Изучение аллельного состава глиадин-кодирующих локусов селекционных линий твердой пшеницы

Автор: Прядун Ю.П., Любимова А.В., Еремин Д.И.
Журнал: Вестник Красноярского государственного аграрного университета @vestnik-kgau
Рубрика: Агрономия
Статья в выпуске: 7, 2020 года.
Бесплатный доступ
Цель исследования - изучение селекционных линий яровой твердой пшеницы для определения их генетической структуры и происхождения. Материалом для исследования послужили зерновки сорта Безенчукская степная (разновидность hordeiforme) и 4 селекционных линий (leucurum), выделенных из его посевов. Так как причиной появления растений другой разновидности могло быть механическое засорение, также был изучен сорт Марина - как вероятный источник сортовой примеси. Установлено, что ни одна из исследованных линий по компонентному составу глиадина не соответствовала сорту Безенчукская степная. У селекционных линий № 1 и 2 преобладал тип спектра, идентичный II биотипу сорта Марина (95,45 и 93,18 % соответственно). Селекционные линии№ 3 и 4 по компонентному составу глиадинабыли близки спектрам сорта Марина. Генетическая формула глиадина линии № 3 Gli-A1d g; B1d la; A2d a; B2d ou, основного биотипа линии№ 4 - Gli-A1d g; B1d la; A2d d; B2d a. Генетическая дистанция по Dice между II и IV биотипами сорта Марина и линиями № 3 и 4 составила 0,2. Генотипы этих линий представляют собой естественные гибриды или мутантные формы, в формировании которых участвовал сорт Марина. Генотипы, от которых были получены исследованные селекционные линии, появились в посевах сорта Безенчукская степная в результате механического засорения. Однако учитывая, что линии № 3 и 4 характеризуются высокими показателями качества зерна и уникальными спектрами глиадина, они были оставлены для дальнейшего изучения и включения в селекционный процесс.
Твердая пшеница, глиадинкодирующие локусы, электрофорез, электрофоретический спектр, генетическая дистанция
Короткий адрес: https://sciup.org/140250684
IDR: 140250684 | УДК: 581.19:631.52:633.112.1 | DOI: 10.36718/1819-4036-2020-7-3-10
The study of allele composition of gliadin-coding loci of durum wheat selection lines
The aim of the research was to study selection lines of spring durum wheat to determine their genetic structure and origin. The material for the study was the grains of Bezenchukskaya stepnaya variety (hordeiforme form) and 4 selection lines (leucurum) isolated from its crops. Since the cause of the appearance of plants of another variety could be mechanical clogging, the variety Marina was studied as a likely source of varietal impurity. It was found that none of the studied lines on the component composition of gliadin corresponded to Bezenchukskaya stepnaya variety. In selection lines № 1 and 2, the type of spectrum prevailed, which was identical to the II biotype of the variety Marina (95.45 and 93.18 %, respectively). According to the component composition of gliadin, selection lines № 3 and 4 were close to the spectra of the variety Marina. Genetic formula of gliadin line№ 3 Gli-A1d g; B1d la; A2d a; B2d a, the main biotype of line № 4 - Gli-A1d g; B1d la; A2d d; B2d a. The genetic distance according to Dice between the II and IV biotypes of the variety Marina and lines № 3 and 4 was 0.2. The genotypes of these lines were natural hybrids or mutant forms, in the formation of which the variety Marina was involved. The genotypes from which the studied selection lines were obtained appeared in the crops of Bezenchukskaya stepnaya variety as a result of mechanical clogging. However, taking into account that lines № 3 and 4 had been characterized by high grain quality and unique gliadin spectra; they were left for further study and inclusion in selection process.
