Экспериментальное изучение особенностей формирования парагемолитическими вибрионами биопленки на поверхностях биотических объектов

Автор: Полеева Марина Владимировна, Чемисова Ольга Сергеевна, Водопьянов Сергей Олегович, Меньшикова Елена Аркадьевна, Курбатова Екатерина Михайловна
Журнал: Вестник Пермского университета. Серия: Биология @vestnik-psu-bio
Рубрика: Микробиология
Статья в выпуске: 4, 2019 года.
Бесплатный доступ
Наиболее распространенными патогенами, встречающимися на морепродуктах, способными вызвать серьезные заболевания у людей, являются парагемолитические вибрионы. Занимая различные экологические ниши, V. parahaemolyticus может существовать в воде в планктонной форме или же быть прикрепленным к инертным частицам, таким как зоопланктон, рыбы, моллюски и ракообразные, и формировать биопленку. Цель работы - изучение особенностей формирования биопленки парагемолитическими вибрионами на поверхностях биотических объектов для дальнейшего совершенствования алгоритма исследования на галофильные вибрионы в зонах рекреации на этапе отбора проб. Изучение проводили бактериологическим методом и методом Real-Time ПЦР. Показано, что парагемолитические вибрионы активно используют в качестве питательных веществ компоненты чешуи и хитина, что способствует образованию биопленки и интенсивному размножению на биотических поверхностях. Высокую концентрацию жизнеспособных клеток в биопленке на поверхности биотических субстратов следует учитывать при выборе объектов исследования на наличие V. parahaemolyticus, особенно в зонах рекреации.
Биопленка, планктон, биотические субстраты, днк, хитин, чешуя
Короткий адрес: https://sciup.org/147227107
IDR: 147227107 | УДК: 579.843:579.26:547.995.12 | DOI: 10.17072/1994-9952-2019-4-417-425
Experimental study of peculiarities of formation of vibrio parahaemolyticus biofilms on biotic surfaces of objects
The most common pathogen found on seafood and capable of causing serious diseases in humans are, V. parahaemolyticus. Occupying different ecological niches, V. parahaemolyticus due to the presence of one polar flagellum can exist in water in plankton form or be attached to inert particles such as zooplankton, fish, mollusks and crustaceans and form biofilm. The aim of the work was to study the features of the formation of biofilm by V. parahaemolyticus on the surfaces of biotic objects for further improvement of the algorithm of research on halophilic vibrions in recreation areas at the stage of sampling. The study of the features of the formation of the biofilm by vibrions was carried out by bacteriological method and Real-Time PCR. It is shown that the utilization of V. parahaemolyticus actively use as nutrients the components of scales and chitin, which contributes to the formation of biofilms and intensive reproduction on biotic surfaces. The high concentration of viable cells of V. parahaemolyticus in the biofilm on the surface of biotic substrates should be taken into account when choosing the objects of study for the presence of V. parahaemolyticus, especially in recreation areas.
Текст научной статьи Экспериментальное изучение особенностей формирования парагемолитическими вибрионами биопленки на поверхностях биотических объектов
Ежегодно в мире вылавливается и потребляется свыше 100 млн т морепродуктов [Cisneros-Montemayor et al., 2016], которые являются скоропортящимися и представляют опасность для здоровья из-за загрязнения патогенами [Reyhanath,
Kutty, 2014]. Поэтому безопасность морепродуктов важна для поддержания здоровья населения.
Наиболее распространенными патогенами, встречающимися на морепродуктах (креветки, рыба, моллюски) и способными вызвать серьезные заболевания у людей, являются парагемолитиче-ские вибрионы [Lee, Liu, Huang, 2003; Беленева,
Масленникова, Магарламов, 2004; Lin, Yu, Chou, 2004; Park et al., 2004; Тарасенко др., 2016]. Занимая различные экологические ниши, V. parahaemolyticus, благодаря наличию одного полярного жгутика, может существовать в воде в планктонной форме или же быть прикрепленным к инертным частицам, таким как взвешенные частицы, зоопланктон, рыбы, моллюски и ракообразные [Скитович, Шадрова, Прунтова, 2015].
