Изучение окислительных процессов в ткани сердца крыс при моделировании дефицита оксида азота

Автор: Медведев Д.В., Звягина В.И.
Журнал: Сибирский журнал клинической и экспериментальной медицины @cardiotomsk
Рубрика: Лабораторные и экспериментальные исследования
Статья в выпуске: 2 т.28, 2013 года.
Бесплатный доступ
Цель исследования: изучение влияния снижения содержания метаболитов оксида азота под действием L-NG-нитроаргинина метилового эфира (L-NAME) - неселективного ингибитора NO-синтазы - на активность сукцинатдегидрогеназы и супероксиддисмутазы и скорость протекания процессов спонтанного окисления белков в сердце крыс. Показано, что L-NAME дозозависимо и с разной интенсивностью снижает уровень метаболитов оксида азота в сыворотке крови и сердце крыс (в сыворотке сильнее, чем в сердце). Ингибирование синтеза оксида азота под действием L-NAME приводит к дозозависимому уменьшению спонтанного окисления белков в сердце крыс, одной из причин которого, возможно, является изменение активности митохондриальных ферментов.
Оксид азота, l-ng-нитроаргинина метиловый эфир (l-name), окислительная модификация белков, супероксиддисмутаза, сукцинатдегидрогеназа, митохондрии
Короткий адрес: https://sciup.org/14919855
IDR: 14919855 | УДК: 577.1:616.127-008.9
Study of the oxidative processes in rat heart tissue in nitric oxide deficit modeling
The purpose of this study was to investigate the effects of decrease in the nitric oxide metabolite content, induced by a nonselective inhibitor of NOsynthase, LNGnitroarginine methyl ester (LNAME), on the activity of succinate dehydrogenase and superoxide dismutase and on the rate of spontaneous protein oxidation in the rat heart. Data showed that the treatment with LNAME dosedependently decreased the level of nitric oxide metabolites in blood serum and, to a lesser degree, in the rat heart. Inhibition of nitric oxide synthesis by LNAME caused a dosedependent decrease in spontaneous protein oxidation in the rat heart due to, perhaps, the changes in the activity of mitochondrial enzymes.
Текст научной статьи Изучение окислительных процессов в ткани сердца крыс при моделировании дефицита оксида азота
Оксид азота (NO) является вторичным мессенджером, аутокринным и паракринным регулятором. В настоящее время механизм действия и биологические эффекты оксида азота активно изучаются. Известно, что NO активирует гемсодержащий фермент – растворимую гуанилат-циклазу (ГЦ). В частности, в сердечной мышце крыс оксид азота может вызывать увеличение синтеза циклического гуанозинмонофосфата (цГМФ) гуанилатциклазой более чем в 23 раза [13]. Помимо этого он способен взаимодействовать с гемами других протеинов, тиолами, белками, углеводами, ионами металлов и т.д., локализованными в разных тканях и органах [3]. Столь высокая реакционная способность создает предпосылки для проявления оксидом азота разнообразных биологических эффектов.
Оксид азота синтезируется из аргинина NO-синтазой (NOS). Существуют 3 изоформы этого фермента: NO-синтаза 1-го типа (нейрональная, nNOS), NO-синтаза 2-го типа (индуцибельная, iNOS), NO-синтаза 3-го типа (эндотелиальная, eNOS). Нейрональная и эндотелиальная NOS – конститутивные формы NO-синтазы, т.е. их экспрессия постоянна. Их активация осуществляется ионами Са2+. В то же время NO-синтаза 2-го типа индуцируется под влиянием воспалительных цитокинов, ее активация не зависит от ионов Са2+ [7]. Изоформы NOS в значительной степени гомологичны, причем для C-концевого домена гомология характерна не только для самих изоферментов, но и для цитохрома P-450 [3].
