Экзогенная регуляция фенотипического проявления признака фотопериодической чувствительности у сои

Автор: Лукомец В.М., Зеленцов С.В., Мошненко Е.В.
Рубрика: Селекция, семеноводство и биотехнология сельскохозяйственных растений
Статья в выпуске: 4 (192), 2022 года.
Бесплатный доступ
Соя является фотопериодически высокочувствительным короткодневным растением. В генофонде сои практически отсутствуют гены и генные комплексы фотопериодической нейтральности. Это очень ограничивает оптимальные длины дня и широтные диапазоны возделывания подавляющего большинства сортов сои. Поэтому задача нейтрализации негативного влияния неоптимальных, прежде всего укороченных, длин дня на сою имеет глобальную значимость. Целью настоящих исследований было изучение возможности экзогенной нейтрализации фотопериодической чувствительности сои путём применения растворов гиббереллина по вегетирующим растениям с целью пролонгации периода их вегетативного роста, формирования дополнительных узлов бобов и семян, обеспечивающих увеличение урожайности на фоне неоптимальных, укороченных длин дня. Исследования проводили в 20192022 гг. в ФГБНУ ФНЦ ВНИИМК (г. Краснодар) на сортах сои Пума, Баргузин, Вита и Славия. На основе теории гормональной регуляции цветения М.Х. Чайлахяна разработан способ экзогенной регуляции генетически детерминированного признака высокой фотопериодической чувствительности сои к укороченным длинам дня. Установлена наиболее чувствительная к экзогенной регуляции фотопериодической чувствительности стадия онтогенеза сои - 4-й тройчатосложный лист (стадия онтогенеза V4), посредством обработки растений раствором гиббереллина. Определена наиболее активная концентрация растворов гиббереллина - 0,003 %, нанесение которой на листья сои на стадии онтогенеза V4 вызывает пролонгацию вегетативного роста растений, обеспечивает формирование дополнительных узлов, бобов и семян. Экзогенная фитогормональная пролонгация вегетативного роста увеличивает урожайность фотопериодически чувствительных сортов сои при их выращивании в условиях укороченных длин дня, в том числе, в повторных летних посевах после уборки озимых колосовых культур.
Соя, летние сроки посева, укороченный фотопериод, реакция на длину дня, фотопериодическая чувствительность, гиббереллин, экзогенная регуляция признака
Короткий адрес: https://sciup.org/142235866
IDR: 142235866 | УДК: 633.853.52:581.1.043 | DOI: 10.25230/2412-608X-2022-4-192-35-52
The exogenous regulation of a phenotypic manifestation of a trait of a photoperiodic sensitivity in soybean
Soybean is a photoperiodically highly sensitive, short-day plant. There are practically no genes and gene complexes of photoperiodic neutrality in the soybean gene pool. This greatly limits the optimal day lengths and latitudinal ranges of cultivation of the vast majority of soybean varieties. Therefore, the task of neutralizing the negative impact of suboptimal, first of all, shortened day lengths on soybeans is of global importance. The purpose of these studies was to study the possibility of exogenous neutralization of photoperiodic sensitivity of soybeans by using gibberellin solutions on vegetative plants in order to prolong the period of their vegetative growth, the formation of additional nodes of beans and seeds, providing an increase in yield against the background of suboptimal, shortened day lengths. The research was carried out in 2019-2022 at the V.S. Pustovoit All-Russian Research Institute of Oil Crops (VNIIMK), Krasnodar, on soybean varieties Puma, Barguzin, Vita and Slavia. Based on the theory of hormonal regulation of flowering by M.H. Chil-akhyan we developed a method for exogenous regulation of a genetically determined trait of high photoperiodic sensitivity of soybeans to shortened day lengths. The most sensitive to exogenous regulation of photoperiodic sensitivity stage of soybean ontogeny - the 4th trifoliate leaf (ontogeny stage V4) was established by treating plants with a solution of gibberellin. The most active concentration of gibber-ellin aqueous solutions was determined - 0.003%, the application of which to soybean leaves, at the stage of ontogenesis V4, causes prolongation of vegetative plant growth, formation of additional nodes, beans and seeds. Exogenous phytohormonal prolongation of vegetative growth increases the yield of photoperiodically sensitive soybean varieties when they are grown under conditions of shortened day lengths, including in second summer crops after harvesting winter crops.
Текст научной статьи Экзогенная регуляция фенотипического проявления признака фотопериодической чувствительности у сои
Введение. Культурная соя является фотопериодически высокочувствительным, короткодневным видом цветковых растений, эволюционно сформировавшимся на северо-востоке Китая в диапазоне длин дня 13–16 ч. Отклонения фотопериодов от биологического оптимума сои оказывают заметное влияние на изменение вегетационного периода, высоты растений и их продуктивности [1; 2; 3; 4; 5; 6].
Признак высокой реакции сои на длину дня определяется эпистатическим взаимодействием всех доминантных и рецессивных аллелей генов фотопериодической чувствительности [7]. Максимальная чувствительность растений сои к продолжительности фотопериода проявляется от появления всходов до окончания массового цветения [8].
Реакция сои на увеличение длины дня выше оптимума проявляется в запаздывании наступления сроков цветения, увеличении размеров элементов цветка, в удлинении длин междоузлий и периода вегетации. При сокращении длины дня ниже критических значений фазы начала цветения и созревания сои наступают раньше. При этом на растениях уменьшаются размеры листовых пластинок, элементов цветка, длин междоузлий, особенно в нижнем и верхнем ярусах, общая высота главных побегов, количество бобов и урожай семян [2; 9; 10; 11; 12; 13]. В связи с этим успешное возделывание подавляющего большинства мирового сортимента современных сортов сои ограничено довольно узким диапазоном оптимальных для роста и развития длин дня. В большинстве случаев широтная зона максимальной адаптивности отдельного генотипа сои ограничена достаточно узкой полосой, по разным оценкам составляющей от ± 100–150 км [14] до ± 100–150 миль (≈160–240 км) [15] на север и на юг от пункта выведения сорта.
Поэтому интродукция выведенных на умеренных широтах и соответствующих длинах дня сортов сои в Северном полушарии Земли в более южные короткодневные регионы, а в Южном полушарии – в более северные влечёт за собой существенное сокращение вегетационного периода, высоты растений и снижение семенной урожайности [16].
Тем не менее широкое внутривидовое разнообразие культурной сои, в том числе и по признаку адаптивности к разной продолжительности дня, а также активная, более чем 100-летняя адаптивная селекция сои позволили распространить возделывание этой культуры на все материки, кроме Антарктиды. Современные сорта культурной сои возделываются от 58–60° с. ш. (Канада, Россия, Швеция) и до 48–53° ю. ш. (Аргентина), образовав почти непрерывный ареал распространения этой культуры по пяти континентам обоих полушарий планеты [9; 17; 18]. Очень скороспелые сорта сои северного экотипа возделываются в высокоширотных условиях Амурской, Иркутской, Омской областей и Хабаровского края России, в странах Прибалтики, в северо-восточном Китае, в Польше, Германии, Швеции, Канаде, на юге Аргентины и в Новой Зеландии.