Текст научной статьи Изучение аллельного состава глиадин-кодирующих локусов селекционных линий твердой пшеницы
Введение. Твердая пшеница (Triticum durum Desf.) – сельскохозяйственная культура, зерно которой является ценнейшим сырьем для макаронной и кондитерской промышленности, а также для создания круп и диетического питания [1]. В селекционном процессе при выведении новых сортов, обладающих комплексом хозяйственно ценных и адаптивно значимых призна- ков, важное место занимает совершенствование системы оценки селекционного материала. На современном этапе селекции для этой цели применяются методы молекулярного и биохимического маркирования [2–4]. Наиболее доступным и эффективным в настоящее время является метод электрофореза запасных спирторастворимых белков – проламинов [5, 6].
Глиадины – запасные спирторастворимые белки зерна пшеницы – достаточно хорошо изучены и успешно применяются в селекционном процессе. Проламины твердой пшеницы наследуются кодоминантно, группами – блоками компонентов и контролируются четырьмя локусами генов – Gli-A1d, Gli-B1d, Gli-A2d и Gli-B2d , расположенными на коротких плечах хромосом первой и шестой гомеологической группы [7]. Глиа-дины характеризуются высоким уровнем полиморфизма, что позволяет, основываясь на их компонентном составе, различать до 80 % сортов [8]. Это дает возможность использовать такие белки для регистрации и идентификации сортов, биотипов и линий, определения биотип-ного состава сортов, а также позволяет проводить оценку селекционного материала. В частности, проламины используются для подтверждения гибридности особей, выявления нетипичных генотипов и установления причин их появления [9–12]. Понимание природы возникновения таких генотипов крайне важно для селекции и семеноводства, особенно если они были обнаружены в гетерогенных сортах, полученных из гибридных популяций. В этом случае появление нетипичных растений может быть объяснено не только механическим засорением, но и разнообразными биологическими процессами: мутациями, свободным опылением, расщеплениями, – происходящими внутри популяции самого сорта. При этом новые генотипы могут стать ценным исходным материалом для селекции. Для включения в селекционный процесс необходимо подтвердить, что они не представляют собой сортовую примесь, т. е. не идентичны генетически уже существующим сортам.
Цель исследования : изучение аллельного состава глиадин-кодирующих локусов селекционных линий твердой пшеницы для определения возможности их включения в селекционный процесс.
Задачи исследования: изучить компонентный состав глиадина селекционных линий; оценить генетическое сходство между исследуемыми линиями и сортами твердой пшеницы; определить причину появления нетипичных генотипов.
Материалы и методы исследования. Материалом для исследования послужили 4 селекционные линии, выделенные из посевов сорта твердой пшеницы Безенчукская степная в Челябинском научно-исследовательском институте сельского хозяйства.
Сорт Безенчукская степная имеет разновидность hordeiforme (табл. 1) . Но при возделывании его на полях Челябинского НИИСХ в посевах были обнаружены нетипичные растения разновидности leucurum.
Таблица 1
Характеристика сортов твердой пшеницы, возделываемых в Челябинском НИИСХ
|
Сорт |
Учреждение-оригинатор |
Происхождение* |
Разновидность |
|
Безенчукская степная |
Самарский НИИСХ имени Н.М. Тулайкова |
Индивидуальный отбор из гибридной популяции F 2 Саратовская золотистая / Безенчукская 182 // Безенчук-ская 182 // Гордеиформе 740 |
hordeiforme |
|
Марина |
Самарский НИИСХ имени Н.М. Тулайкова |
Индивидуальный отбор из F 2 гибридной популяции Валентина / Гордеи-форме 1434. В родословную Гор-деиформе 1434 входят сорта Безен-чукская 182, Саратовская золотистая и Гордеиформе 740 |
leucurum |
*Данные официального сайта Самарского НИИСХ имени Н.М. Тулайкова (URL:
.
Наиболее очевидная причина появления особей другой разновидности – механическое засорение. В данном случае источником появления сортовой примеси мог быть сорт твердой пшеницы Марина, также возделываемый на полях института (см. табл. 1). Однако по показателям продуктивности колоса и качества зерна нетипичные генотипы превосходили сорта Бе-зенчукская степная и Марина (табл. 2). Эти генотипы были размножены и послужили основой для получения 4 селекционных линий твердой пшеницы.
