1,00±0,000 |
0,00 |
|
1950–1960 |
0,68 |
3 |
2,80±0,265 |
0,07 |
1950–1960 |
0,25 |
2 |
1,66±0,265 |
0,17 |
|
1960–1970 |
0,75 |
4 |
3,65±0,376 |
0,09 |
1960–1970 |
0,41 |
3 |
2,40±0,400 |
0,20 |
|
1970–1980 |
0,81 |
6 |
5,42±0,512 |
0,10 |
1970–1980 |
0,74 |
5 |
4,41±0,464 |
0,12 |
|
1980–1990 |
0,88 |
6 |
5,67±0,453 |
0,05 |
1980–1990 |
0,77 |
4 |
3,77±0,308 |
0,06 |
|
1990–2000 |
0,80 |
6 |
5,32±0,573 |
0,11 |
1990–2000 |
0,76 |
4 |
3,82±0,253 |
0,05 |
|
2000–2010 |
0,75 |
4 |
3,66±0,392 |
0,08 |
2000–2010 |
0,75 |
3 |
2,97±0,097 |
0,01 |
|
2010–2022 |
0,80 |
5 |
4,57±0,467 |
0,09 |
2010–2022 |
0,75 |
4 |
3,65±0,367 |
0,09 |
|
Локус Avn C |
Среднее генное разнообразие H |
||||||||
|
До 1938 |
0,50 |
2 |
1,87±0,250 |
0,07 |
До 1938 |
0,333 |
|||
|
1938–1950 |
0,50 |
2 |
1,87±0,250 |
0,07 |
1938–1950 |
0,333 |
|||
|
1950–1960 |
0,67 |
3 |
2,80±0,265 |
0,07 |
1950–1960 |
0,535 |
|||
|
1960–1970 |
0,72 |
3 |
2,94±0,137 |
0,02 |
1960–1970 |
0,629 |
|||
|
1970–1980 |
0,71 |
4 |
3,63±0,336 |
0,09 |
1970–1980 |
0,757 |
|||
|
1980–1990 |
0,41 |
3 |
2,40±0,400 |
0,20 |
1980–1990 |
0,694 |
|||
|
1990–2000 |
0,47 |
3 |
2,50±0,337 |
0,17 |
1990–2000 |
0,678 |
|||
|
2000–2010 |
0,60 |
3 |
2,70±0,317 |
0,10 |
2000–2010 |
0,702 |
|||
|
2010–2022 |
0,80 |
4 |
3,84±0,262 |
0,04 |
2010–2022 |
0,782 |
|||
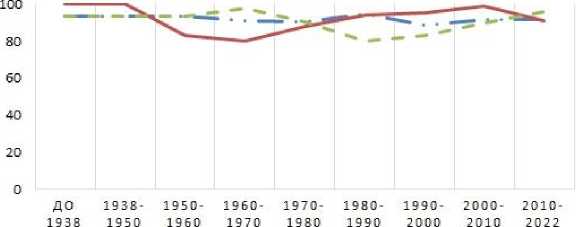
Внутрипопуляционное разнообразие (µ) – это показатель, позволяющий оценить степень раз-нокачественности особей в популяции в единицах «число морф». Чем равномернее распределение аллелей в популяции, тем ближе µ к общему числу вариаций признака в выборке (m). При одинаковой частоте всех признаков µ будет равно m. Так как число выявленных аллелей в группах сортов, возделываемых в разные периоды, отличалось, чтобы нагляднее оценить изме- нения внутрипопуляционного разнообразия во времени, его значения были пересчитаны нами в процентном отношении от общего числа вариаций признака в выборке. Это позволило сравнить значения µ в разных группах сортов. На основе показателя µ рассчитывали показатель доли редких морф (h), по которому можно судить о стабильности анализируемой популяции. По полученным данным были построены графики, представленные на рисунке 2.
ВНУТРИПОПУЛЯЦИОННОЕ РАЗНООБРАЗИЕ, %
--- ‘Avn A Avn В — — Avn С
ДОЛЯ РЕДКИХ МОРФ
• Avn A Дул В — — Avn С
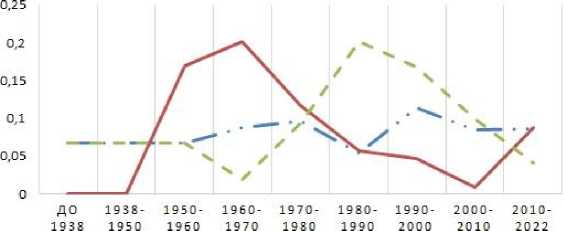
Рис. 2. Внутрипопуляционное разнообразие (µ), % и доля редких морф (h) в популяции сортов овса посевного с 1929 по 2021 г.