Соя также очень активно и широко возделывается в экваториальных и субэкваториальных короткодневных регионах Африки, Центральной Америки, ЮгоВосточной Азии. Причём в тропических и субтропических странах, из-за несущественной разницы по динамике темпера- тур и длин дня в течение года, эта культура может высеваться в качестве второй или третьей культуры после риса, пшеницы, ячменя, кукурузы [9; 17; 18; 19]. По данным ФАО, на африканском континенте общая площадь посевов сои в 2020 г. составляла 2,55 млн га, в Центральной Америке - 0,19 млн га, в Юго-Восточной Азии -1,01 млн га. При этом средняя урожайность сои в странах Африки, по данным за 2020 г., составляла 1,35 т/га, в странах Центральной Америки - 1,66 т/га, в странах Юго-Восточной Азии - 1,47 т/га [20].
Безусловно, в ряде случаев урожайность в некоторых тропических странах лимитируется дефицитом воды в почве. Особенно этот стрессорный фактор выражен в примыкающих к пустыне Сахара африканских странах [19]. Но даже в условиях орошения урожайность сои в экваториальных и субэкваториальных странах на фоне очень коротких (не более 12–13 ч) длин дня заметно уступает этому показателю в основных соепроизводящих странах, расположенных на более высоких широтах субтропического и умеренного климатических поясов. Так, средняя урожайность сои в Аргентине в 2020 г. с общей площади 16,72 млн га составила 2,92 т/га, в Канаде на площади 2,04 млн га -3,12 т/га, в США с общей площади 33,31 млн га - 3,38 т/га [20].
Поэтому экономически невыгодная высокая реакция подавляющего большинства мирового сортимента сортов даже на незначительные отклонения от оптимальных для конкретного генотипа длин дня остаётся одной из важнейших проблем мирового соеводства.
При этом успехи мировой селекции сои на снижение фотопериодической чувствительности, прежде всего, к условиям укороченных экваториальных и субэкваториальных фотопериодов, а также к повторным летним посевам в умеренных широтах на фоне укорачивающихся длин дня оказались мало результативными. Основная сдерживающая причина - почти полное отсутствие в мировом генофонде культурной сои источников и доноров признака полной или хотя бы частичной фотонейтральности. В связи с этим в мировой литературе практически отсутствуют упоминания о результативной селекции сои на этот признак. В открытой печати не выявлены сведения о биотехнологических исследованиях в этом направлении.
Дополнением к периодически активизирующимся поискам селекционно-генетических способов нейтрализации высокой фотопериодической реакции сои на неоптимальные, прежде всего на укороченные длины дня, может быть разработка способов негенетического, экзогенного регулирования фенотипического проявления этого признака.
Впервые возможность физиологической регуляции фотопериодической реакции растений на длину дня была выявлена в 1936 г. советским физиологом, академиком Михаилом Христофоровичем Чайлахяном [21; 22]. В последующие годы М.Х. Чайлахян исследовал и обосновал физиологические механизмы процессов зацветания высших растений различных фотопериодических групп (короткодневных, длиннодневных, нейтральных и т.д.) и установил связь фитогормонов с индукцией цветения растений, и в итоге разработал гормональную теорию цветения высших растений [23; 24; 25; 26]. Им было установлено, что для перехода растений из вегетативной в репродуктивную стадию в их тканях должен сформироваться и накопиться комплекс фитогормонов, названный М.Х. Чайлахяном флоригенным комплексом или флоригеном. Внешними регулирующими факторами времени накопления флоригена являются, прежде всего, длина дня и температура окружающей среды, а также ряд других ресурсов среды [21; 22; 25; 26]. Важнейшим компонентом флориген-ного комплекса в листьях растений М.Х. Чайлахян называл гиббереллины (ГАз), которые не только регулируют инициацию цветения, но и приводят к повыше- нию стрессоустойчивости и увеличению высоты побегов [24; 27].
Выявленные М.Х. Чайлахяном особенности гиббереллинов побудили ряд исследователей использовать эти фитогормоны на разных географических широтах в качестве экзогенного регулятора роста и развития растений сои. Однако полученные результаты оказались неоднозначными.
Так, в ходе проведённых в штате Миннесота, США, экспериментов на широте 45° и при максимальной длине дня 15 ч 37 мин было установлено, что в результате применения растворов гиббереллина (ГА 3 ) на сое в фазы бутонизации и бобо-образования на 30 % увеличилась длина верхних междоузлий. Однако семенная продуктивность обработанных ГА 3 растений сои у разных сортов снизилась в 1,5– 3,0 раза [28].
По данным экспериментов в провинции Квебек, Канада, предпосевная обработка семян сои растворами ГА 3 на широте 47° и при максимальной длине дня 15 ч 50 мин приводила к ускорению прорастания в условиях пониженных температур почвы, что подтверждало выводы М.Х. Чайлахяна о повышении стрессо-устойчивости при увеличении содержания ГА 3 в тканях растений. Но на высоту растений, фотопериодическую реакцию и урожайность сои применение ГА 3 в условиях Квебека не оказало существенного влияния [29].
В исследованиях на южной границе субтропического пояса во Флориде (США) на широтах 27–29° и при максимальных длинах дня 13 ч 50 мин – 14 ч 00 мин применение ГА 3 в фазе всходов сои приводило к дополнительному растяжению нижних междоузлий, однако междоузлия выше пятого оказались укороченными относительно контрольных растений. Количество междоузлий на обработанных растениях не изменялось. В целом применение гиббереллина практически не повлияло на высоту растений сои и на их продуктивность. Изменения реакции на фотопериод также не было отмечено [30].
В отдельных работах был выявлен эффект частичной нейтрализации укороченного фотопериода, фенотипически проявившийся в увеличении высоты растений и их продуктивности.
Так, в субтропических условиях провинции Буэнос-Айрес (Аргентина) на широте 35° и при максимальной длине дня 14 ч 30 мин было установлено, что внесение растворов ГА 3 в фазе цветения сои приводило к снижению фотопериодической реакции на укороченный субтропический день, что фенотипически выражалось в увеличении высоты растений сои, массы и площади листовой поверхности и в удлинении вегетационного периода. Однако у экспериментальных растений оказалась частично заблокирована способность формирования азотфиксирую-щих клубеньков на корнях, что повлекло за собой снижение урожайности по сравнению с необработанным ГА 3 контрольными растениями [31; 32].
При осенних (октябрь) посевах сои на субтропическом юге Ирана на широте 29° (пров. Бушер, северное побережье Персидского залива), и зимней вегетации при длинах дня 10 ч 20 мин – 11 ч 00 мин в результате обработки растений растворами ГА 3 в фазе цветения была зафиксирована частичная нейтрализация фотопериодической реакции сои на очень короткий день. Обработанные ГА 3 растения сои сформировали почти удвоенную высоту, отличались увеличенным количеством продуктивных узлов, бобов и семян. Наиболее значимым результатом данного эксперимента стало увеличение в 1,5–3,0 раза количества сформировавшихся на растениях сои семян [33].
