Эффективная схема индукции полиовуляции для трансплантации эмбрионов у коз

Автор: Аксенова П.В., Айбазов А.-М.М., Коваленко Д.В., Садчикова Е.Р., Гольдман И.Л.
Журнал: Сельскохозяйственная биология @agrobiology
Рубрика: Биотехнологии репродукции
Статья в выпуске: 6 т.48, 2013 года.
Бесплатный доступ
Технология трансплантации эмбрионов складывается из ряда последовательных самостоятельных операций. Одним из ее важных элементов служит индукция полиовуляции у животных-доноров. При этом главной проблемой считается вариабельность ответа яичника на обработку. Различия в реакции яичников на эстрогенные гормоны объясняются изменением эндогенного гормонального фона, который непосредственно влияет на процесс оплодотворения и развитие эмбриона. Нами разработана схема индукции полиовуляции у коз-доноров для трансплантации эмбрионов с высокой степенью синхронизации овуляций (независимо от исходного гормонального фона доноров). Схема предусматривает введение имплантов Крестар на 8 сут; 1-крат-ную инъекцию 500 ед. фоллигона и дробные инъекции 7 мл овагена (с интервалом 12 ч при 1-кратной дозе 1 мл), которые проводят, начиная с 60-го ч до извлечения имплантов; введение 500 ед. хорулона в момент фиксации половой охоты. Применение предложенной схемы дает стабильную полиовуляцию с эффективностью 15,5-17,5 желтых тел и 74,0-90,5 % выхода эмбрионов, пригодных для трансплантации, как в эстральный, так и в анэстральный сезоны.
Полиовуляция, трансплантация, крестар, оваген, хорулон, зааненская порода
Короткий адрес: https://sciup.org/142133461
IDR: 142133461 | УДК: 636.32/38.082.453.5
The effective scheme to induce poliovulation for transplantation of embryos in goats
In embryo transplantation, there are some subsequent manipulations, of which the induction of polyovulation in donors is one of the most important element. At that, the main problem consists in variability of ovary response to treatment. The ovary reaction to the estrogenic hormones differs depending on changes in the endogenous hormonal pattern which can influence directly the fertilization and embryo development. The suggested scheme can be used to induce the polyovulation in donor goats for embryo transplantation with highly synchronized ovulalation that does not depend on an initial hormonal pattern of the donor. According to the scheme, Krestar implants are being used for 8 days. Folligon must be injected in a dose of 500 U. The fractional injections of Ovagene (7 ml in total, by 1 ml each 12 hours) are conducted starting from the 60 hours before the implants are removed. At fixation of estrus the animals were injected with Chorulon in a dose of 500 U. This scheme both in estral and unestral seasons results in a stable polyovulation with the efficiency of 15.5-17.5 yellow bodies and 74.0-90.5 % output of embryos good for transplantation.
Текст научной статьи Эффективная схема индукции полиовуляции для трансплантации эмбрионов у коз
Технология трансплантации эмбрионов складывается из нескольких элементов, причем каждый представляет собой самостоятельную операцию. Первая из них — это подбор доноров, у которых стимулируется множественная овуляция. Теоретические принципы индукции полиовуляции, разработанные еще М.М. Завадовским в 1935 году с целью повышения плодовитости овец, получили продолжение в работах отечественных и зарубежных ученых (1).
В первых экспериментах для стимуляции роста фолликулов и контроля за овуляцией использовались гонадотропные препараты, полученные из мочи беременных женщин (хорионический гонадотропин человека — ХГЧ, пролан) (2, 3). Однако другие исследователи выявили отрицательные эффекты ХГЧ: он снижает оплодотворяемость, вызывает преждевременную регрессию желтых тел, повышает эмбриональную смертность, приводит к разновременному приходу животных в охоту и удлинению этого периода (4). Т. Sugie с соавт. (5) в опытах на овцах с целью индукции суперовуляции обрабатывал животных гонадотропином сыворотки жеребых кобыл (ГСЖК) и ХГЧ в дозе 800 ед. У 93 % из 44 обработанных овец было обнаружено от 1 до 36 овуляций (в среднем 7,9 на одного донора). От 31 овцы получили 132 яйцеклетки, или 4,2 на одного донора, 101 из которых были оплодотворенными, однако 50 % клеток оказались непригодными для пересадки (5).
