Изменение векторов отбора по массе поросят при рождении в процессе формирования популяции в новых условиях среды

Автор: Князев С.П., Никитин С.В.
Журнал: Сельскохозяйственная биология @agrobiology
Рубрика: Динамика генофондов
Статья в выпуске: 6 т.49, 2014 года.
Бесплатный доступ
Изучение изменений в генофондах популяций домашних животных, вызываемых отбором и отражающих микроэволюционные процессы, представляет как практический, так и теоретический интерес. Как правило, в таких работах используются качественные признаки, контролируемые по принципу «один генотип - один фенотип», что существенно упрощает анализ. Мы исследовали динамические процессы в популяции домашних свиней ( Sus scrofa domesticus ) породы ландрас, оценив вариацию массы поросенка при рождении (один из непрерывных количественных признаков, для которых подобная связь неоднозначна). В частности, проведен статистический анализ данных зоотехнического учета более 26 тыс. поросят, родившихся на протяжении 23 лет в условиях Экспериментального хозяйства Сибирского отделения Академии наук (Новосибирская обл.) - регионального племенного репродуктора ландрасов. Предков животных для формируемой популяции завезли из Прибалтики в начале 1960-х годов. Это был племенной элитный молодняк, оцененный по стандартному комплексу селекционных признаков. Весь период существования популяции отбор ремонтного молодняка осуществляли по комплексу признаков в соответствии с действовавшей Инструкцией по бонитировке свиней. Так как Инструкция не содержала стандартов по крупноплодности (массе новорожденного поросенка), отбор по указанному признаку не проводился. Изменения статистических параметров массы новорожденной особи оценивали за каждый год периода наблюдений, поскольку анализируемое поголовье представляет собой не модельную популяцию вивария, а типичное племенное стадо, в котором непрерывная изменчивость признака сопряжена с постоянной «скользящей» сменой поколений и возрастного состава. При длительном анализе динамики признака, который не был селекционным, выявлен движущий отбор по генотипам для локусов, контролирующих скорость роста свиней в онтогенезе (в пре- и постнатальный периоды). В популяциях, где действует такой отбор, масса поросят при рождении может быть использована для ее прогнозирования и в постнатальный период. Применение регрессии родитель-потомок по признаку «масса при рождении» позволило оценить продолжительность адаптации генофонда популяции к новым условиям среды. Оказалось, что процесс адаптации продлился почти два десятилетия, что составляет пять полных смен генераций. В этой же популяции в то же время действовал стабилизирующий отбор, оптимизирующий массу особи при рождении, отсекая как минимальные, так и максимальные значения. Описанный механизм, когда на фоне стабилизирующего отбора происходят цикличные изменения в направлениях векторов движущего отбора, способствует поддержанию в популяции полиморфизма по локусам, которые контролируют интенсивность пренатального роста и крупноплодность. Следует также отметить наблюдаемую неравномерность волнообразной смены этих циклов.
