Изучение механизмов противомикробного действия соединения (+)-erogorgiane
-erogorgiane")
Автор: Нестерова Л.Ю., Ткаченко А.Г., Писцова О.Н.
Журнал: Вестник Пермского университета. Серия: Биология @vestnik-psu-bio
Рубрика: Микробиология
Статья в выпуске: 4, 2016 года.
Бесплатный доступ
Исследованы антибактериальная активность и возможные механизмы действия химически синтезированного соединения (+)-erogorgiane, аналога природного антибиотика, выделенного из морских кораллов Pseudopterogorgia elisabethae. Показано, что оно обладает бактерицидной активностью по отношению к Mycobacterium smegmatis как в логарифмической фазе роста культуры, так и в стационарной. С увеличением времени воздействия снижается концентрация, вызывающая полную гибель клеток в культуре. Эффект (+)-erogorgiane проявляется также по отношению к нерас-тущим клеткам M. smegmatis, которые являются более устойчивыми к действию антибиотиков ри-фампицина, изониазида и левофлоксацина, традиционно используемых для лечения туберкулеза. В то же время грамотрицательный микроорганизм Escherichia coli показал высокую устойчивость к данному препарату. Обнаружено концентрационно-зависимое подавление биопленкообразования М. smegmatis в присутствии (+)-erogorgiane. Ингибирующее действие оказывали как сублетальные, так и субингибиторные концентрации препарата, которые не оказывали влияние на рост культуры и не вызывали гибели клеток. С помощью атомно-силовой микроскопии установлено влияние (+)-erogorgiane на поверхностные структуры клетки: под действием сублетальных концентраций антибиотика увеличивается шероховатость клеточной поверхности. Получить мутантов, устойчивых к действию антибиотика высевом на твердую среду, содержащую препарат в высокой концентрации, не удалось.
Антибиотик, биопленки
Короткий адрес: https://sciup.org/147204794
IDR: 147204794 | УДК: 579.22
Study of the (+)-erogorgiane antimicrobial action mechanisms
The antibacterial activity of a chemically synthesized counterpart of natural antibiotic (+) - erogorgiane extracted from marine coral Pseudopterogorgia elisabethae was studied. This substance has been shown to have bactericidal effect against Mycobacterium smegmatis culture in both logarithmic and stationary growth phases. At the same time, gram negative Escherichia coli is resistant to this compound. The concentration-dependent inhibition of M. smegmatis biofilm formation under sublethal antibiotic action has been observed. The effect of the test compound on the cell surface structure was detected with atomic force microscopy. The attempts to obtain antibiotic resistant mutants on agar media with high antibiotic concentrations failed.
Текст научной статьи Изучение механизмов противомикробного действия соединения (+)-erogorgiane
Туберкулез, вызываемый Mycobacterium tuberculosis, уже в течение многих десятилетий остается одной из главных проблем здравоохранения, являясь причиной более чем миллиона смертей ежегодно [WHO Global Tuberculosis Report, 2015]. Наиболее существенной проблемой, препятствующей эффективному лечению туберкулеза, является быстрое появление ангибиотикорезистснгных форм возбудителя.
Особую тревогу' вызывает распространение штаммов с множественной лекарственной устойчивостью, которое делает существующие методы лечения неэффективными [Migliori, Sotgiu. Gandhi et al, 2013J. Лечение лекарственно устойчивых форм туберкулеза ? как правило, является дорогостоящим и предусматривает использование высокотоксичных препаратов с многочисленными побочными эффектами. В связи с Этим актуальным является поиск новых эффективных противотутберкулезных средств.
В последние годы внимание исследователей привлекли морские кораллы Pseudopterogorgia elisabethae, обитающие в Карибском море и продуцирующие необычные соединения, обладающие
(С Нестерова Л. Ю., Ткаченко А. Г., Писцова О. Н , 2016
фармакологической активностью. Ранее было обнаружено. что соединение дитерпеновой природы (+)-erogorgiane, выделенное из этих организмов,, обладает противотуберкулезной активностью [Do Souza. 2006], однако, изучение сто биологических свойств не проводилось в связи с трудностями его выделения из морских кораллов. Следует отметить что данное соединение по своей структуре значительно отличается от используемых в клинической практике антибиотиков различных классов. Исходя из этого, можно предполагать. что (+)-erogorgiane имеет принципиально иной способ воздействия на бактерии.