Анализы проводили по общепринятым методикам: ГОСТ 10842-89, ГОСТ 10846-91, ГОСТ 54478-2011, ГОСТ 10987-76, ГОСТ 10840-2017; урожайность определяли путем ручного обмолота снопов с последующим пересчетом на т/га.
Для оценки генетической структуры и происхождения селекционных линий были исследованы запасные спирторастворимые белки зерна методом нативного электрофореза. Анализы проводили в лаборатории сортовой идентификации семян Института прикладных аграрных исследований и разработок Государственного аграрного университета Северного Зауралья в 2019 г.
Исследовали по 100 индивидуальных зерновок, отобранных методом случайной выборки от каждой линии, а также от сортов Безенчукская степная и Марина. Анализ проводили в соответствии со стандартной методикой [13]. В качестве стандарта использовали сорт твердой пшеницы Langdon. Идентификацию аллельных состояний блоков компонентов глиадина проводили в соответствии с опубликованными каталогами [7, 14]. За эталонные были приняты спектры сортов Безенчукская степная и Марина, опубликованные в Сводном каталоге электрофоретических спектров глиадина сортов-эталонов, районированных в Российской Федерации до 2010 г. включительно [13].
Таблица 2
Результаты экологического испытания сортов и селекционных линий яровой твердой пшеницы, паровой предшественник, 2018 г.
|
СК s с; i— о о |
аз X ф w аз >S аз о |
1_____ аз £ О Ф аз ^8 |
ф ¥ 1 £ а. ® й ГО о щ |
Клейковина |
1— о сЕ о ф о |
=Е |
|
|
ф аз О О |
|||||||
|
Безенчукская степная |
2,8 |
48,9 |
12,29 |
33,9 |
99, II |
61,0 |
788 |
|
Марина |
2,0 |
53,2 |
11,41 |
30,9 |
90, II |
56,0 |
775 |
|
Линия № 1 |
3,4 |
46,9 |
14,44 |
32,9 |
95, II |
95,0 |
836 |
|
Линия № 2 |
3,3 |
50,1 |
16,60 |
36,9 |
107, III |
94,0 |
824 |
|
Линия № 3 |
3,1 |
53,6 |
16,04 |
35,2 |
106, III |
96,0 |
801 |
|
Линия № 4 |
3,4 |
49,4 |
14,68 |
32,0 |
102, II |
96,0 |
810 |
|
НСР 05 |
0,2 |
2,7 |
0,72 |
1,8 |
5 |
3,7 |
65 |
Для определения генетического сходства между изучаемыми сортами и линиями была применена кластеризация методом попарного внутригруппового невзвешенного среднего (UPGMA – Unweighted Pair-Group Method with Arithmetic Mean). В качестве индекса подобия использовали коэффициент Dice. Построение дендрограммы выполняли с использованием программы TREECON 1.3b.
Результаты исследования. В результате проведенного исследования установлено, что спектры глиадина сортов Безенчукская степная и Марина, возделываемых в Челябинском НИ-ИСХ, соответствуют эталонным. Генетические формулы глиадина сортов имеют вид: Безен-чукская степная – Gli-A1d g; B1d a; A2d a; B2d h+hj; B5d a , Марина – Gli-A1d g; B1d c; A2d a; B2d h+a+h+a; B5d o+o+b+b.
Анализ селекционных линий показал, что линия № 3 гомогенная по компонентному составу глиадина. Линии № 2 и 4 состояли из 2 биотипов, линия № 1 – из 3 (табл. 3).