Употребление морепродуктов, зараженных V. parahaemolyticus, приводит к вспышкам болезней пищевого происхождения (Китай, Австралия, Индонезия, Россия), протекающим по типу пищевой токсикоинфекции. Кроме того, парагемолитиче-ские вибрионы могут быть причиной заболеваний не только пищевого характера людей. Известны случаи инфицирования ран, ушей, связанные с пребыванием в морской воде; имеются сообщения о пневмонии, септицимии, хроническом посттравматическом отите, вызванном V. parahaemolyticus [Бойко, 1994; Deepanjali et al., 2005; Zhang, Orth, 2013; Shaw et al., 2014; Elmahdi, DaSilva, Parveen, 2016].
Непосредственное заражение человека от воды – явление редкое, так как парагемолитические вибрионы, в первую очередь, встречаются в морской воде, которая для питьевых целей не употребляется. В тех же количествах морской воды, которые попадают в организм человека при купании, видимо, не содержится того количества возбудителя, которое могло бы вызвать заболевание. Этим и следует объяснить единичность сообщений о заболеваниях, связанных с водными заражениями человека бактериями V. parahaemolyticus . Однако такую возможность нельзя исключать, учитывая вспышку диареи в г. Абиджан (Кот-д'Ивуар) в 1987 г., обусловленную V. parahaemolyticus и связанную с употреблением воды из лагуны, которая продолжались 8 недель и охватила 84 человека [Хотько, Дмитриев, 2002]. Были зарегистрированы случаи выделения парагемолитических вибрионов и в пресноводных водоемах [Бойко, 1994; Chakraborty, Nair, Shinoda, 1997]. Возможны случаи вторичного инфицирования овощных продуктов при обработке их водой, содержащей V. parahaemolyticus .
На протяжении большей части истории микробиологии микроорганизмы в основном характеризовались как планктонные, свободно взвешенные клетки и описывались на основе их характеристик роста в питательно богатых культуральных средах. Впервые микробиологический феномен того, что микроорганизмы прикрепляются и растут на поверхностях, описал Э. ван Левенгук, увидев микроорганизмы на поверхности зубов, еще в конце XVII в. Достижения технологии электронной микроскопии привели к обнаружению структурных поверхностно-ассоциированных организованных микроорганизмов, заключенных во внеклеточную матрицу полимерного вещества – биопленок [Donlan, 2002].
Биопленка представляет собой совокупность микробных клеток, которые необратимо связаны (не удаляются путем мягкого ополаскивания) с поверхностью и заключены в матрицу преимущественно полисахаридного материала и могут образовываться на самых разных поверхностях, таких как живые ткани, медицинские изделия, трубопроводы [Шварц, 2015]. Граница раздела фаз «твердое вещество-жидкость» обеспечивает идеальную среду для прикрепления и роста микроорганизмов. Отмечено, что степень микробной колонизации увеличивается по мере увеличения шероховатости поверхности, что связано с большей площадью прикрепления [Characklis, McFeters, Marshall, 1990]
Способность бактерий к формированию биопленок приводит к тому, что биопленочные штаммы демонстрируют множественную лекарственную устойчивость [Ahmed et al., 2018], устойчивость к дезинфектантам [Song et al., 2017], что представляет собой потенциальную угрозу для здоровья человека. Формируя биопленки, микроорганизмы могут быть в 1000 раз более устойчивыми к стрессу, чем планктонные формы [Brooun, Liu, Lewis, 2000; Ashrafudoulla et al., 2019], сохраняя при этом способность к заражению, что демонстрирует важность изучения биопленкообразования у болезнетворных микроорганизмов. Опасность заключается также в том, что наблюдается широкое распространение морепродуктов по всей стране из городов, занимающихся рыбным промыслом, что может привести к возникновению заболевания в случае инфицированности их вибрионами в городах, не являющихся прибрежными и не имеющими прямого контакта с морем [Mizan et al., 2016].
Согласно МУК 4.2.2046-06, по эпидпоказаниям на V. parahaemolyticus исследуют пищевые продукты, воду открытых водоемов, гидробионтов. Способность к формированию биопленок требует дополнительного изучения в плане определения объектов, способствующих накоплению возбудителя и требующих внимания при проведении микробиологических исследований.
Так как парагемолитические вибрионы выделяются как из воды поверхностных водоемов, так и из гидробионтов, целью нашей работы явилось изучение особенностей формирования биопленки бактериями вида V. parahaemolyticus на поверхностях биотических объектов для дальнейшего совершенствования алгоритма исследования на га-лофильные вибрионы в зонах рекреации на этапе отбора проб.
