Эффекты введения конъюгата ß-эндорфина с гистоном Н2а на поведенческие и вегетативные показатели у крыс

Автор: Толпыго С.М., Лагутина Л.В.
Журнал: Академический журнал Западной Сибири @ajws
Рубрика: Медицина
Статья в выпуске: 2 (69) т.13, 2017 года.
Бесплатный доступ
Проведена оценка эффектов введения конъюгата ß-эндорфина (ß-Э) с гистоном Н2а на поведенческие и вегетативные показатели у крыс. Показана высокая биологическая активность синтезированного конъюгата ß-Э с гистоном Н2а. Обнаружено, что изменения артериального давления, температуры тела и суммарной двигательной активности при периферическом и внутримозговом введениях конъюгата ß-Э с гистоном Н2а аналогичны эффектам ß-Э, введенного в боковые желудочки мозга. Выявлено сходство действия периферического и внутримозгового введений конъюгата ß-Э с гистоном Н2а, выражающееся в длительных фазными изменениями болевой чувствительности, потребления пищи. Выявлено, что распределение конъ- югата меченого по 125I бета -Э с гистоном Н2а в органах и тканях у крыс существенно отличается от распределения нативного 125I-ß-Э. Содержание меченого конъюгата в мозге в 3-4 раза превышает содержание нативного ß-Э, что свидетельствует о его повышенной проницаемости через гемато-энцефалический барьер.
ß-эндорфин, конъюгат ß-эндорфина, гистон н2а, поведение, вегетативные показатели
Короткий адрес: https://sciup.org/140221984
IDR: 140221984
Effects of administration conjugate ß-endorphin with histone H2a on behavioral and vegetative indices in rats
The effects of administration conjugate ß-endorphin (ß-E) with histone H2a on behavioral and vegetative indices in rats were evaluated. The high biological activity of the synthesized ß-E conjugate with histone H2a is shown. It was found that changes in blood pressure, body temperature and total motor activity in intravenous and intracerebral injections of ß-E conjugate with histone H2a are analogous to ß-E effects introduced into lateral ventricles of the brain. The similarity in action of peripheral and intracerebral injections of ß-E conjugate with histone H2a is revealed, which is expressed in long-term phase changes in pain sensitivity, food intake. It was found that the distribution of conjugate of the labeled 125I ß-E with histone H2a in organs and tissues in rats differs significantly from the distribution of native 125I-ß-E. The content of labeled conjugate in brain is 3-4 times higher than content of native ß-E, which indicates its increased permeability through the blood-brain barrier.
Текст научной статьи Эффекты введения конъюгата ß-эндорфина с гистоном Н2а на поведенческие и вегетативные показатели у крыс
Продукт процессинга проопиомеланокортина в-эндорфин (в-Э) является одним из представителей семейства опиоидных пептидов и распространен в ЦНС, желудочно-кишечном тракте, содержится в сыворотке крови и спинномозговой жидкости животных и человека [3, 5, 8]. Спектр физиологической активности в-Э чрезвычайно широк. Показано, что в-Э является мощным анальгетиком, изменяет деятельность сердечно-сосудистой и дыхательной систем, температуру тела, вызывает разнонаправленные сдвиги в секреции гормонов и обмене медиаторов, а также в значительной степени влияет на различные формы поведения (пищевое, половое, оборонительное и др.) и двигательную активность животных, оказывает положительное эмоциоген-ное действие [5, 6, 8]. Однако современные представления о физиологическом значении в-Э еще далеки от окончательных. При этом, особую сложность представляет оценка соотношения центральных и периферических механизмов, обуславливающих эффекты действия в-Э.
Имеются экспериментальные данные, что катионные белки (гистоны, трансферрин и др.) с изоэлектрической точкой при рН от 10,5 до 12 характеризуются высокой проницаемостью через гематоэнцефалический барьер (ГЭБ). В ряде исследований выявлено также, что конъюгирование регуляторных пептидов с катионизированными белками облегчает их прохождение через ГЭБ за счет рецепторного или абсорбционного трансцитоза, обусловленного электростатическими взаимодействиями конъюгата этих молекул с эндотелием капилляров мозга [7, 9]. Гистон Н2а - один из ядерных белков хроматина (молекулярный вес 15000 д) за счет значительного содержания лизина и аргинина (суммарно 10-11 мол%) проявляет сильно основные свойства, участвует в упаковке нитей ДНК в ядре и в эпигенетиче ской регуляции таких внутриклеточных процессов, как транскрипция, репликация и репарация ДНК, играющих важную роль в механизмах сигнальной трансдукции эндогенных биорегуляторов [4].