Как видно из рисунка 2 и таблицы 2, наиболее стабильные во времени значения µ и h для популяций характерны по локусу Avn А. Это связано с тем, что во всех проанализированных группах сортов, независимо от происхождения и периода возделывания, преобладали два аллеля по данному локусу – А2 и А11. Вероятно, эти два аллельных варианта сцеплены с генами или генными ассоциациями, дающими особые преимущества несущим их генотипам в условиях Тюменской области. Так как данные аллели характерны не только для сортов местной селекции, но и образцов иностранного происхождения, по нашему мнению, их встречаемость в большей степени связана с маркированием хозяйственно ценных, а не адаптивно-значимых признаков. При этом значения генного разнообразия (Н) по локусу Avn А во многих группах сортов имеют более высокие значения, чем по локусам Avn В и Avn С. Это связано с тем, что в изученных популяциях данный локус характеризуется максимальным разнообразием выявленных аллелей, многие из которых имеют частоту встречаемости не более 12 % и в процессе сортосмены элиминируются из популяции. Многие авторы отме- чают снижение величины среднего генного разнообразия в популяциях современных сортов и связывают это с введением в селекционный процесс одних и тех же генотипов [13, 14]. Величина этого показателя у популяций сортов мягкой пшеницы европейской селекции составляет 0,62–0,76 [15], украинской селекции – 0,5–0,6 [16], а для сортов твердой пшеницы – 0,42–0,64 [13].
Показатель внутрипопуляционного разнообразия по локусам Avn В и Avn С в исследуемый период времени волнообразно изменялся. Например, по локусу Avn В отмечено его снижение со 100,0 до 79,9 % (от величины m) в 1929– 1970 гг., а затем возрастание до 99,1 % к 2010 г. Такие же изменения отмечены и для локуса Avn С. Периоды снижения величины µ по одному из этих локусов сопровождаются его повышением по другому. Например, с 1960 по 1990 г. отмечается снижение внутрипопуляционного разнообразия по локусу Avn С, но одновременное его повышение по локусу Avn В. При этом низким значениям внутрипопуляционного разнообразия всегда соответствуют высокие значения доли редких морф. И.Е. Трофимов (2008) отмечает, что значение h возрастает в неблагоприятных условиях при снижении генотипического разнообразия. Частое проявление редких признаков в популяции является отражением ее адаптивных способностей к неблагоприятным условиям среды [17]. Это подтверждают П.Ю. Волкова, С.А. Гераськин (2012), выявившие возрастание биохимического полиморфизма в популяциях сосны обыкновенной, подвергшейся хроническому облучению, а также Е.В. Ульянова и В.Н. По-золотина (2006), оценившие редкие фены в це-нопопуляциях одуванчика. Причина данного явления в том, что в результате сильного влияния отрицательных факторов на популяцию происходит гибель наиболее чувствительных особей. Это ведет к снижению внутрипопуляционоого разнообразия. При этом происходит увеличение генетической изменчивости, что является реакцией популяции на стрессовые воздействия [18, 19]. Выявленные нами изменения величин µ и h хорошо согласуются с этим утверждением. Вероятно, аллели локусов Avn В и Avn С маркируют ассоциации генов, определяющие адаптивно-значимые признаки генотипов. При этом в разные периоды времени важность этих ассоциаций для отбора генотипов в селекционном процессе меняется.
В целом внутрипопуляционное разнообразие по всем трем авенин-кодирующим локусам в настоящее время можно охарактеризовать как высокое. Показатель доли редких морф имеет низкое значение (0,04–0,09), что свидетельствует о стабильности популяции сортов, возделываемых в Тюменской области. Значение среднего генетического разнообразия ( Н ), начиная с 1970-х гг., характеризуется как высокое, и с 1980 г. по настоящее время его величина возрастает. Это свидетельствует об отсутствии генетической эрозии в популяции сортов овса, возделываемых в Тюменской области.
Заключение
-
1. Наиболее стабильные во времени значения внутрипопуляционного разнообразия и доли редких морф для популяций характерны по локусу Avn А. Показатель внутрипопуляционного разнообразия по локусам Avn В и Avn С в период с 1929 по 2021 г. волнообразно изменялся, при этом периоды снижения величины µ по одному из этих локусов сопровождаются его повышением по другому локусу.
-
2. Внутрипопуляционное разнообразие районированных сортов овса в настоящее время характеризуется как высокое, а показатель доли
редких морф имеет низкое значение (0,04–0,09), что свидетельствует о стабильности популяции. Среднее генное разнообразие с 1980 г. по настоящее время возрастает и оценивается как высокое.
-
3. Аллели локусов Avn В и Avn С перспективны для изучения в качестве маркеров ассоциаций генов, определяющих адаптивно-значимые, а локуса Avn В – хозяйственно ценные признаки генотипов в условиях Тюменской области.
Список литературы Изменение показателей генетической структуры популяции сортов овса, возделываемых в Тюменской области, за 90-летний период
- Translating insights from the seed metabolome into improved prediction for lipid-composition traits in oat (Avena sativa L.) / M.T. Campbell [et al.] // Genetics. 2021. 217(3). DOI: 10.1093/genetics/iyaa043.