Материалы и методы исследования
Материал для исследования
В работе были исследованы штаммы V. рa-rahaemolyticus , характеристика которых приведена в табл. 1. Штаммы имели как природное, так и клиническое происхождение и различались по наличию основного фактора патогенности параге-молитических вибрионов – гена tdh – прямого термостабильного гемолизина V. parahaemolyticus . Все штаммы были получены из коллекции Музея живых культур с Центром патогенных для человека вибрионов ФКУЗ Ростовский противочумный институт Роспотребнадзора.
Таблица 1
Характеристика штаммов, использованных в работе
|
№ штамма |
Генетическая характеристика |
Объект выделения |
Место и год выделения |
|
13331 |
tdh+ trh- |
человек |
Япония, 1971 |
|
14704 |
tdh- trh- |
рыба |
Азовское море, 1984 |
|
14810 |
tdh+ trh- |
человек |
г. Новороссийск, 1976 |
|
16696 |
tdh- trh- |
морская вода |
г. Новороссийск, 1973 |
|
19013 |
tdh+ trh- |
человек |
г. Бердянск, 1988 |
|
19149 |
tdh- trh- |
морская вода |
г. Находка, 2009 |
Методы исследования
Питательные среды . Для культивирования штаммов использовали 1%-ную пептонную воду с 2%-ным NaCl (рН 7.8); 2%-ный агар Мартена с 2%-ным NaCl (рН 7.7); 0.3%-ный агар Мартена с 2%-ным NaCl (рН 7.7).
Биотические субстраты . В качестве субстратов использовали чешую рыбы и хитиновый экзоскелет креветки, которые нарезали пластинами 5×5 мм, помещали в количестве 30 штук во флаконы с 50 мл морской воды и автоклавировали при 130ºС, 1.5 атм в течение 60 мин.
Выращивание биопленки. Из суточной агаровой культуры штаммов V. parahaemolyticus готовили взвесь 109 микробных клеток/мл (мк.кл./мл). Во флаконы с 50 мл морской воды и субстратами добавляли взвесь вибрионов до конечной концентрации во флаконе 104 мк.кл./мл. Параллельно проводили высев на твердую питательную среду для определения КОЕ/мл. В качестве контроля все исследуемые штаммы засевали во флаконы с 50 мл морской воды без субстратов. Флаконы инкубировали в течение 1 месяца при комнатной температуре (t = 25±2ºС), что соответ- ствует температурным условиям окружающей среды в летнее время года.
Бактериологические методы . Для контроля жизнеспособности штаммов V. parahaemolyticus использовали метод бактериологического посева супернатантов и отпечатков субстрата на твердую питательную среду – агар Мартена с 2%-ным NaCl [Титова, Веркина, 2016]. Результат оценивали по наличию роста.
Микроскопия . Образцы пластинок субстратов (чешуи и хитина) с сформировавшейся биопленкой окрашивали 0.1%-ным водным раствором Конго красным по методике, предложенной С.В. Титовой и Л.М. Веркиной [2016]. Микроскопию проводили на микроскопе Nicon Eclipse E200 (Japan).
Real-Time ПЦР . Для проведения ПЦР в «реальном времени» отбирали по 0.3 мл из контрольных проб. Фрагменты хитина и чешуи стерильно трижды отмывали физиологическим раствором (рН 7.2) от несвязавшихся клеток, избавлялись от излишков влаги с помощью фильтровальной бумаги и помещали в 0.3 мл дистиллированной воды. Также отбирали по 0.3 мл проб планктонных микроорганизмов. Все пробы обеззараживали прогреванием при 100ºС в течение 30 мин., согласно руководству [Организация …, 2010].
Для проведения Real-Time ПЦР с количественным учетом результатов использовали сконструированные нами праймеры и зонд к гену металло-протеазы/коллагеназы V. parahaemolyticus [Рыков-ская и др., 2018]. Постановку реакции амплификации проводили в автоматическом детектирующем амплификаторе ДТ-Lite (ДНК-технология, Россия). Учет детекции флуоресцентного свечения проводили по каналу ROX с длиной волны 605 нм.
Результаты и их обсуждение
Для проведения Real-Time ПЦР в протокол исследования проб были включены образцы с известным количеством искомой ДНК – калибровочные стандарты, в качестве которых использовали взвеси суточных культур парагемолитических вибрионов с концентрацией клеток 109–102 м.кл./мл. Количество клеток взвесей определяли путем высева на чашки Петри и подсчета КОЕ. Программа проводит построение кривой линии по точкам калибровочных стандартов, в соответствии с которой проводится расчет концентрации исследуемых проб.