Таким образом, положительный эффект от применения растворов гиббереллина для регуляции фотопериодической чувствительности сои уже был выявлен на некоторых стадиях роста и развития растений. В случае подтверждения результативности такого способа нейтрализации негативного эффекта укороченных фотопериодов на сое, его можно было бы использовать для повышения продуктивности сортов сои при их возделывании в короткодневных субтропических и тропических климатических поясах или в повторных летних посевах в южной части умеренного климатического пояса на фоне сокращающихся позднелетних длин дня.
Настоящие исследования были посвящены изучению подтверждения возможности экзогенной гормональной нейтрализации признака фотопериодической чувствительности сои путём применения растворов гиббереллина по вегетирующим растениям с целью пролонгации периода их вегетативного роста и формирования дополнительных репродуктивных органов.
Материалы и методы . Исследования проводили в 2019–2022 гг. на базе ФГБНУ ФНЦ ВНИИМК, г. Краснодар, на широте 45°04′. Максимальная долгота дня на этой широте в период летнего солнцестояния – 15 ч 37 мин. Долгота дня в период посева в 3-й декаде июня – 1-й декаде июля составляла 15 ч 37 мин – 15 ч 32 мин. В период созревания в 3-й декаде сентября – 1-й декаде октября долгота дня сокращалась до 12 ч 00 мин – 11 ч 40 мин. Текущую долготу дня на широте Краснодара от всходов до созревания сои определяли с помощью таблицы времени восхода и захода солнца для данного географического пункта на сайте «Время восхода и захода солнца» [34]. Не смотря на то, что астрономически промежуток времени между восходом и заходом солнца называется долготой дня, в специализированной литературе, посвящённой различным аспектам фотопериодизма растений, наиболее часто употребляется термин «длина дня», равно как и его производное «длиннодневный». Поэтому в настоящей статье в качестве общепринятого синонима долготы дня нами также будет использоваться термин «длина дня».
Материалом для исследований служили четыре очень ранних и ранних сорта сои селекции ФГБНУ ФНЦ ВНИИМК:
Баргузин, Пума, Вита и Славия. Из них сорта Пума и Вита отличаются высокой фотопериодической чувствительностью, сорт Баргузин – средней, сорт Славия – слабой фотопериодической чувствительностью. Посев изучаемых сортов проводили селекционной сеялкой СКС-6А в 3-й декаде июня – 1-й декаде июля. В 2019 и 2021 гг. экспериментальные посевы размещали на парующем участке, в 2020 и 2022 гг. – на участке после уборки озимого ячменя. Площадь делянок 100–117 м2, повторность 3-кратная. Норма высева семян 50 шт./м2 (500 тыс. шт./га).
Для изучения эффективности экзогенной регуляции признака фотопериодической чувствительности использовали водные растворы гиббереллина (ГА 3 ) в концентрациях 0,0003, 0,003 и 0,03 %. В каждом варианте (сорт, срок посева) тракторным навесным опрыскивателем наносили на листовую поверхность растений растворы гиббереллина из расчёта 10 мл/м2. Контролем служило опрыскивание водопроводной водой.
Растворы ГА 3 и воду наносили на растения сои в следующие фазы онтогенеза: 2-й тройчатосложный лист (вегетативная стадия онтогенеза V 2 ); 4-й тройчатосложный лист (вегетативная стадия онтогенеза V 4 ); полное цветение 100 % растений (генеративная стадия онтогенеза R 2 ).
Перед внесением растворов гиббереллина, с целью компенсации попадающих на поверхность почвы в междурядьях и безвозвратно утерянных объёмов растворов, на каждом сорте сои определяли площадь покрытия листьями поверхности почвы путём измерения суммарной площади проекции листьев на почву способом, изложенным в патенте РФ на изобретение № 2759769 [35]. После определения параметров ширины общей проекции листьев на почву в рядах ( а ), степени покрытия листьями затенённой части почвы ( b ) и с учётом ширины междурядий ( l ) производили расчёт суммарной площади проекции листьев каждого сорта на почву ( R ) по формуле:
a (—)
R = —10^.100, (1);
где а – ширина общей проекции листьев на почву в ряду, см;
b – степень покрытия листьями затенённой части почвы;
l – ширина междурядий, см;
R – суммарная площадь проекции листьев на почву в процентах от общей площади посева.
Рассчитанную для каждого сорта площадь покрытия в процентах от общей площади посева использовали для корректировки рабочих концентраций растворов гиббереллина таким образом, чтобы на покрытую листьями площадь поверхности поля попадал раствор гиббереллина в заданной концентрации и объёме.
После полного созревания растений сои все делянки обмолачивали селекционным комбайном. Для морфометрических измерений в каждом сорте, варианте и повторности рендомизированно отбирали сноповые образцы в количестве 25 растений каждого образца. В лаборатории проводили морфометрические измерения высоты растений, количества узлов, бобов и семян. Отдельно, для определения динамики длин междоузлий в пределах главного побега, на рендоми-зированно отобранных 10 контрольных и 10 обработанных гиббереллином растениях сорта Пума измеряли длину междоузлий.
Статистический анализ различий между вариантами при изучении экзогенной регуляции признака фотопериодической чувствительности выполняли общепринятыми методами вычисления выборочных средних и ошибки выборочных средних х̅ ± S х̅ , а также с использованием алгоритма однофакторного дисперсионного анализа экспериментальных данных.
Результаты и обсуждение. В физиологическую основу гипотезы экзогенного регулирования чувствительности сои к длинам дня была положена теория гор40
монального развития растений М.Х. Чай-лахяна [21; 23; 24; 25; 26]. Согласно этой теории, вегетирующие растения переходят к стадии цветения только при накоплении в тканях сбалансированного комплекса эндогенных фитогормонов (флоригенного комплекса), в совокупности приводящих к инициации формирования репродуктивных органов. Чем быстрее накопится и сбалансируется по компонентному составу флоригенный комплекс фитогормонов, тем быстрее растение перейдёт к цветению. У короткодневных растений типа сои, чем короче длина дня в фотопериодически чувствительные стадии онтогенеза, тем быстрее в их тканях накапливается флоригенный комплекс, и наоборот. При летних посевах растения сои развиваются на непрерывно укорачивающихся длинах дня, что также влечёт за собой ускоренное формирование флоригенного комплекса, раннее зацветание, более ранее прекращение формирования дополнительных узлов и уменьшение высоты растений.
На основе анализа литературы нами была сформулирована рабочая гипотеза, предполагающая возможность нейтрализации фотопериодической чувствительности сои за счёт искусственного экзогенного дисбаланса флоригенного комплекса фитогормонов в виде сдвига наступления фазы цветения на более поздний период и пролонгации фазы вегетативного роста в короткодневных условиях. На основе этой гипотезы предполагалось, что если в период активного синтеза и накопления фитогормонов фло-ригенного комплекса у сои дополнительно нанести на растения раствор одного из его компонентов, то за счёт сложения эндогенной и экзогенной долей того или иного фитогормона, необходимого для закладки цветочных почек, фитогормо-нальный баланс будет нарушен. На восстановление баланса за счёт синтеза дополнительных объёмов остальных компонентов флоригена, вполне очевидно, потребуется определённое время. В этот период на фоне временно разбалансированного и неактивного флоригенного комплекса будет продолжаться вегетативный рост в виде дополнительного формирования на верхушке растений сои очередных узлов. После повторного восстановления флоригенного баланса в этих дополнительных узлах вполне могут сформироваться дополнительные бобы и семена.