Свиньи, ландрас, популяция, адаптация, масса поросят при рождении, крупноплодность, регрессия, вектор отбора, движущий отбор, стабилизирующий отбор, микроэволюционые процессы
Короткий адрес: https://sciup.org/142133559
IDR: 142133559 | УДК: 636.4:575.174:591.4 | DOI: 10.15389/agrobiology.2014.6.86rus
Changes of vectors of selection for piglet’s newborn weight during population formation in new environment conditions
Studying changes of gene pools of populations of the domestic animals caused by selection and reflecting micro evolutionary processes, represents both practical and theoretical interest. As a rule, in such works the qualitative traits controlled by the principle «one genotype - one phenotype» are used that significantly simplifies the analysis. We investigated dynamic processes in Landrace population of domestic pigs ( Sus scrofa domesticus ), having estimated a variation of one of continuous quantitative traits, the newborn piglet weight, for which similar relationship is not unambiguous. Statistical analysis of the zootechnical register data was carried out on more than 26 thousand pigs that were born within 23 years at an experimental farm (Novosibirsk Province), being the regional authorized Landrace breed nucleus. Ancestors of the formed population were delivered from Latvia in the early 1960s. They were the elite young animals estimated on a standard complex of selection traits. The entire period of existence of population the formation of its breeding nuclear was carried out on a complex of traits according to existing Instruction for estimation of breeding value of pigs. As the Instruction didn’t contain standards on a large newborn weight, the selection on the specified trait wasn’t made. The changes of statistical parameters of the newborn weight in piglets were estimated for each year of the observation, because the analyzed livestock was not a model laboratory population, but a typical breeding commercial herd in which continuous variability of a trait is interfaced to continuous «sliding» alternations of generations and the variability of age structure. During long-time analysis of the dynamics of this unselected trait, the directional (moving) selection on genotypes for the loci controlling growth rate of pigs in ontogenesis (in pre- and post-natal periods) is revealed. In the populations where such selection works, the newborn weight of piglets can be used for forecasting pig weight during the postnatal period. Application of «parent-descendant» regression on the newborn weight allowed to estimate the duration of a population gene pool adaptation to new environment. It appeared that process of adaptation lasted nearly two decades that maked five full changes of the generations. In the same population at the same time the stabilizing selection optimized an individual animal weight at birth, cutting both minimum and maximum values. The described mechanism includes cyclic vector changes towards driving selection against the stabilizing selection vectors and thus maintains the population polymorphism on loci which control prenatal growth and large weight in the newborns. An observed unevenness of wavy change of these cycles should be also noted.
Текст научной статьи Изменение векторов отбора по массе поросят при рождении в процессе формирования популяции в новых условиях среды
Изменения генофондов популяций, вызываемые как отбором, так и возникшие в результате случайного «дрейфа генов», неоднократно привлекали внимание исследователей (1-3). Среди статей, посвященных этому вопросу, присутствует немалое число выполненных на домашних свиньях (4-12). Однако в таких работах, как правило, используют качественные признаки, удобные однозначностью связи генотипа с фенотипом (один генотип — один фенотип), что существенно упрощает анализ. Мы много лет продолжаем изучать особенности вариации качественных признаков и получили весьма интересные результаты (13-17). В то же время уже первая работа, в которой нами был проведен статистический анализ динамики изменений количественных признаков, вскрыла ряд генетических закономерностей, свойственных именно домашним животным (18). Кроме того, она показала, что признаки, используемые в селекции свиней, позволяют выявлять главным образом закономерности, связанные с искусственным отбором и гораздо менее информативны в отношении отбора естественного. Поэтому был начат поиск признака, пригодного для оценок векторов естественного отбора. В таком качестве первоначально рассматривалось число сосков, но это показатель, хотя и позволял оценить роль случайности в формировании значений количественного признака (19), оказался малочувствителен к динамическим внутрипопуляционным процессам ввиду дискретности и недостаточно широкого диапазона вариации (18, 19). Следующим включенным нами в анализ признаком была масса новорожденного поросенка. Это континуальный (непрерывный) количественный признак с неопределенностью связи генотипа с фенотипом: один фенотип может наблюдаться у разных генотипов, а одному генотипу соответствует множество значений признака (20). Так как масса поросенка при рождении — признак перинатального роста, вариация его значений у гетерозигот включает диапазоны вариации значений у гомозигот по аллелям, составляющим гетерозиготный генотип (21). Поэтому в настоящем исследовании для анализа динамических процессов в популяции свиней был использован непрерывный количественный признак (масса поросенка при рождении) с относительно широким диапазоном вариации и известным генетическим контролем (22-26).
Цель нашей работы — определить, каким формам отбора подвергается масса поросят при рождении в популяции домашних свиней, формируемой при адаптации к новым условиям.