Отбор проб для постановки Real-Time ПЦР и бактериологического высева на питательные среды проводили ежедневно в течение 7 дней, затем один раз в неделю в течение 1 месяца.
В течение первых 3 сут. культивирования отме- чался активный рост парагемолитических вибрионов во всех испытуемых пробах, но степень роста была разной. Так, наименьшей скоростью роста характеризовались штаммы в контрольных флаконах с морской водой. Штаммы, культивируемые в морской воде с биотическими субстратами, имели более высокую скорость роста, что подтверждалось и бактериологическим методом путем подсчета КОЕ/мл, и методом Real-Time ПЦР при определении количества копий ДНК в течение всего периода наблюдения. Слабый рост в контрольных пробах с морской водой, по сравнению с планктоном над хитиновыми пластинками и чешуей, говорит о недостатке питательных веществ и использовании вибрионами биотических субстратов в качестве источника питания.
Через 72 ч. культивирования было отмечено, что штаммы парагемолитических вибрионов, не имеющие гена tdh , образуют на поверхности морской воды с хитином и чешуей довольно плотную пленку, в отличие от токсигенных штаммов, у одного из которых пленка визуально отсутствовала ( V. parahaemolyticus 19013), а у двух других была значительно менее выражена, чем у штаммов, выделенных из объектов окружающей среды.
К 14-м сут. наблюдения пленка на поверхности жидкости во флаконе с пластинками хитина и чешуей увеличилась и стала более плотной. В контрольных пробах при визуальном учете на поверхности раздела фаз «вода-воздух» все штаммы независимо от токсигенности и происхождения биопленки не образовывали на протяжении всего периода наблюдения.
Изучение особенностей роста и формирования биопленок на поверхности субстратов показало, что парагемолитические вибрионы по-разному проявляют себя на хитине и на чешуе рыб. Так, на 6-е сутки культивирования, при отборе пластинок субстратов было отмечено, что клинические токси-генные штаммы в процессе жизнедеятельности утилизируют пластинки чешуи, в результате чего они становятся рыхлыми и при прикосновении к ним распадаются. Согласно литературным данным, чешуя рыб содержит 39–68% азотистых веществ (щелочерастворимых, водорастворимых и солерастворимых белков), 27–49% минеральных веществ (кальций, фосфор, магний и др.), 0.1–0.2% жира. Основная доля белков чешуи приходится на коллаген – 48–59%. Вибрионы способны активно вырабатывать фермент коллагеназу, что объясняет использование чешуи в качестве источника питательных веществ для роста и размножения микробных клеток [Якубова, Котенко, 2004; Антипова, Ву Тхи Лоан, 2009; Као, Нгуен, Карапун, 2016]. Динамика формирования биопленок штаммами поверхностных водоемов представлена на рис. 1.
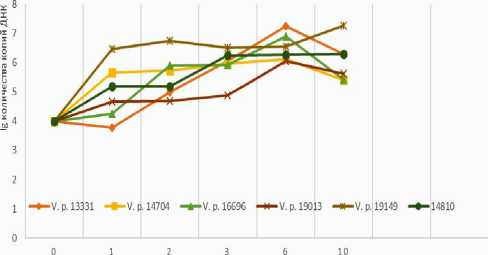
Время культивирования, сутки
Рис. 1 . Динамика изменения количества копий ДНК при выращивании на пластинках чешуи
Столь активное использование рыбьей чешуи в качестве источника питания для роста и размножения парагемолитических вибрионов приводит к выводу о ее возможном применении в качестве основы для приготовления питательных сред. Учитывая, что чешуя является отходом рыбного производства, который образуется в большом количестве, возможное использование ее для приготовления питательных сред привело бы к снижению затрат на их производство.
Данные количественного ПЦР-анализа свидетельствуют о том, что образование биопленки на хитиновом панцире креветок шло активнее, чем на чешуе рыб (рис. 2). Панцирь креветки, по разным данным, состоит из белка – 7–37%, липидов – 3– 5%, углеводов – до 13%, минеральных веществ – до 22% [Осовская и др., 2010; Муравьев, 2017; Аллам, 2018]. Количество хитина составляет от 5– 25%. Хитин панциря креветок Penaeus semisulcatus может быть использован в качестве источника хитина и хитозана [Аллам, 2018].