Анализ ранее опубликованной литературы показал определённую эффективность экзогенной регуляции чувствительности сои к укороченным длинам дня при обработке растений растворами гиббереллина в период цветения. Этот период, как наиболее чувствительная стадия для применения экзогенных фитогормонов, прежде всего гиббереллина, для регуляции начала цветения, вполне адекватно объясняется гормональной теорией развития растений М.Х. Чайлахяна. Однако сам факт фиксации стадии цветения растений сои указывает на уже сформировавшийся флоригенный комплекс, ставший катализатором развития недифференцированных меристем в направлении формирования цветочных бутонов. Следовательно, для более эффективной нейтрализации фотопериодической чувствительности сои в виде пролонгации вегетативного роста и увеличения высоты растений гормональная разбалансировка флоригенного комплекса должна быть осуществлена ещё до начала цветения.
Для проверки этой гипотезы в 2019 г. в летнем посеве (3-я декада июня) на двух очень ранних сортах сои Пума и Баргузин был заложен эксперимент по уточнению наиболее чувствительных стадий развития сои к экзогенному регулированию их фотопериодической чувствительности, а также определения наиболее эффективных рабочих концентраций растворов гиббереллина. С этой целью растворы ГА 3 в концентрациях 0,0003 %, 0,003 % и 0,03 % наносили на листья растений сортов сои Пума и Баргузин при достижении ими вегетативных стадий формирования 2-х и 4-х тройчатосложных листьев (стадии V 2 и V 4 ), а также при достижении репродуктивной стадии онтогенеза – стадии полного цветения (R 2 ).
Первичный ростостимулирующий эффект от применения растворов ГА 3 в виде удлинения междоузлий, увеличения высоты и некоторого осветления зелёной окраски растений проявился уже через неделю после обработки. Самые заметные фенотипические изменения у растений сои были отмечены при обработке растворами гиббереллина в концентрациях 0,003 и 0,03 % в вегетативную стадию формирования 4-х тройчатосложных листьев (V 4 ) перед началом бутонизации (рис. 1).

Рисунок 1 – Первичный ростостимулирующий эффект через 7 суток после применения раствора гиббереллина в концентрации 0,003 % на стадии V 4 , сорт сои Баргузин, 2019 г.
(на переднем плане видны три ряда обработанных раствором ГА 3 растений, на заднем – контрольные растения, обработанные водой)
Эффект запаздывания наступления цветения на обоих сортах на разных вариантах с обработкой ГА 3 относительно контролей при обработке растений в вегетативных стадиях V 2 и V 4 при применении 0,0003%-ных растворов варьировал от 2–3 суток до 4–10 суток при применении 0,03%-ных растворов.
Визуальные различия по высоте растений между контрольными и обработанными вариантами на сортах Пума и Баргузин сохранялись вплоть до созревания. Максимальная высота растений обоих сортов зафиксирована на вариантах с применением растворов ГА3 в концентрациях 0,003 и 0,03 % в стадиях 2-х и 4-х тройчатосложных листьев (стадии V2 и V4) (рис. 2). Каких-либо заметных разли- чий по высоте созревших растений при обработке растворами ГА3 относительно контрольных вариантов в репродуктивную стадию R2 выявлено не было.

Сорт сои Пума

Сорт сои Баргузин
Рисунок 2 – Фенотипические различия по высоте между контрольными и обработанными растворами ГА 3 на стадии 4-го тройчатосложного листа (вегетативная стадия V 4 ) зрелыми растениями сортов сои Пума и Баргузин, 2019 г.:
1 – контрольные (обработка водой) растения;
2 – растения, обработанные растворами ГА 3 в концентрации 0,0003 %;
3 – растения, обработанные растворами ГА 3 в концентрации 0,003 %;
4 – растения, обработанные растворами ГА 3 в концентрации 0,03 %.
Кроме этого, среди растений на этих вариантах выделялись отдельные, ещё более высокорослые особи. Возможно, метаболизм этих растений оказался ещё более отзывчивым на экзогенную фито-гормональную регуляцию фотопериодической чувствительности (рис. 3). Отдельные, более высокорослые растения на вариантах с обработкой растворами ГА 3 также визуально различимы на рисунках 1 и 2. Все подобные растения представляли селекционный интерес и были индивидуально отобраны для дальнейшего изучения.

Контроль Раствор ГА 3 (вода) 0,003 %
Рисунок 3 – Фенотипические различия по высоте между контрольным и выделившимся по признаку высокорослости растением в варианте с обработкой 0,003%-ным раствором ГА 3 на стадии 4-х тройчатосложных листьев, сорт сои Пума, 2019 г.
Морфометрический анализ выборочных средних по признакам высоты созревших растений, количества узлов на главном стебле, количества бобов и семян на растениях, а также их продуктивности позволил более детально установить различия между контрольными и обработанными растворами ГА 3 на разных стадиях онтогенеза растениями (табл. 1).
Таблица 1
Влияние обработки растений сортов сои Пума и Баргузин растворами гиббереллина (ГА 3 ) на морфометрические показатели в стадии созревания при летнем сроке посева на фоне укорачивающихся длин дня
ФГБНУ ФНЦ ВНИИМК, г. Краснодар, 2019 г.