Методика . Исследования проводили на свиньях породы ландрас, рожденных в период 1964-1965 и с 1967 по 1986 годы (Экспериментальное хозяйство, Новосибирская обл.). Предки свиней изучаемой популяции были завезены из Прибалтики в 1961 году как элитный племенной молодняк, соответствующий утвержденному комплексу селекционных признаков (27). В течение периода наблюдений технологии кормления и содержания животных оставались одинаковыми и соответствовали описанным в литературе (28, 29). Животные дважды в сутки получали запаренный стандартный комбикорм типа СК отечественного производства по рационам ВИЖ (30).
При разведении племенного молодняка направления селекции определялись действовавшей инструкцией по поддержанию комплекса признаков для стандарта породы и не предусматривали специального отбора на увеличение размеров животных (9) или отбора по массе при рождении. В процессе создания племенного стада молодняк отбирали 2-кратно — при отъеме (в возрасте 2 мес) и при формировании группы ремонта (5 мес), которая составляла до 25 % от численности репродуктивного ядра.
Для анализа изменений признака сравнивали его статистические параметры по годам наблюдения. Определяли среднее значение массы при рождении у поросят каждого года рождения и сопоставляли его со средними значениями у их отцов и матерей (основное исследование, всего 21 461 потомок от 312 отцов и 1181 матери). Для оценки возможных значений верхней границы показателя дополнительно изучили выборку из 2587 пометов, в которых взвешивали всех родившихся поросят ( n = 26 086).
Статистическую обработку данных проводили общепринятыми методами (31).
Результаты. Свиноферма Экспериментального хозяйства была племенным репродуктором ландрасов в регионе. При формировании популяции, адаптированной к условиям Сибири, специальный отбор на увеличение размеров животных выше стандарта породы не проводился. По- еле завоза ландрасов в хозяйство средняя масса особи при рождении в созданной популяции увеличивалась на протяжении 23 лет, однако это было характерно в первую очередь для особей репродуктивного ядра (отцов и матерей) и лишь в незначительной степени касалось их потомков.
При изучении динамики показателя в качестве наименьшего отрезка времени выбрали календарный год в связи с тем, что поголовье свинофермы было типичным стадом племенного репродуктора сельскохозяйственных животных. В этом стаде (в отличие от модельной популяции из вивария) репродуктивное ядро составляло не группу ровесников из одного и того же поколения, а совокупность разновозрастных (проверяемых и основных) племенных хряков и свиноматок и постоянно переформировывалось за счет ежегодного ввода новых производителей и выбраковки части основного стада (по возрасту или другим причинам). Таким образом, смены поколений в стаде представляют собой непрерывный растянутый во времени процесс. При этом изменчивость изучаемого признака по годам оказывается сопряженной с непрерывной сменой поколений и возрастного состава маточного поголовья. Последний представлял собой из года в год практически стабильную величину, подверженную только случайной вариации, что и послужило главной причиной выбора календарного года как наименьшего отрезка времени при оценке динамики изучаемого показателя.
Вариация массы при рождении (кг) у потомков и их родителей в популяции свиней породы ландрас в период длительной адаптации к разведению в измененных условиях (Экспериментальное хозяйство, Новосибирская обл.)