Поскольку’ для оценки возможности дальнейшего использования данного вещества в качестве лекарственного средства необходимо понимание механизмов действия антибиотика, целью данной работы является изучение антибактериальной активности и возможных механизмов противомикробного действия искусственно синтезирова иного соединения (+)-erogorgiane, которое является аналогом природного вещества, выделенного из морских кораллов.
Для изучения биологической активности и механизмов антибактериального действия (+)-erogorgiane в качестве объекта исследования выбран микроорганизм Mycobacterium smegmatis. близкий к М. tuberculosis^ в то же время нспато-генный и быстрорастущий и потому широко используемый в качестве модельного объекта при исследованиях физиологии возбудителя туберкулёзной инфекции.
Материалы и методы исследования
В качестве объектов исследования использовали штамм Mycobacterium smegmatis me'155. который сохраняли на чашке Петри с LB агаром (Sigma), культивировали на среде Middlebrook 7Н9 (Difco) с добавлением 0.005% Tween 80 при 37°С, 200 об/мин и штамм Escherichia coli GC 4468, который сохраняли на скошенном LB агаре и культивировали на глюкозо-минеральной среде М9 в термостатируемом шейкере GFL 1092 (Германия) 37°С 100 об/мин.
Для определения антибиотикочувствительности полученный инокулят разводили свежей средой до оптической плотности (ОП) 0.1 (□=600нм), доращивали в указанных условиях до нужной стадии роста и помещали 180 МКЛ культуры в лунки полистиролового 96-лу ночного планшета, куда добавляли 20 мкл раствора антибиотика нужной концентрации и тщательно перемешивали. Культивировали без перемешивания (время культивирования указано на графиках). По окончании культивирования количество живых клеток определяли с помощью подсчета колониеобразутощих единиц (КОЕ) после высева проб на чашки Петри с LB агаром.
Для определения влияния антибиотика на био- пленкообразование М. smegmatis выращенную в течение 24 ч. культуру откручивали на центрифуге, удаляли надосадочную жидкость и ресуспендиро-вали в свежей среде Middlebrook 7Н9 (Difco) без добавления Tween 80, доводя до ОП 0.L затем разводили ещё в 10 раз и помещали в лунки планшета. Культивировали в термостате при 37°С без встряхивания. Через 48 ч. из лунок осторожно удаляли планктонную культуру поверхностную пленку' высушивали и фотографировали. Для определения количества КОЕ в соответствующих пробах производили аналогичный посев на среду, содержащую 0.005% Tween 80. затем готовили десятикратные разведения и делали высевы на LB агар.
Профили поверхности клеток изучали с помощью атомно-силовой микроскопии (АСМ) с использованием совмещенной системы сканирования, состоящей из атомно-силового микроскопа Asylum MFP-3D-BIO (Asylum Research. США) и конфокального лазерного сканирующего микроскопа Olympus FV1000. (Olympus Corporation, Япония). Сканирование осуществляли в полукон-тактном режиме на воздухе с использованием кремниевого кантилевера AC240TS с резонансной частотой 50-90 кГц и константой жесткости 0.5-4.4 Н/м. Для характеристики структуры поверхности клеток (шероховатость Sq, индекс I) получали двух- и трехмерные топографические изображения бактерий. Обработку' микрофотографий проводили с помощью программы Igor Pro 6.22 А (WaveMetrics. США).
Статистическая обработка результатов проведена с использованием пакета стандартных программ Statistica 6.0 (“StatSoft ТпсД 2001). Различия считали значимыми при р<0.05.