|
Сорт |
Концентрация раствора ГА 3 , % |
Стадия онтогенеза растений при обработке ГА 3 |
Средние значения хозяйственно ценных признаков* |
||||
|
высота растения, см |
количество на растении, шт. |
продуктивность растения, г |
|||||
|
узлов на главном побеге |
бобов |
семян |
|||||
|
Пума |
Контроль (вода) |
Начало вегетации (2-й лист) |
83,7 ± 2,8 |
16,4 ± 0,6 |
14,3 ± 7,6 |
32,0 ± 5,5 |
6,2 ± 0,8 |
|
Бутонизация (4-й лист) |
84,0 ± 3,3 |
16,2 ± 0,4 |
14,0 ± 5,0 |
30,2 ± 4,4 |
6,9 ± 0,8 |
||
|
Полное цветение (8–9 лист) |
84,6 ± 3,1 |
16,2 ± 0,3 |
14,5 ± 2,7 |
30,6 ± 5,9 |
6,1 ± 0,9 |
||
|
0,0003 |
Начало вегетации (2-й лист) |
86,2 ± 2,3 |
16,0 ± 0,6 |
17,2 ± 8,3 |
33,4 ± 6,5 |
7,8 ± 1,0 |
|
|
Бутонизация (4-й лист) |
82,5 ± 4,7 |
16,2 ± 0,7 |
17,3 ± 5,5 |
34,2 ± 5,8 |
7,6 ± 1,1 |
||
|
Полное цветение (8–9 лист) |
84,9 ± 3,8 |
16,3 ± 0,4 |
18,5 ± 2,5 |
35,4 ± 5,6 |
7,9 ± 0,9 |
||
|
0,003 |
Начало вегетации (2-й лист) |
97,3 ± 4,4 |
16,0 ± 0,6 |
27,4 ± 6,0 |
51,3 ± 4,5 |
9,2 ± 1,3 |
|
|
Бутонизация (4-й лист) |
118,2 ± 5,6 |
19,4 ± 0,6 |
32,1 ± 5,1 |
75,6 ± 4,9 |
12,4 ± 1,5 |
||
|
Полное цветение (8–9 лист) |
87,4 ± 3,7 |
16,7 ± 0,4 |
18,5 ± 3,2 |
32,2 ± 5,0 |
7,9 ± 1,2 |
||
|
0,03 |
Начало вегетации (2-й лист) |
94,6 ± 3,8 |
16,3 ± 0,5 |
26,4 ± 5,9 |
49,5 ± 5,8 |
9,1 ± 0,9 |
|
|
Бутонизация (4-й лист) |
121,5 ± 3,2 |
16,8 ± 0,6 |
28,6 ± 5,7 |
57,9 ± 5,3 |
11,8 ± 1,1 |
||
|
Полное цветение (8–9 лист) |
88,6 ± 3,8 |
16,3 ± 0,4 |
19,3 ± 2,7 |
39,5 ± 5,7 |
8,2 ± 1,0 |
||
|
Баргузин |
контроль (вода) |
Начало вегетации (2-й лист) |
76,9 ± 3,6 |
14,9 ± 0,4 |
14,4 ± 7,7 |
20,5 ± 3,2 |
5,5 ± 0,6 |
|
Бутонизация (4-й лист) |
76,8 ± 3,8 |
14,6 ± 0,5 |
15,1 ± 6,4 |
21,6 ± 2,7 |
5,8 ± 0,4 |
||
|
Полное цветение (8–9 лист) |
77,8 ± 5,7 |
14,7 ± 0,5 |
13,9 ± 4,8 |
21,1 ± 3,6 |
5,0 ± 1,5 |
||
|
0,0003 |
Начало вегетации (2-й лист) |
76,1 ± 4,5 |
15,0 ± 0,4 |
14,7 ± 7,6 |
21,4 ± 3,6 |
7,5 ± 0,6 |
|
|
Бутонизация (4-й лист) |
79,3 ± 4,4 |
15,9 ± 0,4 |
17,3 ± 6,1 |
23,5 ± 2,4 |
8,7 ± 0,8 |
||
|
Полное цветение (8–9 лист) |
77,7 ± 3,8 |
15,2 ± 0,5 |
14,4 ± 4,6 |
22,2 ± 2,6 |
70,3 ± 1,4 |
||
|
0,003 |
Начало вегетации (2-й лист) |
81,4 ± 5,0 |
16,1 ± 0,4 |
25,0 ± 5,6 |
42,3 ± 3,3 |
6,0 ± 1,3 |
|
|
Бутонизация (4-й лист) |
96,7 ± 5,7 |
18,8 ± 0,6 |
29,2 ± 4,1 |
49,6 ± 4,2 |
9,4 ± 1,6 |
||
|
Полное цветение (8–9 лист) |
77,2 ± 3,5 |
15,7 ± 0,4 |
16,5 ± 4,4 |
28,0 ± 3,4 |
5,8 ± 1,6 |
||
|
0,03 |
Начало вегетации (2-й лист) |
80,1 ± 4,2 |
16,3 ± 0,5 |
18,9 ± 3,6 |
29,1 ± 4,2 |
6,3 ± 0,7 |
|
|
Бутонизация (4-й лист) |
85,8 ± 5,3 |
17,3 ± 0,5 |
23,0 ± 4,0 |
40,8 ± 3,6 |
9,0 ± 1,3 |
||
|
Полное цветение (8–9 лист) |
78,4 ± 4,6 |
14,9 ± 0,4 |
19,7 ± 3,3 |
32,2 ± 3,8 |
6,2 ± 1,2 |
||
-
- здесь и далее выборочные средние ± ошибка выборочных средних ( х ± S x )
Контрольные, обработанные водой растения летнего срока посева на фоне укорачивающихся длин дня сорта Пума формировали среднюю высоту 83,7– 84,6 см. Среднее количество узлов на главных побегах контрольных растений – 16,2–16,4 шт., среднее количество бобов с растения – 14,0–14,5 , семян – 30,2–32,0 шт. Средняя продуктивность контрольных растений – 6,1–6,9 г/раст.
Среди вариантов по сроку применения и концентрации растворов гиббереллина лучшим оказался вариант с применением 0,03%-го раствора ГА 3 в стадии онтогенеза V 4 (4-й тройчатосложный лист, начало бутонизации). Высота растений в этом варианте составила 121,5 ± 3,2 см, что превышало контроль в среднем на 37,4 см. Близкими к этим значениям были показатели высоты растений (118,2 ± 5,6 см) в варианте с применением 0,003%-го раствора ГА 3 в той же стадии онтогенеза V 4 . Исходя из величин ошибок выборочных средних, в вариантах с 0,03 и 0,003%-ным растворами ГА 3 на уровне 3,2 и 5,6 см, можно считать средние значения показателей высоты растений в этих вариантах статистически не отличающимися. Все остальные морфометрические параметры у растений сорта Пума оказались максимальными в варианте обработкой 0,003%-ным раствором гиббереллина. Так, среднее количество узлов на главном стебле у таких растений достигало 19,4 ± 0,6 шт., что было выше контрольных растений на 3,0–3,2 шт. Среднее количество бобов на растении в варианте 0,003 % ГА 3 (V 4 ) составило 32,1 ± 5,1 шт., что превышало контрольные показатели на 17,6– 18,1 шт./раст. Среднее количество семян в варианте 0,003 % ГА 3 (V 4 ) достигало 75,6 ± 4,9 шт./раст., что оказалось в 2 раза выше, чем у контрольных растений. Увеличенное количество бобов и семян в варианте 0,003 % ГА 3 (V 4 ), соответственно, обеспечило повышенную продуктивность таких растений, которая составила 12,4 ± 1,5 г/раст., что на 5,5 г/раст. выше контроля.
Варианты обработок растворами гиббереллина в концентрациях 0,0003 и
0,03 % на вегетативных стадиях онтогенеза V 2 и V 4 также обеспечивали увеличенные значения всех морфометрических показателей, но по абсолютным значениям уступали варианту применения растворов ГА 3 в концентрации 0,003 % на стадии 4-х тройчатосложных листьев (V 4 ).
У контрольных растений сорта Баргузин средняя высота варьировала в пределах 76,8–77,8 см. Среднее количество узлов на главных побегах контрольных растений – 14,6–14,9 шт./раст., среднее количество бобов – 13,9–15,1, семян – 20,5–21,6 шт./раст. Средняя продуктивность контрольных растений сорта Баргузин составила 5,0–5,8 г/раст.