|
Год |
Потомки |
Отцы |
Матери |
Коэффициент регрессии ( b xy ) |
||||
|
n |
| M ± m |
n |
| M ± m |
n |
| M ± m |
отец—потомок |
мать—потомок |
|
|
1964 |
524 |
0,98±0,007 |
9п |
1,40±0,116 |
40п |
1,24±0,040 |
- 0,006 |
0,091 |
|
1965 |
405 |
1,14±0,012 |
14 |
1,08±0,074 |
31 |
1,13±0,032 |
- 0,140 |
- 0,403 |
|
1967 |
262 |
1,16±0,013 |
10 |
1,19±0,038 |
28 |
1,15±0,041 |
- 0,670 |
- 0,163 |
|
1968 |
355 |
1,20±0,007 |
6 |
1,18±0,048 |
30 |
1,14±0,046 |
- 0,111 |
- 0,123 |
|
1969 |
592 |
1,29±0,019 |
10 |
1,38±0,092 |
56 |
1,36±0,046 |
0,212 |
0,098 |
|
1970 |
1027 |
1,26±0,004 |
22 |
1,48±0,063 |
100 |
1,35±0,030 |
- 0,007 |
0,032 |
|
1971 |
1116 |
1,38±0,009 |
17 |
1,37±0,051 |
94 |
1,45±0,032 |
- 0,189 |
0,231 |
|
1972 |
1506 |
1,47±0,007 |
24 |
1,38±0,074 |
106 |
1,46±0,030 |
- 0,063 |
0,034 |
|
1973 |
1652 |
1,33±0,012 |
14 |
1,39±0,075 |
122 |
1,46±0,025 |
- 0,067 |
0,018 |
|
1974 |
1255 |
1,42±0,005 |
15 |
1,37±0,064 |
98 |
1,43±0,025 |
- 0,030 |
0,037 |
|
1975 |
1066 |
1,35±0,009 |
15 |
1,39±0,070 |
73 |
1,44±0,025 |
- 0,070 |
- 0,042 |
|
1976 |
748 |
1,42±0,010 |
16 |
1,44±0,055 |
74 |
1,51±0,025 |
- 0,056 |
0,029 |
|
1977 |
1329 |
1,18±0,008 |
22 |
1,56±0,056 |
92 |
1,49±0,017 |
0,038 |
0,024 |
|
1978 |
1451 |
1,28±0,007 |
26 |
1,18±0,076 |
109 |
1,24±0,026 |
0,052 |
- 0,028 |
|
1979 |
1720 |
1,35±0,007 |
30 |
1,47±0,040 |
115 |
1,35±0,023 |
0,007 |
0,029 |
|
1980 |
1187 |
1,31±0,008 |
32 |
1,48±0,047 |
101 |
1,36±0,023 |
- 0,045 |
- 0,141 |
|
1981 |
1663 |
1,34±0,007 |
33 |
1,56±0,063 |
128 |
1,34±0,024 |
- 0,009 |
0,085 |
|
1982 |
1114 |
1,34±0,010 |
29 |
1,51±0,039 |
79 |
1,44±0,033 |
0,224 |
- 0,059 |
|
1983 |
724 |
1,45±0,012 |
24 |
1,59±0,062 |
67 |
1,47±0,037 |
0,046 |
0,134 |
|
1984 |
562 |
1,30±0,014 |
15 |
1,55±0,061 |
53 |
1,49±0,046 |
0,110 |
0,191 |
|
1985 |
853 |
1,24±0,010 |
23 |
1,55±0,058 |
74 |
1,47±0,034 |
0,203 |
0,171 |
|
1986 |
350 |
1,12±0,009 |
10 |
1,32±0,053 |
31 |
1,37±0,038 |
- 0,150 |
- 0,022 |
|
Всего 21 461 |
1,32±0,002 |
312 |
1,44±0,017 |
1181 |
1,38±0,008 |
|||
|
r |
0,33; н.д. |
0,61; P < 0,01 |
0,57; P < 0,01 |
|||||
Примечание. 1964, 1969, 1973, 1977, 1981 и 1985 — годы полной смены генерации животных племенного ядра (отцов и матерей); п (в верхнем индексе) — дети животных, завезенных из Прибалтики; M ± m — среднее значение признака и его ошибка; r — коэффициент корреляции между годом и массой особи при рождении; н.д. — недостоверно.
В основное исследование включили только поросят, для которых массу при рождении учитывали индивидуальным взвешиванием как их самих, так и обоих родителей (табл.). Такое сопоставление полученных средних значений позволяет точнее выявлять тренды изменений с учетом случайной вариации признака и ежегодного изменения состава племенного ядра.