Результаты и их обсуждение
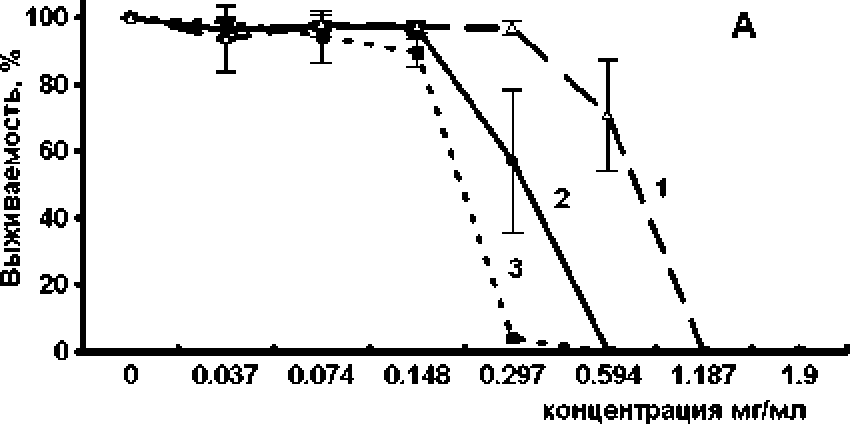
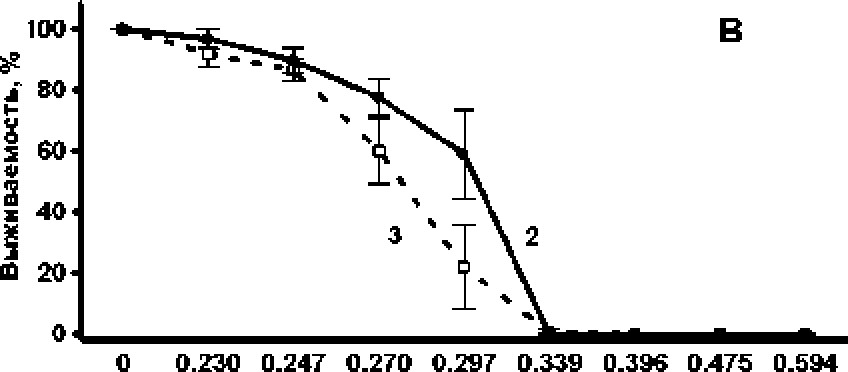
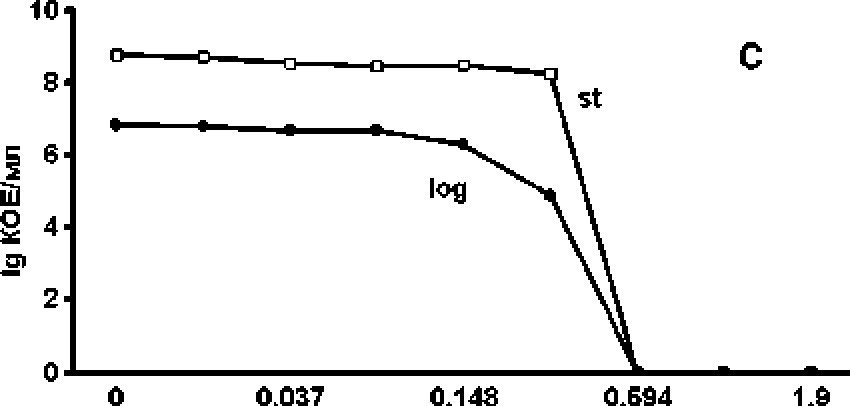
На первом этапе исследования была проведена оценка характера и интенсивности антибактериального действия исследуемого вещества. Показано, что (+)-erogorgiane обладает бактерицидным действием по отношению к М. smegmatis. При этом с увеличением времени воздействия снижается концентрация, которая вызывает полную гибель клеток в культуре (рис. 1А). При более подробной расгитровкс исследуемого препарата удалось установить концентрации, которые оказывают сублс-тальное действие, те. снижают количество клеток, не вызывая полной гибели культуры (рис. 1В). Однако разница между’ суббактериостатическими концентрациями, которые не оказывают влияния на количество клеток, и бактерицидными концентрациями была невелика. Данные концентрации отличались менее чем в 2 раза (рис. 1В). Следует отметить, что бактерицидный эффект исследуемого препарата проявлялся как по отношению к культуре, находящейся в логарифмической фазе роста, так в стационарной (рис. 1С), которая в большинстве случаев является более резистентной к действию различных неблагоприятных факторов, в том числе антибактериальных препаратов [Leisner, Jorgensen, Middelboe. 2016].