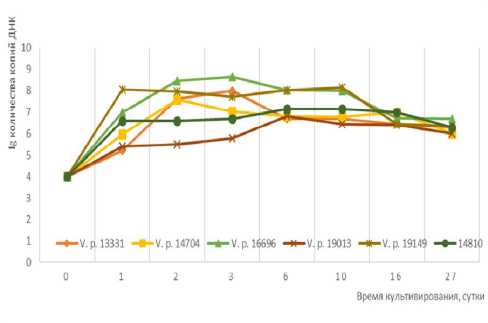
Рис. 2. Динамика изменения количества копий ДНК при выращивании на пластинках хитина
Поскольку хитин имеет компактную структуру, он нерастворим в большинстве растворителей, его биодоступность для микроорганизмов предположительно является невысокой, что объясняет меньшие темпы роста бактерий V. parahaemo-lyticus в планктоне с хитином. Однако наличие большого количества других питательных веществ позволяет микроорганизмам активнее размножаться в сравнении с контрольными флаконами с морской водой, где отсутствуют какие-либо питательные субстраты. Это согласуется с количественными результатами Real-Time ПЦР, согласно которым нарастание количества копий ДНК наиболее активно идет именно в планктоне над чешуей и хитином.
При культивировании контрольных проб с морской водой в течение 72 ч. методом Real-Time ПЦР было показано, что во всех пробах количество копий ДНК увеличивалось (рис. 3). На 6-e сутки инкубирования количество копий ДНК парагемоли-тических вибрионов значительно снизилось, что, учитывая отсутствие каких-либо питательных веществ в морской воде, может свидетельствовать о том, что ДНКаза вибрионов расщепляет ДНК клеток, и продукты расщепления используются для выживания других клеток [Беленева, Масленникова, Магарламов, 2004]. Такая же закономерность прослеживается и при изучении планктонных форм микроорганизмов из проб, содержащих пластинки чешуи и хитина.
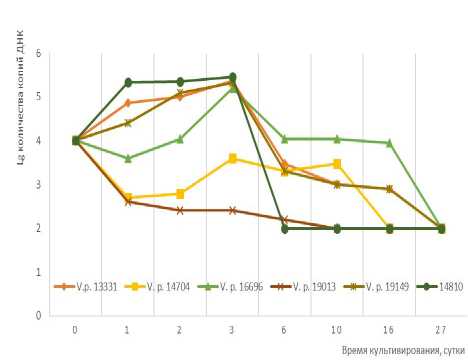
Рис. 3. Динамика изменения количества копий ДНК при выращивании в морской воде
Через 27 сут. культивирования ПЦР анализ показал, что количество копий ДНК в контрольной и планктонных пробах стало менее 103 в мл. При этом в биопленочных пробах количество копий ДНК V. parahaemolyticus было 104–106.
Таблица 2
Отношение количества копий ДНК планктонной формы к биопленочной
|
№ штамма |
Субстрат |
24 ч. |
48 ч. |
72 ч. |
16 сут. |
|
13331 |
чешуя |
2583 |
- |
- |
- |
|
хитин |
1108 |
13.6 |
0.2 |
1.1 |
|
|
14704 |
чешуя |
143.6 |
58.5 |
36.5 |
- |
|
хитин |
150.2 |
0.03 |
0.7 |
0.3 |
|
|
14810 |
чешуя |
59.6 |
156.1 |
10 |
- |
|
хитин |
40.8 |
40.8 |
0.02 |
- |
|
|
16696 |
чешуя |
15056 |
5841 |
457.4 |
3,7 |
|
хитин |
15.7 |
- |
- |
- |
|
|
19013 |
чешуя |
1891 |
4615 |
802.6 |
- |
|
хитин |
1360 |
8943 |
476.3 |
0.2 |
|
|
19149 |
чешуя |
169.7 |
450.6 |
51.9 |
- |
|
хитин |
3.2 |
5.6 |
4.8 |
0.2 |
Было установлено, что через 24 ч. инкубирования соотношение планктонной формы к биопле-ночной максимально независимо от наличия генов токсигенности и происхождения штаммов, т. е. в этот период времени количество копий ДНК в планктоне выше, чем в биопленке (табл. 2), и формирование биопленки находится на начальном этапе. Через 48 ч. у трех штаммов (14810, 19013 и 10149) соотношение биопленочной и планктонной форм продолжало расти, а у трех штаммов снизилось. К 16 дню инкубирования у всех штаммов данное соотношение значительно снизилось (у штамма V. parahaemolyticus 16696 в 4000 раз), что свидетельствует о том, что большая часть планктонных штаммов перешла в биопленочную форму для переживания неблагоприятных условий (снижение питательных веществ и отравление продуктами собственной жизнедеятельности).