Для значений признака положительный тренд оказался статистиче-
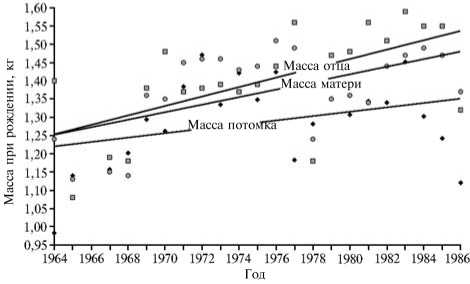
Рис. 1. Тренды по признаку «масса при рождении» в популяции свиней породы ландрас (потомки) и ее эффективной части (отцы — хряки-производители и матери-свиноматки) в период длительной адаптации к разведению в измененных условиях: ♦ — потомки, □ — хряки-производители, о — свиноматки (Экспериментальное хозяйство, Новосибирская обл.).
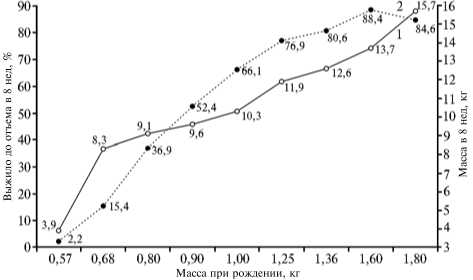
Рис. 2. Сохранность поросят в подсосный период (1) и их масса при отъеме в 8 нед (2) в зависимости от массы поросенка при рождении (графики построены по данным В.Д. Кабанова) (29).
ски значимым (P < 0,01) в группах отцов и матерей, тогда как у потомков (популяции в целом) он был недостоверен и, вероятно, проявился вследствие трендов признака в родительских группах (рис. 1). Таким образом, в репродуктивном ядре имел место отбор на увеличение массы при рождении, тогда как на популяцию в целом его воздействие было незначительно. Отбор в эффективную часть популяции (племенное ядро) включал три компоненты — естественный отбор особей по жизнеспособности в ранний постнатальный период, положительную связь между массой поросенка при рождении и его массой при отъеме, что в явном виде показывают графики (рис. 2), построенные по данным, которые приведены в монографии В.Д. Кабанова (29), а также специфичный для домашних животных стандартизирующий отбор по живой массе (18).
Первые две компо ненты существенно повышали вероятность попадания крупных при рождении особей в репродуктивное ядро популяции. При условии, что рост массы в пре- и постнатальный период могут контролировать одни и те же локусы, в сферу действия стандартизирующего отбора по живой массе попадает и масса новорожденного, несмотря на то, что искусственный отбор по массе при формировании поголовья ремонтного молодняка начинался с 2-месячного возраста (согласно действовавшей в изучаемый период времени Инструкции по бонитировке) (27). Следствием стандартизирующего отбора по массе тела стали наблюдаемые в популяции тренды для признака в родительских группах и статистически значимые различия по массе при рождении между самцами и самками воспроизводящего состава ((ф = 3,06, P < 0,01). Более выраженный положительный тренд (см. рис. 1) и большая масса при рождении (см. табл.) в группе отцов были результатом того, что требования к массе самцов при отборе в воспроизводящий состав существенно выше, чем при отборе самок. Очевидно, что совокупность всех трех компонент отбора формировала в популяции итоговый вектор, направленный не собственно на массу новорожденной особи, а на высокую интенсивность роста как в пре-, так и в постнаталь- ный период. Таким образом, в эффективную часть популяции попадали не просто крупные при рождении особи, но те из них, которые, родившись крупными, продолжали интенсивно расти и в постнатальный период.
’1964 1966 1968 1970 1972 1974 1976 1978 1980 1982 1984 1986 Год
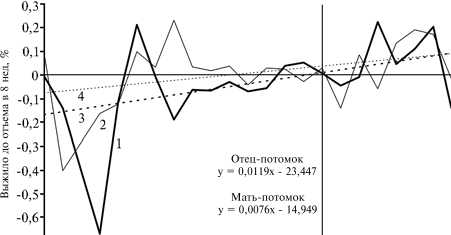
Рис. 3. Изменение коэффициентов регрессии (родитель-потомок) для массы при рождении в популяции свиней породы ландрас, адаптируемой к разведению в непривычных условиях региона: 1 — отец—потомок, 2 — мать-потомок, 3 — тренд коэффициента регрессии отец—потомок, 4 — тренд коэффициента регрессии мать—потомок (Экспериментальное хозяйство, Новосибирская обл.).