В связи с этим представляет значительный интерес исследование физиологической активности своеобразного «химерного» соединения - конъюгата в-Э с катионным белком гистоном Н2а.
Методика.
Эксперименты были выполнены на 64 крысах-самцах популяции "Вистар" с исходной массой тела 250-300 г.
Была разработана оригинальная схема синтеза и синтезирован конъюгат опиоидного пептида β-Э с катионным белком-носителем гистоном Н2а. Конъюгат β-Э с гистоном Н2а получали с помощью бифункционального связывающего соединения глутарового альдегида. Проведен анализ физикохимических свойств полученного конъюгата. Степень связывания пептида с носителем определяли с применением хроматографического анализа кислотных гидролизатов конъюгата и носителя, обработанного связующим соединением, по изменению аминокислотного состава конъюгата по сравнению с контролем, а также по включению меченых по 125I пептидов. Количество связанного β-Э в полученном конъюгате составляло 3-4 молекулы на 1 молекулу гистона.
Животным опытных групп (по 8 крыс) вводили конъюгат β -Э с гистоном Н2а внутривенно (в/в; 1-я группа) и в боковые желудочки мозга (в/ж; 2-я группа). Доза β -Э, связанного с носителем составила 100 мкг/кг и 5 мкг в объеме 5 мкл на животное соответственно. Животным контрольных групп (по 8 крыс) осуществляли в/в и в/ж введение: нативного β -Э (3-я и 4-я группы) в тех же дозах; гистона Н2а, обработанного глутаровым альдегидом, в дозах 100 мкг/кг и 5 мкг в объеме 5 мкл на животное (5-я и 6я группы); физиологического раствора (7-я и 8-я группы).
У всех животных опытных и контрольных групп динамически регистрировали суточное потребление пищи и воды, массу тела.
В фоновый период двукратно и ежедневно после введения исследовали порог болевой чувствительности (tail-flick test). При определении порога болевой чувствительности использовали tail-flick test в модификации [1], основанный на измерении латентного периода реакции отдергивания хвоста крысы при тепловом раздражении сфокусированным световым лучом. Температура луча на поверхности хвоста равна 60о.
____Zl
I % к NaCI /
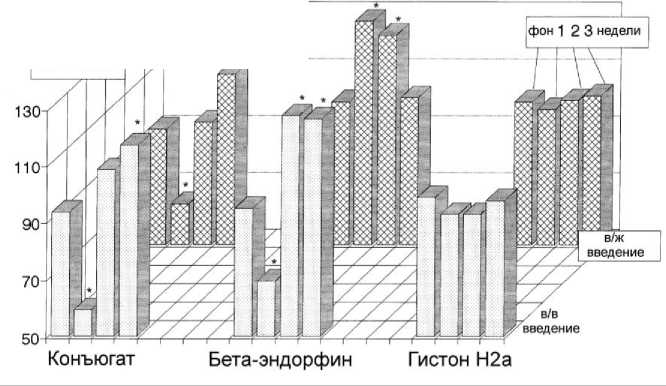
Рис. 1. Динамика среднесуточного потребления пищи в течение эксперимента. * - p < 0,05 по сравнению с фоновым периодом.
Измерение артериального (систолического) давления (АД) у крыс производили непрямым методом при помощи датчика АР-212 фирмы RTF (Германия) на хвостовой артерии. Температуру тела измеряли ректально.
Трижды за весь период экспериментов (в фоновый период, на 2–3-ий и 22–24-ий день после введения веществ) у всех крыс определяли суммарную (горизонтальную и вертикальную) двигательную активность с помощью прибора "Varimex" фирмы "Columbus Instrument", США.
Локализацию канюль в боковых желудочках мозга определяли фото-экспресс методом на замороженных срезах.