И.Я. Шихов (6) для суперовуляции на 12-е сут полового цикла вводил фоллигон в дозе 1000 И.Е. с последующей обработкой простагландином Ф2-альфа (эстрофан) в дозе 250 мкг. Число овуляций в среднем на
Исследования выполнены в рамках Международной программы «Разработка технологий и организация опытного производства высокоэффективных и биологически безопасных лекарственных средств нового поколения и биологически полноценного детского питания на основе лактоферрина человека, получаемого из молока трансгенных животных, на 2009-2013 г.г.» («БелРосТрансген-2», Россия-Белоруссия»).
одного донора составило 10,2 (с вариабельностью от 0 до 18), на одного положительного донора было получено 6,9 яйцеклетки, из них полноценных 4,7 (6). J.P. Ryan, основываясь на собственных экспериментах, утверждает, что повышение числа нормальных эмбрионов обеспечивается совместным применением ГСЖК и фолликулостимулирующего гормона ФСГ-п (7).
При индукции полиовуляции главной проблемой считается вариабельность ответа яичника на обработку (8-11). Различия в реакции яичников на эстрогенные гормоны объясняют изменением эндогенного гормонального фона, который непосредственно влияет на процесс оплодотворения и развитие эмбриона (12, 13). Также было выяснено, что наличие выраженного желтого тела на момент инъекции гонадотропина гарантирует хорошую реакцию суперовуляции. Наличие в яичнике фолликула одновременно с желтым телом снижает результат гормональной обработки. В этих случаях в отношении степени полиовуляции результат часто оказывается отрицательным (14).
Современные исследования по индукции полиовуляции у коз также подтверждают, что индивидуальная изменчивость в реакции яичников отмечается даже при применении стандартных схем и доз гонадотропных препаратов. Более того, ни одна из проверенных и рекомендованных схем гормональной обработки не обеспечивает устойчивого ответа яичников (15, 16). Следовательно, совершенствование этой методики для получения полиовуляции со стабильно высоким процентом гормональных ответов у доноров и достаточным числом полноценных, способных к имплантации эмбрионов остается задачей, которая все еще требует решения.
Нашей целью была разработка оптимальной схемы гормональной обработки коз-доноров для обеспечения контролируемой полиовуляции.
Методика . Исследования осуществляли в условиях опытной станции. В экспериментальные группы отбирали коз зааненской породы 2-3-й лактации. Опыты проводили как в эстральный, так и в анэстральный сезон. Общее число животных составило 68 гол. Для стимуляции полиовуляции применяли препараты производства компании «Intervet» (Нидерланды).
Для пролонгации/имитации лютеиновой фазы и синхронизации половых циклов использовали ушные импланты Крестар (действующее вещество — синтетический аналог прогестерона норгестамет). Имплант вводили в области латеральной поверхности уха в месте рыхлого соединения кожи с хрящом, где отсутствуют крупные кровеносные сосуды. Для этого имплантатором с вложенным в него имплантом делали прокол кожи в зоне ушного кармана и осторожно продвигали иглу параллельно гребню ушной раковины у ее каудального края. После продвижения на 2-4 см иглу немного отводили назад и выталкивали под кожу имплант мандреном. Для прерывания лютеиновой фазы и стимуляции развития фолликулов применяли гонадотропные препараты фоллигон и оваген. Фол-лигон представляет собой ГСЖК, оваген содержит овечий питуитарный ФСГ (NIADDK-oFSG-17). Для индукции одновременных овуляций созревших фолликулов использовали препарат хорулон (действующее вещество — ХГЧ), обладает активностью лютеинизирующего гормона (ЛГ).
Сроки наступления и длительность половой охоты определяли с помощью козлов-пробников. Выборку коз в охоте начинали спустя 12 ч после извлечения имплантов и повторяли каждые 2 ч до окончания периода охоты. Коз в охоте осеменяли интрацервикально свежеполученной неразбавленной спермой согласно инструкции (17).
Количество половых и гонадотропных гормонов в крови подопытных животных в течение эксперимента контролировали, регистрируя периодически концентрацию ЛГ, ФСГ, прогестерона и эстрадиола методом ИФА. Схему экспериментов приводили в соответствие с полученными данными о гормональном фоне коз.