Применение растворов гиббереллина в концентрации 0,003 % на стадии 4-х тройчатосложных листьев у этого сорта также оказало максимальный ростостимулирующий эффект. Высота растений на этом варианте составила 96,7 ± 5,7 см, что превышало контроль на 18,9–19,9 см. Среднее количество узлов на главном стебле у растений этого варианта достигало 18,8 ± 0,6 шт., что было выше, чем у контрольных растений, на 3,9–4,2 шт. Среднее количество бобов на растениях достигало 29,2 ± 4,1 шт., что превышало контрольные показатели на 14,1– 15,9 шт./раст. Среднее количество семян составило 49,6 ± 4,2 шт./раст., что было выше, чем у контрольных растений, на 28,0–28,5 шт./раст. Продуктивность растений этого варианта у сорта Баргузин составила 9,4 ± 1,6 г/раст., превысив контрольные показатели на 3,8–4,4 г/раст., или на 62–88 %.
Так же, как и на сорте Пума, на растения сорта Баргузин положительное, хотя и более слабое воздействие, оказали варианты обработок растворами ГА 3 в концентрациях 0,0003 и 0,03 % на вегетативных стадиях онтогенеза V 2 и V 4 .
Применение растворов гиббереллина в фазе полного цветения (R2) на обоих сортах Пума и Баргузин оказало незначительное стимулирующее влияние на все морфометрические показатели. В рамках нашей гипотезы этот срок обработки растворами ГА3 оказался излишне поздним, поскольку ранее сформировавшийся фло-ригенный комплекс эндогенных фитогормонов уже вызвал формирование репродуктивных органов.
В целом анализ первичных данных структурного анализа растений сои сортов Пума и Баргузин показал, что наиболее выраженным ростостимулирующим эффектом, обеспечивающим увеличение всех морфометрических показателей, включая элементы продуктивности растений, является обработка растворами гиббереллина на вегетативной стадии онтогенеза V 4 (4-й тройчатосложный лист) в концентрации 0,003 %. Полученный эффект можно предварительно соотнести с экзогенно обусловленным, временным снижением фотопериодической реакции сои на ускоряющие начало цветения и уменьшающие количество узлов и высоту растений укороченные позднелетние световые фотопериоды.
Ранее в наших исследованиях были изучены особенности фотопериодической реакции растений сои на неоптимальные длины дня, которые могут быть выявлены на основе анализа динамики ярусного изменения длин междоузлий в пределах главного стебля растения сои, а также характера отклонений от нормального распределения этого параметра [13; 36]. Было установлено, что чем адаптивнее генотип растения сои к сложившимся длинам дня в течение вегетации, тем симметричнее формируется кривая распределения длин междоузлий в пределах растения. И наоборот, чем менее асимметричнее кривая распределения этих показателей, тем сильнее отличались от оптимума складывающиеся в течение вегетативного роста световые фотопериоды. Тогда же был предложен полевой экспресс-метод визуальной оценки степени адаптивности сои к складывающимся длинам дня. Если сумма длин трех верхних междоузлий завершившего рост растения сои примерно равна сумме длин трех нижних междоузлий, то такое растение адаптировано к данному фотопериоду. Если сумма длин верхних междоузлий меньше суммы длин нижних междоузлий, то такое растение формировалось в условиях укороченных длин дня [13; 36].
В настоящих исследованиях также был применён ранее разработанный нами алгоритм определения фотопериодической реакции на неоптимальные длины дня. У 10 растений сорта Пума с одинаковым количеством междоузлий, рендомизиро-ванно отобранных на контрольном варианте и на варианте с применением 0,003%-го раствора ГА 3 в стадии онтогенеза V 4 , были измерены и усреднены длины междоузлий в пределах главных побегов. Динамика распределения этих показателей по главным побегам растений в виде кривых представлена на рисунке 4.
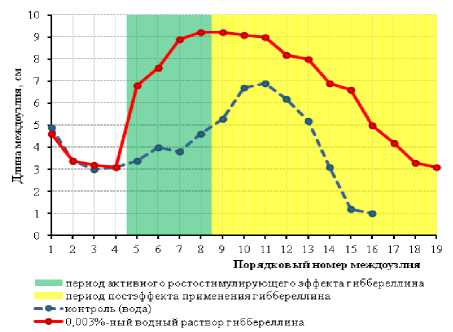
Рисунок 4 – Влияние 0,003%-ного водного раствора ГА 3 на динамику длин междоузлий в пределах главных побегов растений сорта сои Пума, формирующихся на фоне укорачивающихся длин дня, 2019 г.
Как следует из данных, представленных на рисунке 4, сумма длин трех нижних междоузлий у контрольных растений сорта Пума в среднем составляла 11,3 см, а сумма самых верхних междоузлий оказалась в 2 раза меньше – 5,3 см. Такое соотношение сумм длин верхних и нижних междоузлий свидетельствует о негативном влиянии укорачивающихся длин дня в конце вегетативного роста растений, в 45
результате которого формируются более короткие верхние междоузлия. Кривая распределения длин междоузлий у контрольных растений имеет выраженную асимметричность в виде правого скоса.
Сумма длин трех нижних междоузлий у растений этого же сорта Пума, обработанных 0,003%-ным раствором ГА 3 на вегетативной стадии онтогенеза V 4 , составила 11,2 см. При этом сумма длин трех верхних междоузлий, в отличие от контрольного варианта, достигла 10,6 см. Кривая распределения длин междоузлий в пределах главного побега также визуально выглядит симметричнее по сравнению с контрольными растениями.
В целом применение раствора гиббереллина после завершения примерно двухнедельного периода активного ростостимулирующего эффекта гиббереллина на фоне укорачивающихся длин дня привело к более симметричному распределению длин междоузлий в пределах обработанных растений. Такой тип распределения длин междоузлий и соотношения сумм длин нижних и верхних междоузлий на растениях соответствуют ослабленной фотопериодической чувствительности сои в условиях неоптимальных (укороченных) фотопериодов. Это является дополнительным подтверждением гипотезы о возможности экзогенной фитогормональной регуляции фотопериодической чувствительности.
Для практического подтверждения возможности экзогенной регуляции фотопериодической чувствительности сои в 2020 г. был заложен эксперимент с применением растворов с наиболее эффективной (0,003 %) концентрацией гиббереллина. Сорта Пума и Баргузин были посеяны 24 июня после уборки озимого ячменя на делянках площадью 117 м2 в 3-кратной повторности. Всходы появились 30 июня при длине дня 15 ч 34 мин. Обработку 0,003%-ным раствором ГА3 осуществляли на стадии формирования 4-х тройчатосложных листьев. Наиболее активный рост растений изучаемых сортов проходил на фоне укорачивающихся длин дня – 15 ч 15 мин – 14 ч 15 мин. Созрева- ние сортов Пума и Баргузин отмечено в 3-й декаде сентября при длине дня 14 ч 00 мин – 13 ч 50 мин.
В условиях 2020 г. обработанные гиббереллином и контрольные варианты обоих сортов сои созрели практически одновременно. После полного созревания растений на всех делянках в конце сентября учитывали урожайность контрольного (необработанного) и обработанного ГА 3 вариантов (табл. 2).