Ряд патологических состояний, например, сердечнососудистые, инфекционные, воспалительные заболевания, тромбозы, злокачественные опухоли, заболевания мочеполовой системы, мозговые повреждения при инсультах и др. могут быть связаны с недостатком или гиперпродукцией оксида азота в организме [3]. Недостаток NO играет важную роль в патогенезе сердечно-сосудистых заболеваний. Однако в большинстве случаев исследования ограничиваются изучением участия оксида азота в регуляции сосудистого тонуса. В то же время влияние снижения синтеза NO на сердце, наблюдаемое при ряде патологических состояний, остается недостаточно изученным. Особенно это касается биохимических показателей самих клеток сердца.
Немаловажно изучение участия NO в процессах перекисного окисления липидов и белков. Одной из объективных методик оценки окислительной модификации белков (ОМБ) является спектрофотометрическое определение количества карбонильных групп и иминогрупп в белках по реакции образования динитрофенилгидра- зонов. На сегодняшний день ОМБ признана одним из наиболее ранних и стабильных показателей поражения различных тканей организма при свободно-радикальной патологии [1].
Дыхательная цепь митохондрий является одним из главных источников активных форм кислорода (АФК) в клетках [6]. Особую роль генерация АФК этими органеллами играет для клеток сердца, т.к. в кардиомиоцитах много митохондрий, и в них активно протекают процессы аэробного окисления, ведь окислительное фосфорилирование – главный источник энергии для сердечной мышцы. Поэтому процессы свободно-радикального окисления в клетках сердца можно объяснять с позиций изменения активности митохондриальных оксидоредуктаз. С этой целью в данной статье рассматривается изменение активности супероксиддисмутазы (СОД) как одного из главных ферментов антиоксидантной защиты и сукцинатдегидрогеназы (СДГ) как важного связующего звена в процессах аэробного окисления, а именно между циклом трикарбоновых кислот и дыхательной цепью митохондрий.
Цель: изучить влияние снижения содержания метаболитов оксида азота в сыворотке крови и митохондриях клеток сердца под действием L-NG-нитроаргинина метилового эфира (L-NAME) – неселективного ингибитора NO-синтазы – на активность митохондриальных оксидоредуктаз (СОД и СДГ) и скорость протекания процессов спонтанного окисления белков в сердце.
Материал и методы
Исследование проводилось на 24 крысах-самцах линии Wistar, которые были разделены на три равные группы. Крысам первой (контрольной) группы (n=8) внутрибрюшинно вводился 0,9 %-й раствор хлорида натрия (NaCl) 1 раз в сутки в течение 7 дней, второй (n=8) – внутрибрюшинно вводился препарат L-NAME (L-NG-нит-роаргинина метиловый эфир) – неселективный ингибитор NO-синтазы в виде 0,5%-го раствора 1 раз в сутки в течение 7 дней из расчета 25 мг препарата на 1 кг массы тела крысы, а третьей (n=8) – внутрибрюшинно вводился L-NAME в дозе 200 мг на 1 кг также 1 раз в сутки в течение 7 дней. Дозы L-NAME были выбраны в соответствии с литературными данными [11, 14]. Немедленно после забоя животных (на 8-й день эксперимента) брали кровь и сердце. Сыворотку крови использовали для определения содержания в ней метаболитов NO. Из ткани сердца получали гомогенат и выделяли из него митохондрии методом дифференциального центрифугирования [10]. Для оценки окислительной модификации белков использовали надосадочную жидкость, а осадок, содержащий митохондрии, ресуспендировали в 0,25 М растворе сахарозы с добавлением детергента Тритона Х-100 (для разрушения митохондриальных мембран) и далее использовали для определения активности митохондриальных ферментов: СДГ и митохондриальной Mn-зависимой СОД, а также для измерения концентрации метаболитов NO в митохондриях.