Выводы
-
1. Утилизация парагемолитическими вибрионами в качестве питательных веществ компонентов чешуи и хитина способствует образованию биопленки и интенсивному размножению на биотических поверхностях.
-
2. Высокую концентрацию жизнеспособных клеток парагемолитических вибрионов в биопленке на поверхности биотических субстратов следует учитывать при выборе объектов исследования на наличие V. parahaemolyticus , особенно в зонах рекреации.
Список литературы Экспериментальное изучение особенностей формирования парагемолитическими вибрионами биопленки на поверхностях биотических объектов
- Аллам А.Ю. Разработка технологии получения хитозана из панциря креветки Penaeus semisulcalcatus и изучение возможности его использования в качестве пищевых покрытий и добавок: автореф. дис. … канд. техн. наук. Астрахань, 2018. 20 с.
- Антипова Л.В., Ву Тхи Лоан. Чешуя прудовых рыб - источник пищевого продукта [Электронный ресурс]. URL: http://www.tstu.ru/education/elib/pdf/st/2009/antipova
- Беленева И.А., Масленникова Э.Ф., Магарламов Т.Ю. Физиолого-биохимические свойства галофильных вибрионов Vibrio parahaemolyticus и V. alginolyticus, изолированных из гидробионтов залива Петра Великого Японского моря // Биология моря. 2004. Т. 30, № 2. C. 114-119.
- Бойко А.В. Факторы персистенции микроорганизмов // Эпидемиология, клиника, диагностика, лечение и профилактика важнейших инфекционных болезней: материалы конф. Тамбов; Астрахань, 1994. С. 33.
- Бойко А.В., Погорелова Н.П., Жигарева Т.М. Эколого-эпидемиологические аспекты распространенности Vibrio parahaemolyticus в пресноводном регионе // Журнал микробиологии, эпидемиологии и иммунологии. 1994. № 3. С. 49-51.
- Као Т.Х., Нгуен Т.М., Карапун М.Ю. Чешуя рыб как источник получения пищевого желатина // Молодой ученый. 2016. № 23. С. 113-115.
- Методы выявления и определения парагемолитических вибрионов в рыбе, нерыбных объектах промысла, продуктах, вырабатываемых из них, воде поверхностных водоемов и других объектах: МУК 4.2.2046-06. М.: Федеральный центр госсанэпиднадзора Минздрава России, 2006. 26 с.
- Муравьев А.А. Растворы смесей целлюлозы и хитина в ионных жидкостях и композиционные материалы на их основе: автореф. дис. … канд. хим. наук. СПб., 2017. 24 с.
- Организация работы лабораторий, использующих методы амплификации нуклеиновых кислот при работе с материалом, содержащим микроорганизмы I-IV групп патогенности: МУ 1.3.2569-09. М.: Федеральный центр гигиены и эпидемиологии Роспотребнадзора, 2010. 51 c.
- Осовская И.И. и др. Хитин-глюкановые комплексы. Физико-химические свойства и молекулярные характеристики: учеб. пособие / под. ред. Г.М. Полторацкого. СПб., 2010. 52 с.
- Рыковская О.А. и др. Разработка способа идентификации V. parahaemolyticus с помощью ПЦР в режиме реального времени // Здоровье населения и среда обитания. 2018. № 3 (300). С. 48-50.
- Скитович Г.С., Шадрова Н.Б., Прунтова О.В. Vibrio parahaemolyticus: распространение, выявление и методы идентификации // Ветеринария сегодня. 2015. № 3 (14). С. 66-70.
- Тарасенко Т.Т. и др. Заболеваемость прочими кишечными инфекциями в Приморском крае // Здоровье. Медицинская экология. Наука. 2016. Т. 3, № 66. С. 127-134.
- Титова С.В., Веркина Л.М. Моделирование биопленок холерного вибриона на твердых поверхностях (стекло и пластик) и визуализация их в световом и люминисцентном микроскопах // Клиническая и лабораторная диагностика. 2016. Т. 61, № 4. С. 238-241.