Таблица 2
Влияние внесения 0,003%-ных растворов ГА з на морфометрические показатели растений в стадии созревания и урожайность у сортов сои Пума и Баргузин при посеве в 3-й декаде июня
ФГБНУ ФНЦ ВНИИМК, г. Краснодар, 2020 г.
|
Сорт сои |
Вариант |
Высота растения, см |
Кол-во узлов на главном побеге, шт. |
Кол-во семян на растении, шт. |
Урожайность |
|
|
т/га |
откл. от контроля, ± Δ |
|||||
|
Пума |
Контроль (вода) |
66,5 ± 2,0 |
10,3 ± 0,4 |
34,1 ± 3,1 |
0,91 |
– |
|
0,003%-й раствор ГА 3 |
106,0 ± 8,1 |
14,4 ± 0,5 |
66,2 ± 6,9 |
1,34 |
+0,43 |
|
|
Баргузин |
Контроль (вода) |
44, 5± 1,3 |
9,5 ± 0,3 |
38,9 ± 4,2 |
0,68 |
– |
|
0,003%-й раствор ГА 3 |
63,5 ± 2,4 |
12,2 ± 0,5 |
54,6 ± 5,1 |
1,03 |
+0,35 |
|
|
НСР 05 |
– |
– |
– |
– |
0,31 |
|
Анализ таблицы 2 показывает, что конечная высота обработанных ГА 3 растений сорта Пума достигла 106,0 ± 8,1 см, что на 39,5 см больше, чем средняя высота контрольных растений. Количество узлов на главных побегах обработанных растений в среднем составило 14,4 ± 0,5 шт./раст. У контрольных растений этот показатель не превышал 10,3 ± 0,4 шт./га. На обработанных гиббереллином растениях сформировалось в среднем по 66,2 ± 6,9 семян, что почти в 2 раза выше, чем у контрольных (34,1 ± 3,1 шт./раст.). Соответственно, увеличенное количество семян на растениях, обработанных 0,003%-ным раствором ГА 3 обеспечило прибавку урожая в этом варианте 0,43 т/га, или +47,3 %.
Аналогичные закономерности выявлены и на сорте сои Баргузин. Прибавка вы- соты у обработанных ГА3 растений по сравнению с контролем составила +19 см. Количество узлов на главном побеге превысило контроль в среднем на 2,7 шт./раст., количество семян – на 15,7 шт./раст. Урожайность контрольного, без обработки, варианта сорта Баргузин составила 0,68 т/га, а экспериментального, с обработкой 0,003%-ным раствором ГА3, – увеличилась до 1,03 т/га, что обеспечило прибавку урожая 0,35 т/га.
Невысокая урожайность контрольных и обработанных ГА 3 сортов сои Пума и Баргузин при летнем посеве определялась дефицитом осадков в июне и в августе, ограничившим формирование бобов и семян. Тем не менее даже в таких погодных условиях экзогенная регуляция фотопериодической чувствительности за счёт обработки раствором гиббереллина вызвала заметное увеличение высоты растений, количества узлов и семян, что обеспечило достоверное увеличение продуктивности изучаемых сортов.
На основании проведённых в 2019– 2020 гг. исследований и полученных результатов по разработке экзогенной регуляции фотопериодической чувствительности сои нами в Роспатент была подана заявка и в 2021 г. получен патент РФ на изобретение № 2759769 «Способ повышения урожайности сои на фоне укороченных длин дня» [35].
В 2021–2022 гг. исследования по экзогенной регуляции фотопериодической чувствительности сои путём обработки растений 0,003%-ными растворами гиббереллина были продолжены.
В 2021 г. для снижения модифицирующего эффекта недостатка влаги в почве летний посев сои осуществляли в 3-й декаде июня на парующем участке. Дополнительным фактором в наших исследованиях стало включение в эксперименты ещё двух сортов – очень раннего, высоко фотопериодически чувствительного сорта сои Вита и раннего, слабо фотопериодически чувствительного сорта Славия.
Посев осуществляли 21 июня на делянках площадью 108 м2 в 3-кратной повторности. Всходы всех сортов получены 28 июня при длине дня 15 ч 35 мин. Обработку 0,003%-ным раствором ГА 3 осуществляли во 2-й декаде июля в фазе формирования 4-х тройчатосложных листьев при длине дня 15 ч 08 мин. Наиболее активный рост растений всех изучаемых сортов проходил на фоне укорачивающихся длин дня – 15 ч 10 мин – 14 ч 20 мин. После полного созревания растений в конце сентября учитывали урожайность контрольного (необработанного) и обработанного ГА 3 вариантов у каждого сорта (табл. 3).
Таблица 3
Влияние внесения растворов ГА 3 на морфометрические показатели растений в стадии созревания и урожайность у сортов сои при посеве в 3-й декаде июня на парующем участке
ФГБНУ ФНЦ ВНИИМК, г. Краснодар, 2021 г.
|
Сорт сои |
Вариант |
Высота растения |
Урожайность |
||
|
см |
откл. от контроля, ± Δ |
т/га |
откл. от контроля, ± Δ |
||
|
Пума |
Контроль (вода) |
93 |
– |
2,23 |
– |
|
0,003%-й раствор ГА 3 |
101 |
+8 |
2,48 |
+0,25 |
|
|
Баргузин |
Контроль (вода) |
74 |
– |
1,75 |
– |
|
0,003%-й раствор ГА 3 |
85 |
+11 |
2,12 |
+0,37 |
|
|
Вита |
Контроль (вода) |
76 |
– |
2,08 |
– |
|
0,003%-й раствор ГА 3 |
85 |
+9 |
3,30 |
+0,22 |
|
|
Славия |
Контроль (вода) |
105 |
– |
2,55 |
– |
|
0,003%-й раствор ГА 3 |
108 |
+3 |
2,63 |
+0,08 |
|
|
НСР 05 |
– |
– |
– |
0,19 |
– |
Как и в предыдущем 2020 г., у сорта Пума в варианте с обработкой растений 0,003%-ными растворами гиббереллина отмечено заметное увеличение высоты растений (+8 см) по сравнению с контрольными, обработанными водой растениями. У самого раннего и самого низкорослого в опыте сорта Баргузин прибавка высоты обработанных ГА3 растений по сравнению с контрольными составила 11 см (рис. 5).

Рисунок 5 – Сравнительная высота обработанных 0,003%-ным раствором гиббереллина (слева) и контрольных растений (справа) летнего посева сорта
Баргузин на стадии завершения вегетативного роста, 2021 г.
На 9 см относительно контроля увеличилась высота обработанных растворами ГА 3 растений сорта Вита. Однако высота растений сорта Славия в результате применения ГА 3 увеличилась всего на 3 см. Это можно объяснить собственной, генетически обусловленной пониженной фотопериодической чувствительностью этого сорта.
Средняя по всем сортам и вариантам урожайность в эксперименте 2021 г. составила 2,39 т/га, что более чем в 2 раза превышало среднюю урожайность аналогичного эксперимента в 2020 г. Вполне очевидно, что это было обусловлено достаточными запасами влаги на парующем участке, а также обильными осадками в июне и в августе – 126,8 и 88,4 мм соответственно.