Наличие и качество полиовуляции определяли по результатам осмотра яичников, оценки возраста и полноценности полученных эмбрионов с применением лапароскопии либо лапаротомии. К пригодным для трансплантации относили оплодотворенные ооциты, а также 2- и 4-блас-томерные эмбрионы без нарушений в строении и целостности оболочки, развивающиеся при инкубировании.
При разработке схемы варьировали дозы, сроки и кратность введения препаратов.
Полученные результаты обрабатывали статистически в программе BioStat.
Результаты . Использованный прием синхронизации половых циклов заменяет трудоемкую процедуру выявления особей в охоте с последующим отслеживанием ее длительности и расчета средней индивидуальной продолжительности по каждому животному, как это принято при индукции суперовуляции в скотоводстве. Для синхронизации охоты группу коз фронтально (без учета полового цикла) обрабатывали прогестагеном. При этом у животных, которые находились в фолликулярной фазе, имитировали лютеиновую фазу (у особей, уже находящихся в лютеиновой фазе, она пролонгировалась). Последующее удаление импланта, прерывающее действие прогестагена, обеспечивало получение синхронизированных половых циклов на известной стадии у всех животных группы.
При определении оптимального срока аппликации имплантов (8, 9, 10 и 12 сут, доза — 1,5 мг по действующему веществу, д.в.) выяснили, что ее продолжительность не имела значения. Особи, которым имплант вводили на 8 и 12 сут (крайние значения), в равной степени синхронно приходили в охоту, и при соответствующей гормональной обработке эффективность полиовуляции у них достоверно не различалась. Диапазон концентраций прогестерона у коз, находящихся на разных стадиях полового цикла, начинал выравниваться уже к 5-м сут обработки (рис. 1), и к 78-м сут синхронизация половых циклов наблюдалась во всех сравниваемых группах.
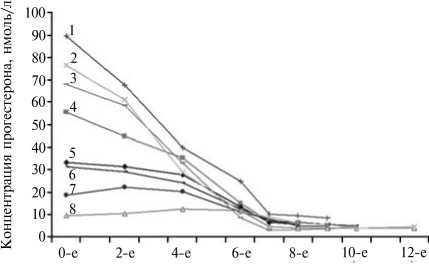
Срок обработки, сут
Рис. 1. Динамика концентрации прогестерона при синхронизации половых циклов у коз зааненской породы при применении имплантов Крестар: 1-8 — особи, исходно находящиеся на разных стадиях полового цикла.
Следовательно, время аппликации ушных имплантов можно уменьшить с 12 до 8 сут, что позволяет существенно сократить срок всей гормональной обработки (на 4 сут, или на 33,3 %).
Далее мы сравнили эффект введения животным ова-гена по двум схемам. В одной группе козы получали препарат дважды в сутки с интервалом 12 ч в течение 4 сут в равных дозах (по 0,8 мл/гол. утром и вечером), во второй — также с интервалом 12 ч в течение 4 сут, но в возрастаю - щей дозировке (по 0,6; по 0,8; по 0,8 и по 1 мл/гол. утром и вечером). Общая доза овагена в обеих группах составила 6,4 мл, или 5,6 мг д.в. на одно животное. Кроме того, всем особям из обеих групп за 60 ч до извлечения импланта вводили по 500 ед. фоллигона и в начале периода половой охоты по 150 ед. хорулона.
|
1. Результаты индукции полиовуляции у коз зааненской породы при применении схемы с постоянной (I группа) и возрастающей (II группа) дозировкой овагена ( M ± m ) |
||
|
Показатель | |
I группа |
| II группа |
|
Всего обработано, гол. |
7 |
9 |
|
Реагировало полиовуляцией, гол/% |
7/100 |
8/100 |
|
Реакция полиовуляции, желтых тел/гол. |
10,67±2,10 |
12,17±3,44 |
|
Положительных по извлечению доноров, гол/% |
9/100,0 |
8/100,0 |
|
Извлечено эмбрионов, шт/гол. |
7,35±1,17 |
8,02±2,22 |
|
В том числе: |
||
|
пригодных для трансплантации |
3,26±0,33 |
3,42±1,15 |
|
непригодных для трансплантации |
4,09±0,56 |
4,58±1,13 |
|
Из них неоплодотворенные яйцеклетки, шт. |
3,64±0,55 |
3,72±1,05 |
|
Выход пригодных эмбрионов, % |
44,4 |
42,6 |
При обеих описанных схемах введения овагена (табл. 1) практически 100 % особей пришли в охоту при показателе полиовуляции 10,7012,17 желтого тела (ж.т.) на животное (р > 0,05). Однако в обоих вариантах высокий процент неоплодотворенных клеток и низкий выход полноценных эмбрионов свидетельствовали о несинхронности овуляций. На яичниках отмечались петехии, имелись желтые тела разного возраста, созревшие, но не лопнувшие фолликулы.