- Хотько Н.И., Дмитриев А.П. Водный фактор в передаче инфекций. Пенза, 2002. 232 с.
- Шварц Т.А. Биопленки как микробное сообщество // Вестник Курганского государственного университета. 2015. № 1, вып.7. С. 41-44.
- Якубова О.С., Котенко А.Л. Чешуя как источник получения ихтиожелатина // Вестник Астраханского государственного технического университета. 2004. № 2 (21). С. 130-134.
- Ahmed H.A. et al. Molecular characterization, antibiotic resistance pattern and biofilm formation of Vibrio parahaemolyticus and V. cholerae isolated from crustaceans and humans // International Journal of Food Microbiology. 2018. Vol. 274. P. 31-37.
- Ashrafudoulla Md. et al. Genetic Relationship, Virulence Factors, Drug Resistance Profile and Biofilm Formation Ability of Vibrio parahaemolyticus Isolated From Mussel // Front. Microbiol. 2019. Vol. 10. 513.
- DOI: 10.3389/fmicb.2019.00513
- Brooun A., Liu S., Lewis K. A dose-response study of antibiotic resistance in Pseudomonas aeruginosa biofilms // Antimicrobial Agents and Chemotherapy. Mar. 2000. P. 640-646.
- Chakraborty S., Nair G.B., Shinoda S. Pathogenic vibrios in the natural aquatic environment // Rev. Environ. Health. 1997. Vol. 12 (2). P. 63-80.
- Characklis W.G., McFeters G.A., Marshall K.C. Physiological ecology in biofilm systems // Biofilms / Characklis W.G., Marshall K.C., eds. New York: John Wiley & Sons, 1990. P. 341-394.
- Cisneros-Montemayor A.M. et al. A global extimate of seafood consumption by coastal indigenous peoples // PLoS One. 2016. 11. e0166681. 10.1371/journal.pone. 0166681.
- DOI: 10.1371/journal.pone.0166681
- Deepanjali A. et al. Seasonal variation in abundance of total and pathogenic Vibrio parahaemolyticus bacteria in oysters along the southwest coast of India /// Appl. Environ. Microbiol. 2005. Vol. 71, № 7. P. 3575-3580.
- Donlan M.R. Biofilms: Microbial Life on Surface // Emerg. Infect. Dis. 2002. Vol. 8, № 9. P. 881-890.
- Elmahdi S., DaSilva L.V., Parveen S. Antibiotic resistance of Vibrio parahaemolyticus and Vibrio vulnificus in various countries: A review // Food Microbiol. 2016. Vol. 57. P. 128-134.
- Lee K.K., Liu P.C., Huang C.Y. Vibrio parahaemolyticus infectious for both humans and edible mollusk abalone // Microbes Infect. 2003. Vol. 5. P. 481-485.
- Lin C., Yu R.C., Chou C.C. Susceptibility of Vibrio parahaemolyticus to various environmental stresses after cold shock treatment // Int. J. Food Microbiol. 2004. Vol. 92. P. 207-215.
- Mizan M.F.R. et al. Effects of NaCl, glucose, and their combinations on biofilm formation on black tiger shrimp (Penaeus monodon) surfaces by Vibrio parahaemolyticus // Food Control. 2016. Vol. 89. P. 203-209.
- Park K.S. et al. Functional characterization of two type III secretion systems of Vibrio parahaemolyticus // Infect. Immun. 2004. Vol. 72. P. 6659-6665.
- Reyhanath P.V., Kutty R. Incidence of multidrug resistant Vibrio parahaemolyticus isolated from Ponnani, South India // Iran. J. Microbiol. 2014. Vol. 6, № 2. P. 60-67.
- Shaw K.S. et al. Antimicrobial susceptibility of V. vulnificus and V. parahaemolyticus recovered from recreational and commercial areas of Chesapeake Bay and Mareland coastal bays // PLoS One. 2014. Vol. 9 (2).
- DOI: 10.1371/journal.pone.0089616
- Song X. et al. Effect of temperature on pathogenic and non-pathogenic Vibrio parahaemolyticus biofilm-formation // Food Control. 2017. Vol. 7. P. 485-491.
- Zhang L., Orth K. Virulence determinants for Vibrio parahaemolyticus infection // Curr. Opin. Microbiol. 2013. Vol. 16, № 1. P. 70-77.