концентрация мг/мл
концентрация мг'мл
Рис. L Бактерицидное действие антибиотика на культуру Mycobacterium smegma tis.
Ашибиотк добавляли в культуру в ло^рифмической (А, В, С) и стационарной (С) фазе роста. Экспозиция с антибиотиком 1 - 5 ч ; 2, С - 24 ч.; 3 - 48 ч.
Эффект антибиотика проявлялся также по отношению к нерастущим клеткам, которые были помещены в буфер и после этого подвергнуты действию антибиотика. Такие клетки являются более устойчивыми к действию антибиотиков. Традиционные антибиотики, применяемые для лечения туберкулеза. такие как изониазид, рифампицин и ле-вофлоксацин, в аналогичных условиях нс приводили к полной гибели культуры. Более того, в культуре, взятой в стационарной фазе роста, изониазид практически не имел эффекта на количество клеток, а рифампицин снижал число клеток на 2 порядка (рис. 2). В этом эксперименте мы использовали концентрации антибиотиков, которые намного превышают терапевтические и в 100 раз превышают концентрации, которые препятствуют росту культуры (МПК). Таким образом, (+)-erogorgianc, в отличие от традиционных антибиотиков, способен нс только препятствовать размножению микобактерий, но и полностью убивать клетки вне зависимости от стадии роста культуры, препятствуя образованию персисторных форм ми-
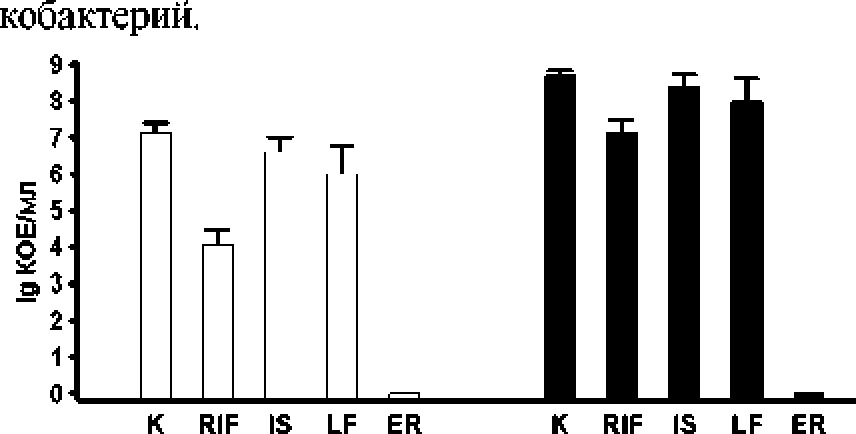
Рис. 2, Действие антибиотиков на нерастущую культуру М. smegmatis.
Антибиотики добавляли в культуру М. smegmatis^ взятую из логарифмической (светлые столбцы) и стационарной (черные столбцы) фазы роста, отмытую от питательной среды и ресуспецдированную в PBS , буфере. Экспозиция 24 ч.
К - контроль без добавки антибиотика; RIF - рифампицин (488 мкг/мл); IS - изониазид (488 мкг/мл); LF -левофлоксацин (63 мкг/мч); ER - (+)-erogorgiane (399 мкг/мч)
Грамотрицательный микроорганизм Esch eri -chia coU. в свою очередь, оказался совершенно нечувствительным к исследуемому антибиотику7. Нам не удалось обнаружить не только бактерицидного, но также и бактериостатического эффекта ни в одной из исследованных концентраций препарата. Исходя из этого, можно предположить, что мишень для действия (+)-erogorgiane специфична для грамположительных микроорганизмов, а возможно, для микобактерий. Несмотря на то, что действие исследуемого соединения на культуру Е. сой не сопровождается гибелью клеток, нами был обнаружен отклик некоторых генов, ответственных за адаптацию этого микроорганизма к стрессовым условиям на его действие (данные не показаны). Данный подход будет использован для дальнейшей работы по расшифровке механизма действия антибиотика.