На основании обследования яичников и анализа полученных результатов мы модифицировали описанную схему, увеличив количество хорулона до 300 ед., и сравнили эффект в двух группах животных, из которых в первой особей обрабатывали по предыдущей схеме, во второй — с применением удвоенного количества хорулона (табл. 2). Увеличение дозы хорулона значительно улучшило результаты обработки. Реакция полиовуляции и, главное, выход пригодных для трансплантации эмбрионов достоверно возросли и составили соответственно 12,6 ж.т./гол. и 68,7 %.
2. Результаты индукции полиовуляции у коз зааненской породы при исходной (I группа) и повышенной (II группа) дозе хорулона ( M ± m )
|
Показатель | |
I группа |
| II группа |
|
Всего обработано, гол. |
3 |
6 |
|
Реагировало полиовуляцией, гол/% |
3/100 |
6/100 |
|
Реакция полиовуляции, желтых тел/гол. |
10,60±2,80 |
14,32±1,54 |
|
Положительных по извлечению доноров, гол/% |
3/100,0 |
6/100,0 |
|
Извлечено эмбрионов, шт/гол. |
8,33±2,03 |
12,60±1,25 |
|
В том числе: |
||
|
пригодных для трансплантации |
3,85±1,14 |
8,65±0,49 |
|
непригодных для трансплантации |
4,48±0,90 |
3,95±0,19 |
|
Из них неоплодотворенные яйцеклетки, шт. |
3,00±1,11 |
2,32±0,25 |
|
Выход пригодных эмбрионов, % |
46,1 |
68,7 |
При индукции полиовуляции с применением экзогенных гормональных препаратов важно контролировать концентрацию собственных гормонов животного. Оценив динамику показателей для ЛГ, ФСГ, прогестерона и эстрадиола на фоне гормональной обработки (рис. 2), мы обнаружили, что их значения по сывороточным гормонам у коз при индукции полиовуляции были ниже, чем при естественном половом цикле. Особенно это проявилось в отношении ФСГ, что вступает в некоторое противоречие с высокой активностью фолликулогенеза у обработанных животных. Однако в целом динамика концентрации половых и гонадотропных гормонов соответствовала последовательности введения экзогенных препара- тов и изменениям в организме, обусловленным индукцией искусственного эстрального цикла.
В связи с достаточно низким уровнем ЛГ в сыворотке крови у обрабатываемых коз доза хорулона была еще раз увеличена (до 500 ед.), а время введения препарата изменили (вместо начала охоты его стали вводить через 3 ч после ее начала), причем для ускорения поступления всей дозы к клеткам-мишеням внутримышечное введение заменили на внутривенное. Кроме того, так как в 90 % случаев охота у доноров наступала в промежутке от 12 до 20 ч после извлечения импланта и к последней инъекции овагена животным уже был введен хорулон (то есть необходимость в заключительной инъекции овагена отпадала), однократную дозу препарата увеличили с 0,8 до 1,0 мл, а число введений сократили с 8 до 7. Для
А
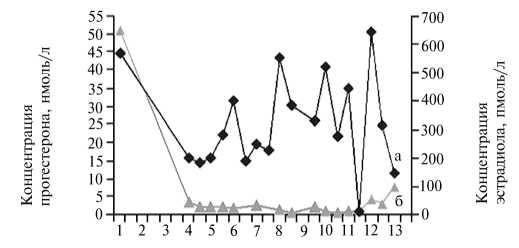
Б
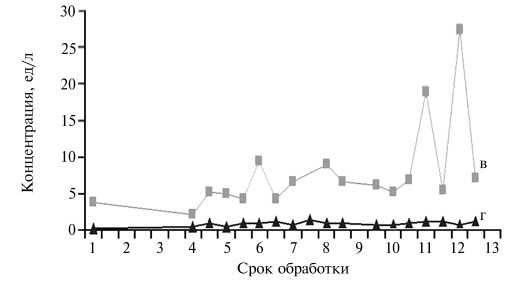
Рис. 2. Динамика концентрации половых (А; а — эстрадиол, б — прогестерон) и гонадотропных гормонов (Б; в — ЛГ, г — ФСГ) в сыворотке крови коз зааненской породы при применении разработанной схемы индукции полиовуляции: 1 — 0-е сут обработки (введение импланта), 2 — 2-е сут, 3 — 4-е сут, 4 — 6-е сут, 5 — оваген + фоллигон, 6 — извлечение импланта, 7 — за 3 ч до охоты, 8 — начало охоты, 9 — 3 ч от начала охоты, 10 — 5 ч от начала охоты, 11 — 7 ч от начала охоты, 12 — 10 ч от начала охоты, 13 — конец охоты.