Общее содержание белка в пробах определяли по методу Лоури с помощью стандартизированного набора
DiaSyS Diagnostic Systems. Окислительную модификацию белков оценивали по методу R.L. Levine [12] в модификации Е.Е. Дубининой [5], который основан на реакции взаимодействия карбонильных групп и иминогрупп окисленных аминокислотных остатков с 2,4-динитрофенил-гидразином (2,4-ДНФГ) с образованием 2,4-динитрофе-нилгидразонов, обладающих специфическим спектром поглощения в ультрафиолетовой и видимой областях спектра. При этом образуются альдегид-динитрофенил-гидразоны (А-ДНФГ) и кетон-динитрофенилгидразоны (К-ДНФГ). А-ДНФГ являются ранним маркером окислительной деструкции белков, а КДНФГ – поздним [4]. Оптическую плотность образовавшихся динитрофенилгид-разонов регистрировали на спектрофотометре СФ-2000 при длинах волн 254, 270, 280, 356, 363, 370, 430 и 530 нм.
Активность СДГ исследовали с помощью метода, основанного на определении восстановленного гексацианоферрата [10]. Активность СОД определяли при помощи метода В.А. Костюка [8].
Определение метаболитов NO (нитритов и нитратов) проводили в митохондриальной фракции гомогената сердечной мышцы и в сыворотке крови с помощью метода в модификации В.А. Метельской [9] на иммунофер-ментном анализаторе StatFax 3200.
Полученные в ходе исследования результаты обрабатывались с помощью программы Microsoft Excel, 2003. Для определения различий между независимыми группами (контрольной и получавшими L-NAME) использовали U-критерий Манна–Уитни. Уровень отличий рассматривался как статистически значимый при вероятности ошибки (p<0,05).
Результаты и обсуждение
Введение L-NAME статистически достоверно (p<0,01) приводит к дозозависимому снижению содержания метаболитов оксида азота как в сыворотке крови, так и в митохондриальной фракции гомогената сердца крыс (табл. 1).
Однако это снижение более выражено в сыворотке, чем в митохондриях. Это может быть обусловлено как особенностями распределения L-NAME в организме, так и некоторой селективностью его действия в отношении ингибирования разных изоферментов NOS. Так, L-NAME несколько сильнее ингибирует конститутивные формы NOS (nNOS, eNOS), чем индуцибельную [3]. Основным поставщиком NO для крови является eNOS эндотелия сосудов. В сердце присутствуют все 3 изоформы NО-син-таз: nNOS локализуется в саркоплазматическом ретикулуме и митохондриях клеток сердца, eNOS – в инвагинациях плазматической мембраны кардиомиоцитов (каве-олах), iNOS – в кардиомиоцитах вдоль сократительных волокон, в их плазматических мембранах, включая T-ту-булы, ядерную оболочку, митохондрии и комплекс Гольджи, а также в макрофагах, фибробластах, эндотелиоци-тах коронарных сосудов и эндокарда, гладкомышечных клетках коронарных сосудов [2]. Причем iNOS экспрессируется в клетках сердца как при патологии, так и в норме. Учитывая, что активность iNOS в сотни раз выше, чем
Таблица 1
Концентрация метаболитов NO в сыворотке крови и митохондриях сердца, мкмоль/л (результаты представлены в форме: среднее значение±стандартное отклонение, M±s)
|
Крысы, получавшие в/б 0,9%-й раствор NaCl в течение 7 дней |
Крысы, получавшие в/б L-NAME в дозе 25 мг/кг в течение 7 дней |
Крысы, получавшие в/б L-NAME в дозе 200 мг/кг в течение 7 дней |
|
|
Концентрация метаболитов NO в сыворотке крови, мкмоль/л Концентрация метаболитов NO в митохондриях сердца, мкмоль/л |
111,8±9,09 96,6±19,33 |
74,6±8,56** ( ↓ 33,28%) 78,0±13,24** ( ↓ 19,2%) |
31,7±12,88** ( ↓ 71,63%) 52,8±13,81** ( ↓ 45,33%) |
Примечание: ** – p<0,01 – по сравнению с контрольной группой.