Масса новорожденного поросенка — поли-генный признак (20, 21) с выраженной неопределенностью связи генотипа с фенотипом, что подразумевает не только элементы случайности, но и возможность адекватного ответа на внешние воздействия. Параметром, отражающим влияние обоих этих факторов, может служить регрессия между фенотипами родителя и потомка, которая учитывает одновременно два типа неопределенности — для связей генотип—фенотип ро дителя и генотип—фенотип потомка. Свиньи породы ландрас были завезены из Прибалтики в Западную Сибирь, где оказались в непривычных условиях, адаптация к которым повлекла изменения в генофонде формирующейся популяции. Для анализируемого нами признака этот процесс отразила динамика коэффициентов регрессии родитель—потомок, которая для отцов и матерей соответствует (см. табл.) уравнениям линейных трендов (рис. 3).
При рассмотрении такой динамики следует учитывать, что на графиках имеется протяженный начальный отрезок с отрицательными значениями коэффициентов регрессии (см. рис. 3). Можно предположить, что в новых условиях кормления и содержания важное значение приобрело то, что у рождавшихся более крупными поросят сохранность в подсосный период была выше, а масса при отъеме больше (см. рис. 2) (29). В результате популяция оказалась под действием отбора на высокую интенсивность роста поросят в пре- и постнатальный период. Очевидно, в начальный пе риод существования популяции неопределенность связи генотип— фенотип по исследуемому признаку была высокой. Как следствие, у крупных при рождении родителей в потомстве масса поросят при рождении не превышала среднего значения для популяции и, соответственно, регрессия родитель—потомок была отрицательной. Отбор на высокую скорость роста в период онтогенеза постепенно снижал неопределенность связи генотип— фенотип, и через полтора десятилетия (к 1979 году) сформировался генофонд, в котором неоднозначность снизилась до уровня положительной регрессии родитель—потомок (см. рис. 3). Это может означать, что доля особей с адаптированной к местным условиям относительно устойчивой связью генотип—фенотип превысила долю таковых с неустойчивой связью.
Как показывает исследование, в изучаемой популяции ландрасов действовал отбор, направленный на увеличение массы особи при рождении. Тем не менее, масса новорожденного поросенка оставалась относительно стабильной благодаря действию генетико-физиологических механизмов, которое направлено на удержание величины признака в рамках, выход за пределы которых несовместим с жизнью либо для потомка, либо для ма- тери. Иными словами, масса поросенка при рождении может варьировать в разных популяциях или в одной популяции по годам (см. табл.), но в определенных пределах, поэтому отбор по этому признаку, в сущности, неэффективен. Нижняя граница значений массы при рождении, очевидно, определяется высокой смертностью мелких поросят. Механизм, обеспечивающий снижение массы до оптимальной для популяции величины, очевидно, должен быть связан с какими-либо другими признаками. В частности, у свиньи как многоплодного животного неизбежен конфликт между массой поросенка при рождении и числом поросят в помете, что обусловливает отрицательную корреляцию между этими признаками (32). Такая корреляция (r = -0,82; P < 0,001) наблюдалась и в исследуемой популяции.