- Nutritional and functional characterization of different oat (Avena sativa L.) cultivars / M.S. Ibrahim [et al.] // International Journal of Food Properties. 2020. 23(1). P. 1373–1385. DOI: 10.1080/10942912.2020.1806297.
- Genetic diversity of gliadin-coding alleles in bread wheat (Triticum aestivum L.) from Northern Kazakhstan / M. Utebayev [et al.] // PeerJ. 2019. DOI: 10.7717/peerj.7082.
- Генетическое разнообразие современных российских сортов яровой и озимой твердой пшеницы по глиадинкодирующим локусам / А.М. Кудрявцев [и др.] // Генетика. 2014. Т. 50, № 5. С. 554–559.
- Чертов Н.В. Анализ генетического разнообразия посадок дуба черешчатого (Quercus robur L.) на основании полиморфизма ISSR-маркеров // Фундаментальные и прикладные исследования в биологии и экологии: мат-лы регион. студ. науч. конф. с междунар. участием / Перм. гос. нац. исследовательский ун-т. Пермь, 2018. С. 43–46.
- SSR marker-based DNA fingerprinting of Sub1 introgressed lines in the background of tradi-tional rice varieties of Assam India / A. Gau-tam [et al.] // Indian Journal of Biochemistry and Biophysics. Vol. 59 (3). March 2022. P. 350–356.
- Спектры проламинов в агроэкологической оценке коллекционного материала ячменя / Н.В. Зобова [и др.] // Достижения науки и техники АПК. 2018. № 32 (5). С. 45–47. DOI: 10.24411/0235-2451-2018-10511.
- Динамика генетического разнообразия сортов овса в Тюменской области по авенин-кодирующим локусам / А.В. Любимова [и др.] // Вавиловский журнал генетики и селекции. 2020. Т. 24 (2). С. 123–130. DOI: 10.18699/VJ20.607.
- Любимова А.В. Изучение характера наследования компонентов авенина у гибридов F2 от скрещивания овса посевного сибир-ской селекции // Аграрный вестник Урала. 2022. № 02 (217). С. 48–59. DOI: 10.32417/ 1997-4868-2022-217-02-48-59.
- Генетический контроль авенинов и принципы их классификации / В.А. Портянко [и др.] // Генетика. 1987. Т. 23, № 5. С. 845–853.
- Nei M., Li W. Mathematical model for studying genetic variation in terms of restriction endo-nucleases // Proc. Natl. Acad. Sci. USA, 1979, Vol. 76. P. 5269–5273.
- Биометрия / Н.В. Глотов [и др.]. Л.: Ленин-град. гос. ун-т им. А.С. Пушкина, 1982. 263 с.
- Генетическое разнообразие современных российских сортов яровой и озимой твердой пшеницы по глиадинкодирующим локусам / А.М. Кудрявцев [и др.] // Генетика. 2014. Т. 50, № 5. С. 554. DOI: 10.7868/S001 6675814050099.
- Lyalina E.V., Boldyrev S.V., Pomortsev A.A. Current state of the genetic polymorphism in spring barley (Hordeum vulgare L.) from Rus-sia assessed by the alleles of hordein-coding loci // Russian Journal of Genetics. 2016. Vol. 52, № 6. P. 565–577. DOI: 10.1134/S10227 95416060077.
- Сравнительный анализ динамики генетического разнообразия по глиадинкодирую-щим локусам среди сортов озимой мягкой пшеницы Triticum aestivum L., созданных за 40-летний период научной селекции в Сербии и Италии / А.Ю. Новосельская-Драго-вич [и др.] // Генетика. 2007. Т. 43, № 11. С. 1478–1485.
- Анализ генотипов сортов озимой мягкой пшеницы ННЦ «Институт земледелия НА-АН» по аллелям локусов запасных белков / Е.В. Заика [и др.] // Вестник Белорусской государственной сельскохозяйственной академии. 2014. № 4.С. 53–57.
- Трофимов И.Е. Некоторые результаты популяционно-фенетического анализа Nicro-phorus vespillo (Coleoptera, Silphidae) из калужского городского бора // Зоологический журнал. 2008. № 87 (6). С. 658–664.
- Волкова П.Ю., Гераськин С.А. Анализ полиморфизма супероксиддисмутазы в хронически облучаемых популяциях сосны обыкновенной // Радиационная биология. Радиоэкология. 2012. Т. 52, № 4. С. 370.
- Ульянова Е.В., Позолотина В.Н. Клональ-ное разнообразие и редкие фены в ценопо-пуляциях Taraxacum officinale S. L. из зоны Восточно-Уральского радиоактивного следа // Доклады Академии наук. 2006. Т. 406, № 5. С. 715–717.