контрольная группа
L-NAME 25 мг/кг
L-NAME 200 мг/кг сдг
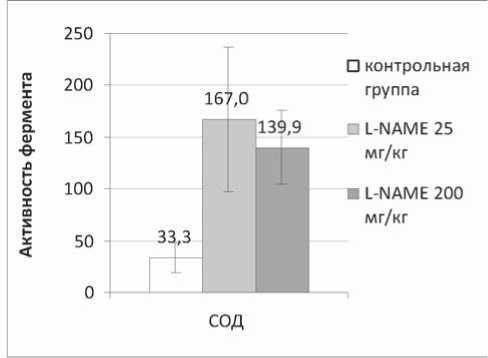
Рис. 2. Изменение активности СОД (оптическая плотность, у.е./ мг белка) в сердце под действием L-NAME в дозах 25 мг/кг и 200 мг/кг (p<0,05 – по сравнению с контрольной группой)
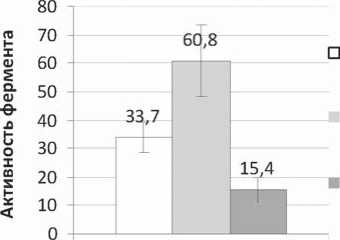
Рис. 1. Изменение активности СДГ (нмоль сукцината/мин на г белка) в сердце под действием L-NAME в дозах 25 мг/кг и 200 мг/кг (p<0,05 – по сравнению с контрольной группой)
у конститутивных форм NO-синтазы, она, по-видимому, является главным продуцентом NO в кардиомиоцитах. Поэтому не столь значительное снижение содержания метаболитов NO в митохондриях клеток сердца по сравнению с сывороткой крови можно объяснить менее выраженным ингибированием iNOS под действием L-NAME.
Следует также отметить, что сам факт ингибирования синтеза NO в митохондриях сердца указывает на наличие в этом органе эстераз, обеспечивающих гидролиз L-NAME как сложного эфира до его активного метаболита L-NG-нитроаргинина (L-NNA).
L-NAME вызывает значительное изменение активности митохондриальных ферментов, которые отражены на рисунках 1 и 2.
L-NAME повышает активность митохондриальной Mn-зависимой СОД, причем при введении ингибитора в дозе 25 мг/кг активность фермента повышается более чем в 4 раза, а в дозе 200 мг/кг – в 3,2 раза. Активность СДГ изменяется разнонаправленно: доза 25 мг/кг приводит к увеличению активности фермента на 80,39%, а доза 200 мг/кг – к ее снижению на 54,28%.
Повышение активности СДГ можно объяснить с позиций ингибирования оксидом азота ферментов цикла Кребса (аконитатгидратазы) и дыхательной цепи митохондрий (цитохром-с-оксидазы) [3]. Снижение синтеза NO под действием L-NAME в дозе 25 мг/кг, по-видимому, приводит к повышению скорости реакций цикла трикар- боновых кислот и дыхательной цепи митохондрий из-за снижения ингибирующего действия оксида азота. Понижение активности СДГ в результате более выраженного ингибирования синтеза NO после введения L-NAME в дозе 200 мг/кг может быть связано с нарушением коронарного кровотока и доставки к миокарду кислорода и, как следствие, с замедлением окислительных процессов в кардиомиоцитах. Повышение активности митохондриальной СОД под действием L-NAME в обеих дозах, возможно, также является следствием нарушения коронарного кровотока с последующей реперфузией. L-NAMЕ является обратимым ингибитором NOS, и вполне возможно, что при введении этого препарата 1 раз в сутки синтез NO успевает частично восстановиться, а это, в свою очередь, приводит к временному восстановлению коронарного кровотока. Таким образом, можно предположить, что повышение активности митохондриальной СОД является механизмом адаптации к стрессу, и в данном случае это не следует рассматривать как однозначно положительное влияние L-NAME на состояние антиоксидантной системы. Кроме того, необходимо учитывать, что под действием СОД из супероксидного анион-радикала образуется перекись водорода [6], также являющаяся токсическим соединением, способным вызывать дисбаланс системы антиоксидантной защиты.