Возможно, одним из факторов, ограничивающих максимальную массу при рождении, служит общая масса всех поросят-однопометников (масса помета). Теоретически график ее зависимости от числа поросят в помете можно представить в виде двух отрезков. На первом график стремится к значению, предельному для популяции, на втором он колеблется несколько ниже этого уровня. То есть первый отрезок — расту-
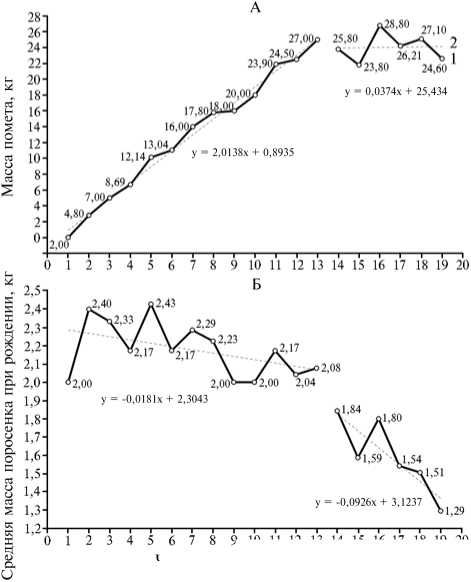
Рис. 4. Масса помета (А) и средняя масса поросенка при рождении (Б) в зависимости от числа поросят в помете в популяции свиней породы ландрас, адаптируемой к разведению в непривычных условиях региона: . 1 — фактическое распределение показателя по классам многоплодия, 2 — тренд значений признака (Экспериментальное хозяйство, г. Новосибирск).
9 10 11 12 13 14 15 16 17 18 19 20
Число поросят в помете, шт.
щая ния, ная щая
наклонная прямая ли-второй — горизонталь-прямая линия, лежа-ниже предельных зна- чений. Для проверки этого предположения были привлечены дополнительные данные по новорожденным поросятам, у родителей которых (одного или обоих) по тем или иным причинам масса при рождении оказалась не зафиксированной в материалах зоотехнического учета. В резуль- тате сформировали выборку из 2587 пометов (26 086 новорожденных поросят), в которых взвешивали всех родившихся поросят, включая мертворожденных. Обнаруженная в этой выборке максимальная масса помета при рождении равнялась 28,8 кг (рис. 4, А). Чтобы определить конечную точку первого (наклонного) отрезка графика, мы сравнили величину достоверности аппроксимации R2 из программы Microsoft Excel для трех наиболее вероятных значений величины помета. Для пометов из 12, 13 и 14 поросят значение R2 составило соответственно 0,9880; 0,9905 и 0,9823. То есть первый (наклонный) отрезок соответствует пометам из 1-13, второй (горизонтальный) — 14-19 особей. Коэффициенты корреляции между числом потомков в помете и массой помета для первого и второго отрезка графика оказались равны соответственно +0,995 (P < 0,001) и +0,039 (недостоверен), иными словами, в целом график адекватно описывал зависимость массы помета при рождении от числа потомков в нем (см. рис. 4, А). Масса помета при рождении связана как с числом особей в помете, так и с массой новорожденного, поэтому ограничения, которые организм самок налагает на максимальную величину общей массы помета, распространяются и на среднюю массу новорожденной особи, приобретая вид отрицательной корреляции между многоплодием и крупноплодностью. Второй график для зависимости между массой особи при рождении и числом потомков в помете, очевидно, должен повторять первый, но в обратном порядке: вначале горизонтальный отрезок, затем понижающийся наклонный (см. рис. 4, Б). Коэффициенты корреляции (r) между числом потомков в помете и средней массой новорожденного были равны -0,467 (недостоверен) и -0,856 (P < 0,05) соответственно для первого и второго отрезка графика, то есть результат согласуется с нашим предположением. Поэтому можно утверждать, что в изученной популяции действовал естественный стабилизирующий отбор, оптимизировавший соотношение массы и числа потомков в помете, компонентой которого служит вектор, направленный на уменьшение массы новорожденной особи.
Поскольку в популяциях свиней заводских пород идет постоянный стандартизирующий отбор по многоплодию, а между многоплодием и массой поросенка при рождении существует отрицательная корреляция, можно предположить, что при усилении отбора по многоплодию должен усиливаться естественный отбор, направленный на уменьшение размеров новорожденного. После того как в генофонде популяции закрепляются аллели, обеспечивающие такое многоплодие, снова должен начаться отбор на большую массу новорожденных поросят (потому что их сохранность в
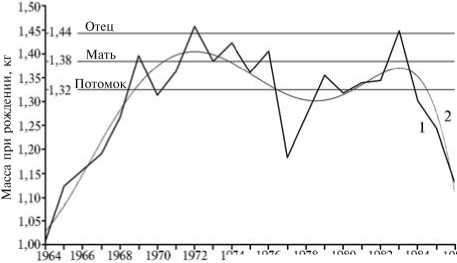
Рис. 5. Изменение массы при рождении в популяции свиней породы ландрас за 23-летний период адаптации к разведению в непривычных условиях региона: 1 — фактическое, 2 — выровненный тренд значений признака (Экспериментальное хозяйство, Новосибирская обл.).