одновременного оплодотворения вышедших в яйцевод ооцитов вместо 3-4-кратного осеменения доноров с начала охоты, использовали только 2-кратное — через 6 и 10 ч после ее начала.
По описанной схеме были обработаны доноры в четырех экспериментах (в эстральный и анэст-ральный сезоны года). Проведенные нами опыты показали достаточно однородные результаты (табл. 3).
Все козы пришли в охоту. Состояние половой охоты характеризовалось наличием и полноценным проявлением всех ее феноменов (положительная локомоторная реакция на самца, течка, рефлекс неподвижности).
Продолжительность периода охоты в среднем составила 22-28 ч, время от извлечения импланта до начала охоты — 14,5-19 ч. При этом
отмечали определенную закономерность: чем меньше времени от извлечения импланта до начала охоты, тем больше ее продолжительность.
Реакция полиовуляции составила 15,52-17,5 ж.т./гол., при этом доля клеток, пригодных для трансплантации, была очень высока и составляла 74,3-90,4 % от всех извлеченных.
При этом не наблюдалось перестимуляции яичников, не было кист, созревших, но не лопнувших фолликулов. Размеры яичников составляли
2,80±0,26х2,12±0,20 см при диаметре желтых тел 4-6 мм (рис. 3).
-
3. Результаты получения пригодных для трансплантации эмбрионов у коз зааненской породы при применении разработанной схемы индукции полиовуляции ( M ± m )
Число доноров
Время от извлечения импланта до начала охоты, ч
Продолжительность охоты, ч
Реакция полиовуляции, число желтых тел
Вымыто клеток, в среднем на 1 гол.
Из них пригодных для трансплантации, шт/%
|
I |
опыт |
||||
|
7 |
18,70±1,34 |
22,75±1,73 II |
17,34±1,55 опыт |
12,33±1,26 |
11,34±1,61/83,3 |
|
5 |
17,50±1,81 |
21,92±2,49 III |
15,52±1,25 опыт |
12,79±1,35 |
9,50±0,65/74,3 |
|
8 |
16,60±1,61 |
23,58±2,17 IV |
17,50±2,01 опыт |
12,50±1,99 |
10,64±1,89/84,0 |
|
6 |
14,50±0,81 |
27,83±2,69 |
16,00±2,35 |
13,83±2,74 |
13,00±3,09/90,4 |
Примечание. Опыты проводились в разные годы на разном поголовье в эстральный и анэстральный сезон.
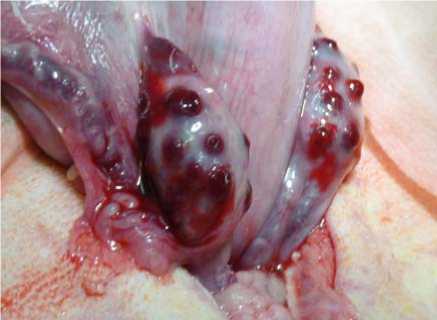
Рис 3. Яичники с желтыми телами при индукции полиовуляции у коз зааненской породы.
Таким образом, нами разработана схема индукции полиовуляции у коз. Она предусматривает введение иплан-тов Крестар на 8 сут; 1-кратную инъекцию 500 ед. фол-лигона и дробные инъекции 7 мл овагена с интервалом 12 ч при 1-кратной дозе 1 мл (начало инъекций — за 60 ч до извлечения имплантов); введение 500 ед. хорулона в момент фиксации половой охоты. Предложенная схема дала хорошие результаты полиовуляции с высокой степенью синхронизации овуляций, о чем свидетельствует высокий процент полученных одновозрастных клеток, пригодных для трансплантации.