В специальной серии экспериментов после в/в введения конъюгата меченого по 125I β -Э c гистоном Н2а (опытная группа – 6 крыс) и нативного 125I- β -Э (контрольная группа – 6 крыс) оценивали динамику изменения активности проб крови (0,1 мл) в различные сроки после введения (через 5, 15, 30, 45, 60, 90, 120, 150, 180 мин), затем животных забивали путем декапитации и исследовали характер распределения 125I β -Э в органах и тканях по активности стандартных навесок (1 г). Для определения активности использовали жидкостный сцинтилляционный счетчик. Конъюгат меченого по 125I β -Э c гистоном Н2а и нативный 125I- β -Э вводили в объеме 0,1 мл на животное. Активность 10 мкл вводимых растворов составляла 10000 мкКи.
Результаты обрабатывали статистически с использованием t-критерия Стьюдента.
Результаты и их обсуждение.
Было показано, что в/в и в/ж введение конъюгата β -Э c гистоном Н2а вызывает у животных кратковременное повышение АД (10–15 мин), повышение температуры тела и снижение суммарной двигательной активности (в течение 2–3 дней). Подобные изменения наблюдали также и при в/ж введении нативного β -Э.
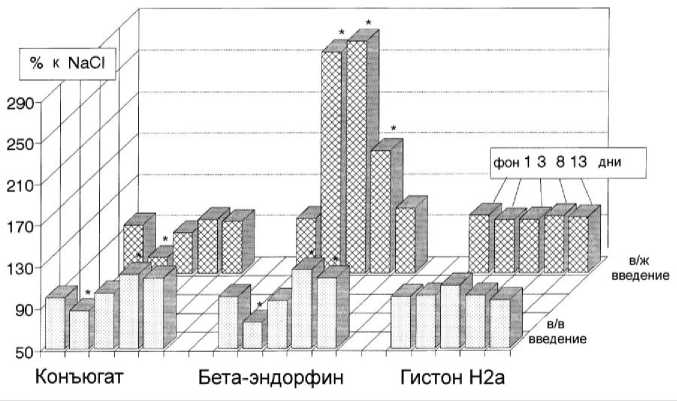
Рис. 2. Изменения болевой чувствительности в ходе эксперимента (* - p<0,05 по сравнению с фоновым периодом).
Обнаружено, что в/в и внутримозговое введение конъюгата β -Э c гистоном Н2а приводит к пролонгированным фазным изменениям пищевого поведения крыс, выражающимся в уменьшении количества потребляемой пищи в течение 1-й недели после введения и последующем его увеличении. Периферическое введение нативного β -Э оказывает аналогичное влияние на потребление пищи, в то время как в/ж введение оказывает только стимулирующее действие на прием пищи (рис. 1).
Были выявлены длительные фазные изменения болевой чувствительности в условиях в/в и внутримозгового введения конъюгата β-Э c гистоном Н2а, включавшие гипералгезию (в течение первых четырех суток после введения), сменявшуюся в дальнейшем отчетливой гипоалгезией. Такой же характер имели изменения болевой чувствительности после в/в введения нативного. После в/ж введения нативного β-Э был отмечен выраженный гипоалге-тический эффект (рис. 2).
Таким образом, эффекты как в/в, так и в/ж введений конъюгата β -Э c гистоном Н2а на пищевое поведение и болевую чувствительность однонаправленны, а в случае введения нативного β -Э – полярны.
В экспериментах с в/в введением конъюгата меченого по 125I β -Э c гистоном Н2а показано, что по сравнению с контролем (нативным 125I- β -Э) замедлено его выведение из организма. Распределение конъюгата меченого по 125I β -Э c гистоном Н2а в органах и тканях у крыс существенно отличается от распределения нативного 125I- β -Э. Следует подчеркнуть, что содержание меченого конъюгата в мозге в 3–4 раза превышает содержание нативного β -Э (рис. 3).