Следующий этап исследований был посвящен оценке влияния (+)-erogorgianc на способность М. smegmatis к биопленкообразованию. Известно, что микроорганизмы в составе биопленок значительно более устойчивы к действию антибиотиков и иммунной системы хозяина [Syal, Maiti, Naresh et al., 2016]. Хотя роль биопленкообразования в патогенезе микобактерий остается неясной, многие их виды способны формировать биопленки [Arai, Niikawa, Kobayashi, 2013].
Установлено, что присутствие (+)-erogorgiane подавляет биопленкообразование М, smegmatis (рис. 3). Ингибирующее действие оказывали как суб л стальные, так и субингибиторные концентрации препарата, которые не имели эффекта на рост культуры и не вызывали гибели клеток (рис. 3). Влияние исследуемого препарата на биопленкообразование может осуществляться посредством различных механизмов, поскольку7 процесс формиро- вания биопленок у микобактерий регулируется множеством факторов [Sharma, Petchiappan, Chatter] L 2014]. Одним из возможных механизмов является изменение поверхностных структур клетки, которое, в свою очередь, влияет на способность бактерий формировать биопленки [Aguayo, Bozec,
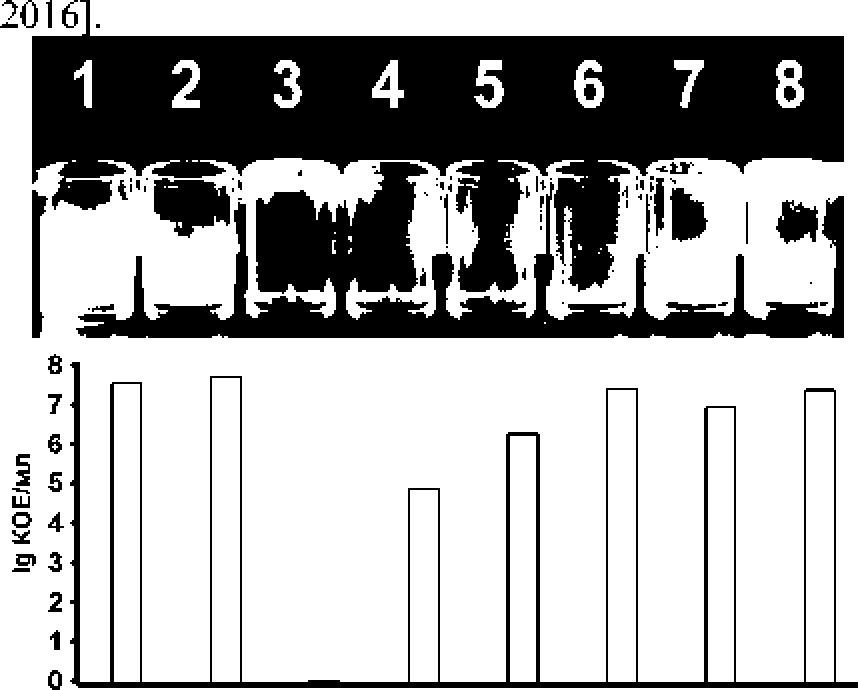
1 2 3 4 5 6 7 В
Рис. 3* Влияние (±)-erogorgiane на биопленко-об ра зовам ие A A smegmatis.
На фото поверхностная биопленка после высушивания. Столбики диаграммы отображают количество КОЕ в лунках. соответствующих лункам на фото, с добавлением Tween 80.
Концентрация антибиотика: 1:2 - контроль без антибиотика. 3 - 0.594 мг/мл: 4 - 0.297: 5 - 0.148 мг/мл: 6 -0,074 мг/мл; 7 - 0.037 мг/мл; 8 - 0.019 мг/мл
Отсутствие действия препарата на клетки гра-мотрицательных Е. с oh свидетельствует о том, что возможной мишенью для действия антибиотика является клеточная стенка грамположительных бактерий. В пользу этого предположения также свидетельствуют и данные, полученные с помощью атомно-силовой микроскопии. Установлено, что под действием сублетальных концентраций (±)-erogorgiane происходит статистически достоверное возрастание шероховатости поверхности клеток А/. smegmatis по отношению к контролю (таблица).