Введение L-NAME в дозе 25 мг/кг не приводит к статистически значимому изменению содержания белка в
Таблица 2
Концентрация белка в гомогенате ткани сердца, лишенном митохондрий, мг/мл (М±s)
|
Крысы, получавшие в/б 0,9%-й раствор NaCl в течение 7 дней |
Крысы, получавшие в/б L-NAME 25 мг/кг в течение 7 дней |
Крысы, получавшие в/б L-NAME 200 мг/кг в течение 7 дней |
|
|
Общий белок, мг/мл гомогената ткани сердца |
4,5±1,12 |
3,9±1,71 ( ↓ 15%) |
6,3±1,02* ( ↑ 39%) |
Примечание: * – p<0,05 – по сравнению с контрольной группой.
Длина волны, нм
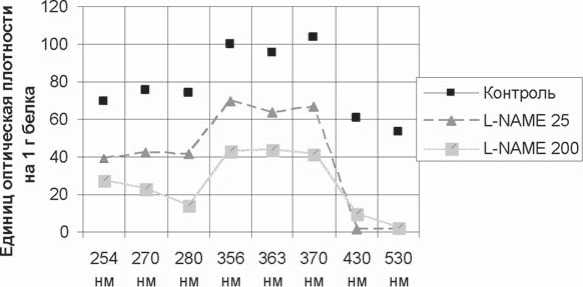
Рис. 3. Показатели спонтанной ОМБ
сердце, а доза 200 мг/кг достоверно его увеличивает на 39% (табл. 2).
При этом речь, конечно, идет только о белках, растворимых в 0,25 М растворе сахарозы, т.е. о саркоплазматических белках. Как известно, повышение содержания в сердце этих белков обычно наблюдается при патологии этого органа.
Изучение окислительной модификации белков в без-митохондриальной фракции гомогената сердца крыс после ингибирования синтеза оксида азота под действием L-NAME в дозах 25 и 200 мг/кг дало результаты, представленные на рисунке 3.
Введение L-NAME в дозе 25 мг/кг достоверно (p<0,05 – по сравнению с контрольной группой) снижает показатели спонтанной ОМБ при длине волны 430 нм (соответствует основным альдегид-динитрофенилгидразонам (АДНФГ) и кетон-динитрофенилгидразонамов (КДНФГ)) в 49,5 раз. L-NAME же в дозе 200 мг/кг достоверно снижает значения спонтанной ОМБ при λ =356 нм (что соответствует нейтральным АДНФГ) на 38,2% (p<0,01 – по сравнению с контрольной группой), а при 363 нм (что соответствует нейтральным КДНФГ) – на 31,3% (p<0,05 – по сравнению с контрольной группой).
Исходя из полученных при изучении ОМБ данных, можно заключить, что влияние L-NAME на процессы окисления белков в клетках сердца сильно зависит от дозы. Введение препарата в количествах 25 и 200 мг/кг в течение 7 дней дозозависимо замедляет процессы спонтанного окисления белков, практически не влияя на соотношение альдегидных и кетоновых производных протеинов (т.к. почти не изменяется спектр поглощения).
Снижение спонтанного окисления белков в сердце можно объяснить с разных позиций. Во-первых, такое изменение может быть связано со снижением концентрации метаболитов оксида азота; ведь NO сам по сути является свободным радикалом (содержит неспаренный электрон), а при взаимодействии с супероксидным анион-радикалом (O2·–) образует сильнейший окислитель – пероксинитрит (ОNOO–) [6]. Во-вторых, снижение окисления белков в сердечной мышце можно объяснить, исходя из изменения активности митохондриальных ферментов. Так, L-NAME в обеих дозах (25 и 200 мг/кг) вызывает выраженное увеличение активности митохондриальной СОД, что неизбежно должно снижать в клетке концентрацию O2·–, а следовательно, и ОNOO–. Более выраженное уменьшение скорости процессов спонтанного окисления белков при введении L-NAME в дозе 200 мг/кг, чем в дозе 25 мг/кг логично вытекает из изменения активности СДГ. Как уже отмечалось, дыхательная цепь митохондрий является важным источником генерации АФК в кардиомиоцитах. Ее активность непосредственно связана с активностью ферментов цикла трикарбоновых кислот, поставляющего доноры водорода для дыхательной цепи. Снижение активности СДГ под действием L-NAME в дозе 200 мг/кг должно приводить как к уменьшению скорости ферментативных реакций цикла Кребса, так и к ингибированию митохондриальной цепи переноса электронов. Результатом вышеперечисленного является замедление процессов свободнорадикального окисления в клетках сердца.