Тем не менее даже в благоприятных по влагообеспеченности условиях, применение 0,003%-ных растворов ГА 3 за счёт экзогенной нейтрализации негативного влияния укороченных длин дня оказало положительный эффект на урожайность всех изучаемых сортов. Прибавка урожайности от применения ГА 3 на сорте
Пума составила +0,25 т/га, на сорте Баргузин – +0,37 т/га; на сорте Вита – +0,22 т/га. На сорте Славия, отличающемся пониженной фотопериодической чувствительностью, прибавка урожайности оказалась самой низкой в опыте – всего +0,08 т/га. Тем не менее даже у этого сорта был выявлен положительный эффект от экзогенной нейтрализации негативного эффекта укорачивающихся фотопериодов, обеспечивший самую высокую в опыте урожайность – 2,63 т/га.
В 2022 г., с целью изучения реакции изучаемых сортов на экзогенную регуляцию фотопериодической чувствительности на фоне ещё более коротких длин дня, их посев был осуществлён после уборки ячменя, но на 2 недели позже – 4 июля. Однако вследствие двукратного поверхностного предпосевного лущения стерни на фоне отсутствия осадков в этот период верхний горизонт почвы оказался излишне пересушен. Основная часть всходов появилась 11 июля при длине дня 15 ч 23 мин, оставшаяся часть всходов появилась только к 18–20 июля при длине дня 15 ч 12 мин –15 ч 08 мин. Неодновременное появление всходов в пределах каждого сорта повлекло за собой невыро-вненность растений по стадиям онтогенеза. При достижении большей частью растений стадии онтогенеза V 4 (при длине дня 14 ч 20 мин) и наступлении срока их обработки растворами гиббереллина часть поздно взошедших растений находилась на стадиях от V 2 до V 3 , что повлекло за собой некоторое снижение эффективности от обработок растворами ГА 3 .
Более поздние посевы сои в 2022 г. и их наиболее активный рост проходили на фоне ещё более коротких по сравнению с предыдущим годом длин дня – 14 ч 00 мин – 13 ч 20 мин. Следствием этого стало формирование более низких растений на контрольных вариантах всех изучаемых сортов. Так, если в 2021 г. высота контрольных растений сорта Пума составляла 93 см, то в условиях 2022 г. их высота не превышала 77 см. Высота растений сорта Баргузин в 2021 г. достигала 74 см, а в 2022 г. – 67 см. У сорта Вита – 76 и 65 см; у сорта Славия – 105 и 85 см соответственно. Однако даже в таких, ещё более короткодневных условиях, фотопериодическая чувствительность у всех четырех сортов сои была частично нейтрализована обработкой 0,003%-ными растворами гиббереллина, что проявилось в увеличении высоты растений и их урожайности (табл. 4).
Таблица 4
Влияние внесения растворов ГА 3 на морфометрические показатели растений в стадии созревания и урожайность у сортов сои при посеве в 1-й декаде июля
ФГБНУ ФНЦ ВНИИМК, г. Краснодар, 2022 г.
|
Сорт сои |
Вариант |
Высота растений |
Урожайность |
||
|
см |
откл. от контроля, ± Δ |
т/га |
откл. от контроля, ± Δ |
||
|
Пума |
Контроль (вода) |
77 |
– |
0,92 |
– |
|
0,003%-й раствор ГА 3 |
95 |
+18 |
1,18 |
+0,22 |
|
|
Баргузин |
Контроль (вода) |
67 |
– |
1,00 |
– |
|
0,003%-й раствор ГА 3 |
83 |
+16 |
1,24 |
+0,24 |
|
|
Вита |
Контроль (вода) |
65 |
– |
1,02 |
– |
|
0,003%-й раствор ГА 3 |
78 |
+7 |
1,31 |
+0,16 |
|
|
Славия |
Контроль (вода) |
85 |
– |
1,47 |
– |
|
0,003%-й раствор ГА 3 |
96 |
+11 |
1,54 |
+0,07 |
|
|
НСР 05 |
– |
– |
– |
0,23 |
– |
Так, у сорта Пума высота обработанных растворами ГА 3 растений достигла 95 см, что оказалось выше контрольных растений на 18 см. Урожайность контрольного варианта этого сорта составила 0,92 т/га, обработанного 0,003%-ными растворами ГА 3 – 1,18 т/га, что на 0,22 т/га (+ 28,3 %) выше, чем в контроле. У сорта Баргузин высота обработанных ГА 3 растений превышала контрольные растения на 16 см и составила 83 см. Урожайность этого сорта на варианте с ГА 3 достигала
-
1,24 т/га, что выше необработанного контроля на 0,24 т/га, или на 24 %. У сорта Вита превышение высоты обработанных ГА 3 растений над контрольными составляло 7 см, а превышение урожайности – 0,16 т/га, или 28,4 % соответственно.
У слабо фотопериодически чувствительного сорта Славия реакция на нейтрализацию негативного эффекта укороченных позднелетних длин дня закономерно была пониженной. Тем не менее увеличение высоты обработанных 0,003%-ными растворами гиббереллина растений относительно контроля составило +11 см. Прибавка урожайности у этого сорта оказалась незначительной – всего 0,07 т/га, или 4,8 %. Это позволяет сделать вывод о высокой эффективности экзогенной регуляции в виде снижения негативной реакции на укороченные длины дня для фотопериодически чувствительных сортов. У слабо фотопериодически чувствительных сортов сои типа сорта Славия эффект экзогенной регуляции сортовой реакции на неоптимальные длины дня менее выражен.
Выводы . Впервые, на основе гормональной теории развития растений академика М.Х. Чайлахяна разработан способ экзогенной регуляции генетически детерминированного признака высокой фотопериодической чувствительности сои к укороченным длинам дня, вызывающий пролонгацию вегетативного роста растений, увеличение количества узлов, высоты растений и урожайности.
У сои установлена наиболее чувствительная к экзогенной регуляции фотопериодической чувствительности стадия онтогенеза – 4-й тройчатосложный лист (стадия онтогенеза V 4 ), позволяющая вызывать временный дисбаланс соотношения эндогенных фитогормонов флори-генного комплекса за счёт экзогенного нанесения на листья растений растворов гиббереллина.
Определена наиболее активная концентрация растворов гиббереллина – 0,003 %, нанесение которых на листья сои на стадии онтогенеза V4 вызывает пролонгацию вегетативного роста и формирование дополнительных узлов на главных побегах растений.
На сформированных за счёт экзогенной пролонгации вегетативного роста дополнительных узлах растений сои формируются дополнительные бобы и семена.
Экзогенная пролонгация вегетативного роста увеличивает урожайность фотопериодически чувствительных сортов сои при их выращивании в условиях неоптимальных для таких генотипов, укороченных длин дня, в том числе в повторных летних посевах после уборки озимых колосовых культур.
Экзогенная регуляция фотопериодической чувствительности сортов сои с генетически детерминированным признаком пониженной реакции на неоптимальные длины дня при их выращивании на фоне укороченных длин дня, в том числе при летних сроках посева, закономерно менее эффективна.