Изменение шероховатости клеточной поверхности под действием антибиотика
|
Вариант опыта |
Шероховатость Sq, нм |
|
Контроль |
153.8 (±7,5) |
|
Сублетальная концентрация (+)-erogorgiane (0,297 мг/мл) |
217.0 (±21.6) * |
Приведены средние значения шероховатости ± с.
*- статистически значимое отличие от контроля без добавки антибиотика. Достоверность различий определяли с помощью критерия Ньюмена—Кейлса.
Предполагалось, что следующим этапом работы должен стать анализ мутантов, устойчивых к действию (+)-erogorgiane. Однако наши попытки получить устойчивые клоны высевом большого количества клеток на твердую среду, содержащую высокие концентрации препарата [Kern, Oethinger, Jellen-Ritter, et al. 2000], не дали результатов. Возможно, это связано с особенностями действия данного антибиотика. Можно предположить, что мутаций в одном или двух генах недостаточно, чтобы микроорганизм стал нечувствителен к данному соединению, как это имеет место в отношении других антибиотиков [McMurry, McDermott, Lexy, 1999; Telenti. Honore, Bernasconi etal., 1997].
Заключение
Таким образом, искусственно синтезированный аналог природного антибиотика (+)-erogorgiane оказывает бактерицидное действие на клетки М smegmatis независимо от стадии роста культуры, а также на нерастущие клетки. Данное вещество в сублетальных концентрациях препятствует образованию биопленок микобактериями.
Несмотря на то, что на данном этапе исследований нельзя точно указать конкретный механизм действия (±)-erogorgiane на клетки микобактерий, нами установлено, что его действие приводит к изменению поверхностных структур клетки.
Работа выполнена при поддержке РНФ (проект 15-13-00092).
Список литературы Изучение механизмов противомикробного действия соединения (+)-erogorgiane
- Aguayo S., Bozec L. Mechanics of bacterial bells and initial iurface colonisation//Advances in Experimental Medicine and Biology. 2016. Vol. 915. P. 245-260
- Arai M., Niikawa H., Kobayashi M. Marine-derived fungal sesterterpenes, ophiobolins, inhibit biofilm formation of Mycobacterium species//Journal of Natural Medicines. 2013. Vol. 67, № 2. P. 271-275
- De Souza M. V.N. Marine natural products against tuberculosis//The Scientific World Journal. 2006. Vol. 21, № 6. P. 847-861
- Kern W.V., Oethinger M., Jellen-Ritter A.S., Levy S.B. Non-target gene mutations in the development of fluoroquinolone resistance in Escherichia coli//Antimicrobial Agents and Chemotherapy. 2000. Vol. 44, № 4. P. 814-820
- Leisner J.J., Jorgensen N.O., Middelboe M. Predation and selection for antibiotic resistance in natural environments//Evolutionary Applications Journal. 2016. Vol. 9, № 3. P. 427-434
- McMurry L.M., McDermott P.F., Levy S.B. Genetic evidence that InhA of Mycobacterium smegmatis is a target for triclosan//Antimicrobial Agents and Chemotherapy. 1999. Vol. 43, № 3. P. 711-713
- Migliori G.B., Sotgiu G., Gandhi N.R. et al. Drug resistance beyond extensively drug-resistant tuberculosis: individual patient data meta-analysis//European Respiratory Journal. 2013. Vol. 42, № 1. P. 169-179
- Sharma I.M., Petchiappan A., Chatterji D. Quorum sensing and biofilm formation in mycobacteria: role of c-di-GMP and methods to study this second messenger//International Union of Biochemistry and Molecular Biology Life. 2014. Vol. 66, № 12. P. 823-834.
- Syal K., Maiti K., Naresh K. et al. Synthetic arabino-mannan glycolipids impede mycobacterial growth, sliding motility and biofilm structure//Glycoconju-gate Journal. 2016. № 4
- Telenti A., Honore N., Bernasconi C. et al. Genotypic assessment of isoniazid and rifampin resistance in Mycobacterium tuberculosis: a blind study at reference laboratory level//Journal of Clinical Microbiology. 1997. Vol. 35, № 3. P. 719-723
- World Health Organisation. Global Tuberculosis report 2015. France, 2015. 192 p