Ни одна из исследованных линий не соответствовала сорту Безенчукская степная. Это указывает на то, что генотипы, от которых были получены эти линии, появились в посевах сорта Безенчукская степная в результате механического засорения. У селекционной линии № 1 преобладал тип спектра (95,45 %), по компонентному составу глиадина идентичный II био- типу сорта Марина. К этому же биотипу относилась большая часть исследованных зерновок линии № 2 (93,18 %). Остальные типы спектра, обнаруженные в этих линиях, встречались с частотой от 1,14 до 6,82 % и соответствовали биотипам линий № 3 и 4. Селекционные линии № 3 и 4 по компонентному составу глиадина были близки спектрам сорта Марина (рис. 1), в каждом случае отличаясь аллелями по двум локусам.
Для того чтобы оценить генетическое сходство исследуемых образцов, нами была проведена кластеризация (рис. 2).
В результате образцы разделились на 2 кластера. Первый образовали I и III биотипы сорта Марина и сорт Безенчукская степная. Оставшиеся образцы сформировали второй кластер, включающий 2 подкластера (Dice = 0,2). В первый подкластер вошли II и IV биотипы сорта Марина, а также основные биотипы селекционных линий № 1 и 2. Генетическая дистанция по Dice между сортом Марина и этими биотипами равнялась нулю. Второй подкластер сформировали две группы образцов. В первую группу объединились селекционная линия № 3 и идентичные ей биотипы, выявленные нами в линиях № 1, 2 и 4. Вторую группу сформировали основной биотип линии № 4 и II биотип линии № 2. Генетическая дистанция между линией № 3 и основным биотипом линии № 4 составила 0,16.
Таблица 3
Результаты лабораторного сортового контроля селекционных линий твердой пшеницы
|
Номер линии |
Кол-во типов спектра, шт. |
Соответствие спектру сорта (да/нет) |
Формула глиадина |
|
|
Марина |
Безенчукская степная |
|||
|
1 |
3 |
Да, II биотип (95,45 %) |
Нет |
Преобладающий биотип: Gli-A1d g; B1d c; A2d a; B2d a Gli-A1 d g; Bi d la; A2 d d; B2d a (3,41 %); Gli-A1 d g; Bi d la; A2d a; B2d ou (1,14 %) |
|
2 |
2 |
Да, II биотип (93,18 %) |
Нет |
Преобладающий биотип: Gli-A1d g; B1d c; A2d a; B2d a Gli-Al d g; Bi d la; A2d a; B2d ou (6,82 %) |
|
3 |
1 |
Нет |
Нет |
Gli-A1d g; B1d la; A2d a; B2d ou |
|
4 |
2 |
Нет |
Нет |
Преобладающий биотип: Gli-A1d g; B1d la; A2d d; B2d a Gli-A1d g; Bi d la; A2d a; B2d ou ( 1,14 %) |
Л1 Л2 ЛЗ L. Б.с. М. Л4 ЛЗ L.
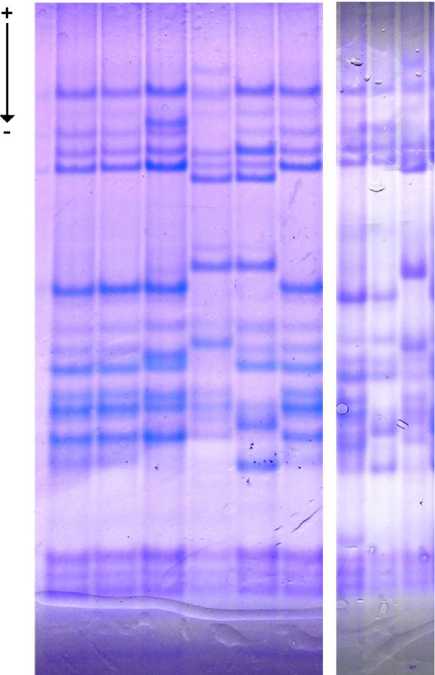
Рис. 1. Электрофоретические спектры преобладающих биотипов селекционных линий твердой пшеницы: Л1–Л4 – линии № 1–4; Б.с. – Безенчукская степная (I биотип); М. – Марина
(II биотип); L. – Langdon (стрелкой указано направление миграции белков при электрофорезе)
В .4 03 02 0.1
-I-----------------------I-----------------------1-----------------------I-----------------------
Марина (I)
Марина (III)

Безенчукская степная (I)
Безенчукская степная (II)
Линия №1 (III)
Линия №2 (II)
Линия №3
Линия №4 (II)
Линия №4 (I)
Линия №1 (II)
— Марина (IV)
■ Линия №2 (I) Марина (II)
Линия №1 (I)
Рис. 2. Дендрограмма сортов и селекционных линий твердой пшеницы
(шкала показывает значение генетической дистанции по Dice, на ветвях указаны значения bootstrap-индексов, в скобках – номера биотипов)
Близость сорта Безенчукская степная к I и III биотипам сорта Марина объяснима общностью их происхождения (см. табл. 1). Компоненты глиадина наследуются блоками, частота рекомбинаций внутри которых крайне низка. Характер наследования блоков компонентов глиадина кодоминантный. Это выражается в том, что при скрещивании гибриды F 1 имеют весь набор компонентов проламина родительских особей [15]. В следующих поколениях происходят расщепления, однако все потомки будут иметь разнообразные комбинации только тех блоков компонентов, которые были у родительских особей.