Выводы
L-NAME дозозависимо и с разной интенсивностью снижает уровень метаболитов оксида азота в сыворотке крови и сердце крыс (в сыворотке сильнее, чем в ткани сердца).
Ингибирование синтеза оксида азота под действием L-NAME приводит к дозозависимому уменьшению спонтанного окисления белков в сердце крыс, одной из причин которого, возможно, является изменение активности митохондриальных ферментов.
Список литературы Изучение окислительных процессов в ткани сердца крыс при моделировании дефицита оксида азота
- Абаленихина Ю.В. Фомина М.А., Чурилов Г.И. и др. Окислительная модификация белков тимуса крыс под влиянием меди в ультрадисперсной форме//Фундаментальные исследования. -2012. -№ 11. -С. 1315-1319.
- Гарматина О.Ю., Ткаченко М.Н., Мойбенко А.А. Индуцибельная синтаза оксида азота при патологии сердца (обзор литературы и собственных исследований)//Теоретична медицина. -2005. -Т. 11, No. 4. -С. 645-660.
- Граник В.Г., Григорьев Н.Б. Оксид азота (NO). Новый путь к поиску лекарств: монография. -М.: Вузовская книга, 2004. -360 с.
- Губский Ю.И., Беленичев И.Ф., Левицкий Е.Л. и др. Токсико-логические последствия окислительной модификации белков при различных патологических состояниях (обзор литературы)//Современные проблемы токсикологии. -2005. -Т. 8, № 3. -С. 20-27.
- Дубинина Е.Е., Бурмистров С.О., Ходов Д.А. и др. Окислительная модификация белков сыворотки крови человека, метод ее определения//Вопросы мед. химии. -1995. -Т. 41, № 1. -С. 24-26.
- Дубинина Е.Е. Продукты метаболизма кислорода в функциональной активности клеток (жизнь и смерть, созидание и разрушение). Физиологические и клинико-биохимические аспекты. -СПб.: Медицинская пресса, 2006. -С. 276-282.
- Козловский В.И. Механизмы регуляции коронарного кровотока, опосредованной эндотелиальными сосудорасширяющими факторами: монография. -Гродно: ГрГМУ, 2011. -216 с.
- Костюк В.А., Потапович А.И., Ковалева Ж.В. Простой и чувствительный метод определения активности супероксид дисмутазы, основанный на реакции окисления кверцетина//Вопросы мед. химии. -1990. -№ 2. -С. 88-91.
- Метельская В.А., Гуманова Н.Г. Скрининг метод определения уровня метаболитов оксида азота в сыворотке//Клиническая лабораторная диагностика. -2005. -№ 6. -С. 15-18.
- Методы биохимических исследований (липидный и энер гетический обмен)/под ред. М.И. Прохоровой. -Л.: Изд во Ленинградского университета, 1982. -327 с.
- Покровский М.В., Покровская Т.Г., Корчаков В.И. и др. Эндотелиопротекторные эффекты L-аргинина при моделировании дефицита окиси азота//Экспериментальная и клиническая фармакология. -2008. -Т. 71, № 2. -С. 29-31.
- Levine R.L., Garland D., Oliver C.N. et al. Determination of carbonyl content in oxidatively modified proteins//Methods in enzymology. -1990. -Vol. 186. -P. 464-478.
- Murad F. Discovery of some of the biological effects of nitric oxide and its role in cell signaling//Bioscience reports. -1999. -Vol. 19, No. 3. -P. 133-154.
- Zun Yi Wang, Hakanson Rolf. Role of nitric oxide (NO) in ocular inflammation//British J. of Pharmacology -1995. -Vol. 116. -P. 244-245.