1974 1976 1978 1980 1982 1984 1986 Год
подсосный период выше), который будет продолжаться до тех пор, пока не приведет к снижению многоплодия. В результате начнется следующий цикл со сменой векторов отбора, и так весь период существования популяции. Как следствие конфликта между отбором на увеличение массы особи при рождении и ограничением верхнего значения этого показателя динамика массы новорожденного на достаточно про должительном отрезке времени будет иметь вид неравномерных волнообразных колебаний вокруг оптимального для популяции значения. Именно такой процесс и наблюдался в исследуемой популяции (рис. 5). На протяжении 23 лет были выявлены две волны: среднее значение живой массы росло от 1,01 до 1,46 (с 1964 по 1972 год), затем снижалось до 1,18 кг (1977 год), затем вновь увеличивалось до 1,45 кг (1983 год) и падало до 1,13 кг (1986 год). В целом можно заключить, что давление отбора «сверху» (элиминация наиболее крупноплодных особей, которую отражают графики на рисунке 4 и пояснения к нему) даже несколько сильнее, чем давление «снизу», что, веро- ятно, связано с наличием ряда других факторов, снижающих массу новорожденного поросенка до оптимальной.
Отметим, что при многолетнем мониторинге воспроизводства популяции в течение адаптации к новым условиям примененный нами классический метод популяционных исследований динамики признака по годам (33) оказался эффективнее, чем ряды, построенные по небольшому числу точек (см. табл.), представляющих годы полной смены поколений репродуктивного ядра. Причина, на наш взгляд, в том, что классический подход предназначен для работы именно с реально существующими, а не модельными популяциями. Полученные в настоящем исследовании выводы способствуют развитию представлений о генетико-селекционных аспектах крупноплодности у свиней (20, 34).
Итак, в популяции свиней породы ландрас, которая формировалась в процессе приспособления к новым условиям среды, выявлен движущий отбор по генотипам для локусов, контролирующих скорость роста свиней в целом, включая как пре-, так и постнатальный периоды. В популяциях, где действует такой отбор, масса поросят при рождении может быть использована при прогнозировании роста в постнатальный период. Применение регрессии родитель-потомок по признаку «масса при рождении» позволило оценить длительность адаптации генофонда популяции к новым условиям среды. Оказалось, что процесс адаптации генофонда продлился почти два десятилетия, что составляет пять полных смен генераций. В этот период выявлены два ключевых события: срок достижения «точки» преобладания особей с пониженной неопределенностью связи «генотип-фенотип» составил полтора десятилетия, а достижение популяционного оптимума (которое можно считать завершением процесса адаптации) произошло практически на 5 лет позже. В этой же популяции в то же время действовал и стабилизирующий отбор, оптимизирующий массу особи при рождении, отсекая как минимальные, так и максимальные значения. Представляется, что это должно способствовать поддержанию в популяции полиморфизма по локусам, контролирующим интенсивность пренатального роста и крупноплодность, за счет того, что на фоне стабилизирующего отбора неравномерно (волнообразно) происходят цикличные изменения направления векторов движущего отбора.
ФГБОУ ВПО Новосибирский государственный Поступила в редакцию аграрный университет, 13 августа 2013 года
CHANGES OF VECTORS OF SELECTION FOR PIGLET’S NEWBORN WEIGHT DURING POPULATION FORMATION IN NEW ENVIRONMENT CONDITIONS
S.P. Knyazev1, S.V. Nikitin