Отсутствие генетических различий между первыми биотипами линий № 1, 2 и II биотипом сорта Марина подтверждает, что семенной материал сорта Безенчукская степная подвергся механическому засорению. Линии № 1 и 2 представляют собой размноженный II биотип сорта Марина. Механическое смешивание сортов – частое явление на предприятиях, возделывающих несколько сортов одной культуры. Этому способствует использование для посева, уборки и доработки зерна одной и той же техники и плохая ее очистка от семян предыдущего сорта.
Линии № 3 и 4 не идентичны, но генетически близки сорту Марина, что указывает на их родство. На наш взгляд, они появились в результате биологического засорения. Появление биологической примеси возможно, например, в случае расщеплений при остаточной гетерозиготности сорта, а также в результате спонтанной гибридизации и мутационных процессов [16]. Сорт Марина включен в Государственный реестр селекционных достижений РФ в 2009 г. и возделывается более 10 лет. Остаточная гетерозиготность в данном случае маловероятна. Для твердой пшеницы характерно закрытое цветение, однако, как и у других самоопыляющихся сельскохозяйственных культур, часть цветков может цвести открыто. Этому способствует повышенная температура и влажность воздуха [17]. Результатом становятся естественные скрещивания, которые приводят к появлению новых генотипов. По нашему мнению, генотипы линий № 3 и 4 представляют собой естественные гибриды либо мутантные формы, в формировании которых участвовал сорт Марина. Учитывая, что линии № 3 и 4 характеризуются высокими показателями качества зерна, они являются перспективными для дальнейшего изучения и включения в селекционный процесс.
Выводы
-
1. В результате электрофоретического анализа установлено, что все селекционные линии представляют собой механическую примесь. Линии № 1 и 2 имеют генетическую формулу глиадина Gli-A1d g; B1d c; A2d a; B2d a, идентичны II биотипу сорта Марина.
-
2. Селекционные линии № 3 и 4 по компонентному составу глиадина отличаются от сорта Марина (Dice = 0,2). Генетическая формула глиадина линии №3 Gli-A1d g; B1d la; A2d a; B2d ou, основного биотипа линии № 4 – Gli-A1d g; B1d la; A2d d; B2d a. Эти генотипы представляют собой спонтанные гибриды или мутантные формы II биотипа сорта Марина.
-
3. Линии № 3 и 4, характеризующиеся уникальными спектрами глиадина, оставлены для дальнейшего изучения и включения в селекционный процесс.
Список литературы Изучение аллельного состава глиадин-кодирующих локусов селекционных линий твердой пшеницы
- Самофалова Н.Е., Иличкина Н.П., Авраменко М.А. и др. Коммерческие сорта твердой пшеницы и особенности их семеноводства // Зерновое хозяйство России. 2016. № 6 (48). С. 42-47.