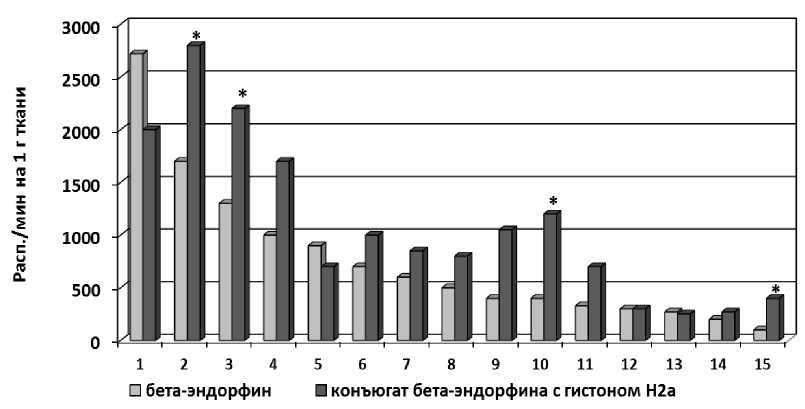
Рис. 3. Распределение конъюгата меченого по 125I β -Э c гистоном Н2а и 125I- β -Э в органах и тканях у крыс (через 3 часа после в/в введения). 1 – почки; 2 – надпочечники; 3 – мочевой пузырь; 4 – кишечник; 5 – желудок; 6 – щитовидная железа; 7 – печень; 8 – поджелудочная железа; 9 – легкие; 10 – тимус; 11 – селезенка; 12 – сердце; 13 – семенники; 14 – мышцы; 15 – мозг (*– p<0,05).
На основании сравнительного анализа эффектов периферического и внутримозгового введения конъюгата β -Э c гистоном Н2а и нативного β -Э можно полагать, что характер их взаимодействия с опиатными рецепторами различен и определяется особенностями их конформационной структуры. Полученные данные позволяют рассматривать конъюгат c гистоном Н2а как частичный конкурентный антагонист эндогенного β -Э, в отличие от нативного экзогенно введенного β -Э, действующего как специфический лиганд опиатных рецепторов. Учитывая эндогенное происхождение гистона Н2а, являющегося одним из основных внутриклеточных белков, можно предположить существование в организме эндогенных соединений, аналогом которых, вероятно, является синтезированный в настоящей работе конъюгат β -Э с гистоном Н2а. По-видимому, соединения такого рода оказывают модулирующее влияние на процессы интеграции центральной и периферической опиоидной систем организма.
Полученные результаты согласуются с нашим предположением [2] о возможной самостоятельной физиологической роли эндогенных конъюгатов регуляторных пептидов с белками, как нового класса биорегуляторов, в процессах системной организации физиологических функций, имеющих, по-видимому, преимущественно интегративное значение по сравнению с нативными пептидами.
Список литературы Эффекты введения конъюгата ß-эндорфина с гистоном Н2а на поведенческие и вегетативные показатели у крыс
- Калюжный Л.В. Физиологические механизмы регуляции болевой чувствительности. -М.: Медицина, 1984. -216 с.
- Котов А.В., Толпыго С.М., Певцова Е.И., Обухова М.Ф. Белковопептидные комплексы в механизмах врожденных и приобретенных форм поведения//Вестник РАМН. -2001. -№ 4. -С. 36-43.
- Маслов Л.Н., Лишманов Ю.Б., Терашвили М., Малкова Н.В. Эндорфиновое звено эндогенной опиоидной системы: локализация, рецепция, функция//Патол. физиол. и эксп. терапия. -2004. -№ 3. -С. 15-23.
- Молекулярная биология клетки: в 3-х томах/Б. Альбертс, А. Джонсон, Д. Льюис и др. -М.-Ижевск: НИЦ «Регулярная и хаотическая динамика», Институт компьютерных исследований, 2013. -Том I. -С. 325-359.
- Bodnar R.J. Endogenous opiates and behavior: 2012//Peptides. -2013. -Vol. 50. -P. 55-95.
- Bodnar R.J. Endogenous opiates and behavior: 2014//Peptides. -2016. -Vol. 75. -P. 18-70.
- Kumagai A.K., Eisenberg J.B., Pardridge W.M. Absorptive-mediated endocytosis of cationized albumin and a beta-endorphin-cationized albumin chimeric peptide by isolated brain capillaries. Model system of blood-brain barrier transport//J.Biol.Chem. -1987. -Vol. 262, № 31. -P. 15214-15219.
- Olson G.A., Olson R. D., Kastin A.J. Endogenous Opiates//Peptides. -1989. -Vol. 10. -P. 205-236.
- Pardridge W.M., Triguero D., Buciak J.L. Beta-endorphin chimeric peptides: transport through the blood-brain barrier in vivo and cleavage of disulfide linkage by brain//Endocrinology. -1990. -Vol. 126, № 2. -P. 977-984.