- Давоян Э.Р., Беспалова Л.А., Давоян Р.О. и др. Использование молекулярных маркеров в селекции пшеницы на устойчивость к бурой ржавчине в Краснодарском НИИСХ им. П.П. Лукьяненко // Вавиловский журнал генетики и селекции. 2014. Т. 18, № 4/1. С. 732-738.
- Хлесткина Е.К. Молекулярные маркеры в генетических исследованиях и в селекции // Вавиловский журнал генетики и селекции. 2013. Т. 17, № 4/2. С. 1044-1054.
- Тоболова Г.В. Изменение биотипного состава сорта мягкой пшеницы Тюменская 80 в процессе семеноводства // Аграрный вестник Урала. 2009. № 10. С. 12-14.
- Любимова А.В., Ярова Э.Т., Еремин Д.И. Изменение биотипного состава сортов яровой тритикале в процессе возделывания // Вестник КрасГАУ. 2018. № 5 (140). С. 3-8.
- Якубышина Л.И., Казак А.А., Логинов Ю.П. Использование метода электрофореза в семеноводстве ячменя сорта Одесский 100 // Известия Оренбургского государственного аграрного университета. 2017. № 5 (67). С. 56-59.
- Кудрявцев А.М. Создание системы маркеров твердой пшеницы (T. durum Desf.) и ее применение в научных исследованиях и практических разработках: автореф. дис. … д-ра биол. наук. М., 2007. 48 с.
- Поморцев А.А., Кудрявцев А.М., Конарев В.Г. и др. Методика проведения лабораторного сортового контроля по группам сельскохозяйственных растений. М.: Росминформагротех, 2004. 96 с.
- Остапенко А.В., Тоболова Г.В. Создание базы данных сортов рода Avena L. На основе изменчивости компонентного состава проламинов // Аграрный научный журнал. 2015. № 12. С. 24-26.
- Тоболова Г.В. Идентификация и сортовая чистота партий элиты пшеницы в Тюменской области // Сибирский вестник сельскохозяйственной науки. 2012. № 3 (226). С. 12-18.
- Поморцев А.А., Лялина Е.В., Калабушкин Б.А. Гордеин-кодирующие локусы как генетические маркеры у ячменя // Тр. по прикладной ботанике, генетике и селекции. 2009. Т. 165. С. 32-36.
- Любимова А.В., Еремин Д.И. Региональные особенности полиморфизма авенинов овса, возделываемого в Западной Сибири // Тр. по прикладной ботанике, генетике и селекции. 2018. Т. 179, № 2. С. 85-95.
- DOI: 10.30901/2227-8834-2018-2-85-95
- Упелниек В.П., Новосельская-Драгович А.Ю., Шишкина А.А., Мельник В.А. и др. Лабораторный анализ белков семян пшеницы: технологическая инструкция. М.: ВАШ ФОРМАТ, 2013. 173 с.
- Кудрявцев А.М., Дедова Л.В., Мельник В.А., Шишкина А.А. и др. Генетическое разнообразие современных российских сортов яровой и озимой твердой пшеницы по глиадинкодирующим локусам // Генетика. 2014. Т. 50, № 5. С. 554-559.
- Остапенко А.В., Тоболова Г.В. Применение метода электрофореза проламинов овса для определения гибридной природы зерен F1 // Вестник КрасГАУ. 2017. № 2 (125). С. 14-21.
- Розова М.А., Зиборов А.И., Егиазарян Е.Е. Изменение урожайности и параметров качества сорта яровой твердой пшеницы Харьковская 46 под влиянием экологических факторов // Тр. по прикладной ботанике, генетике и селекции. 2017. Т. 178, № 3. С. 59- 66.
- DOI: 10.30901/2227-8834-2017-3-59-66
- Тоболова Г.В. Биология цветения у пшеницы Triticum carthlicum Nevski в лесостепи Тюменской области // Земледелие. 2013. № 6. С. 43-45.
