Электрическая эквивалентная схема биологических объектов растительного происхождения

Автор: Голев И.М., Санин В.Н., Титов С.А., Коротков Л.Н.
Журнал: Вестник Воронежского государственного университета инженерных технологий @vestnik-vsuet
Рубрика: Биотехнология, бионанотехнология и технология сахаристых продуктов
Статья в выпуске: 4 (62), 2014 года.
Бесплатный доступ
Представлены результаты измерения комплексного электрического сопротивления биологических тканей растительного происхождения. Измерения проводились при Т=296 K в диапазоне частот от 5 до 500 кГц. В качестве электродов использовались покрытые оловом (чистота 99,9%) медные пластины. Экспериментально исследованы следующие объекты: образцы паренхимной ткани яблока в виде цилиндров диаметром 20 мм и длиной 20 мм; яблочный сок, полученный при механическом разрушении клеток; отжатая яблочная мезга (содержание сока не более 20%), полученная центробежной сепарацией, в которой разрушена система клеток. Для растительной ткани с целостной системой клеток в области 10 3 - 10 5 Гц наблюдается ярко выраженный минимум угла фазового сдвига. В случае отсутствия клеток его величина существенно уменьшается. Рассмотрена эквивалентная электрическая схема тканей, проведен расчет всех ее элементов. Эквивалентом емкости двойного электрического слоя на границе раздела металлического измерительного электрода и межклеточной жидкости является элемент С 1. Электрическое сопротивление этого слоя переменному току характеризуется элементом R 1. Цепочка параллельно включенных активного сопротивления и емкости описывает систему растительных клеток. Емкость С 2 обусловлена электрической емкостью мембран клеток, а сопротивление R 2 - электрическим сопротивлением мембран и внутриклеточного пространства. Совпадение экспериментальных и расчетных данных в области частот более 10 3 Гц удовлетворительное. В области меньших частот наблюдаются отличия. Это может быть связано с особенностями поведения двойного электрического слоя. Вместе с тем, в области частот, где проявляются электрические свойства клеточной структуры исследуемой ткани совпадение хорошее, что свидетельствует о справедливости рассматриваемой эквивалентной схемы. Показано, что величина комплексного электрическое сопротивление растительной ткани в диапазоне частот от 10 3 до 10 5 Гц позволяет получить электрические параметры клеточной структуры: характеристическую частоту, электрические емкость и сопротивление, которые на практике могут быть использованы для идентификации продукции и оценки степени влияния различных внешних факторов на ее качество.
Электрическая эквивалентная схема, дисперсии, биологические ткани, характеристическая частота
Короткий адрес: https://sciup.org/14040318
IDR: 14040318 | УДК: 538.9
Electrical equivalent circuit of biological objects of vegetable
The results of measurements of complex biological tissues electrical resistance of vegetable origin are presented. The measurements were performed at T=296 K in the frequency range from 5 to 500 kHz. As the electrodes were covered with tin (purity of 99.9%) copper plates.. Experimentally investigated the following objects: samples parenchymal tissue of Apple in the form of cylinders with a diameter of 20 mm and a length of 20 mm; Apple juice, obtained by mechanical destruction of cells; pressed Apple pulp (juice content of not more than 20%)obtained by the centrifugal separation, which destroyed the system of cells. For plant tissue with a holistic system of cells in the field 10 3 - 10 5 Hz is observed pronounced minimum angle of phase shift. In the absence of cells and its value is greatly reduced.The equivalent electrical circuit fabrics are considered. The calculation of all its elements is made. The equivalent capacitance of the electrical double layer at the interface of metal measuring electrode and extracellular fluid is element of C 1. The electrical resistance of this layer alternating current is characterized by the element R 1. Chain parallel connected resistance and capacitance describes the system of plant cells. The capacitance C 2 is due to the electrical capacity of the cell membranes, and the resistance R 2 is the electrical resistance of the membranes and intracellular space.The coincidence of experimental and calculated data in a frequency range of more than 10 3 Hz satisfactory. In the region of lower frequencies is observed differences. This may be due to the specific behavior of the electrical double layer. However, in the frequency region where the electrical properties of the cell structure of the investigated tissue match good, which proves the validity of the considered equivalent circuit. It is shown that the value of the complex electrical impedance of vegetable tissue in the frequency range from 10 3 Hz to 10 5 allows receiving the cell structure electrical parameters: the characteristic frequency, the electric resistance and capacitance. In practice, these parameters can be used to identify the product and evaluate the influence degree of various external factors on its quality
Текст научной статьи Электрическая эквивалентная схема биологических объектов растительного происхождения
Измерение частотной зависимости электрического импеданса биологических объектов растительного происхождения при воздействии гармонического электрического сигнала дает обширную информацию о таких электрофизических параметрах их тканей как модуль комплексного сопротивления Z , угол фазового сдвига между током и напряжением, активная и реактивная (как правило емкостной) составляющие Z и Z C [1–3].
В настоящее время весьма актуальной является задача установления связи этих параметров с внутренней структурой этих объектов, в частности с параметрами клеток ткани. Это позволило бы перейти к количественным биохимическим характеристикам растительной ткани и существенно повысить эффективность данного метода.
Известно большое количество эквивалентных электрических схем биологических тканей [2], позволяющих интерпретировать экспериментальные результаты измерения их импеданса.
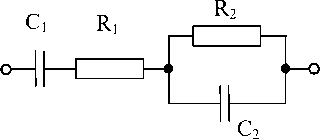
Рисунок 1. Электрическая эквивалентная схема
В работе рассмотрен один из вариантов усовершенствованной эквивалентной схемы (рисунок 1) ткани биологического объекта, представляющего собой систему растительных клеток, заполненных клеточным соком, и межклеточное пространство. В нашей экспериментальной ситуации эквивалентом емкости двойного электрического слоя на границе раздела металлического измерительного электрода и межклеточной жидкости является элемент С1. Электрическое сопротивление этого слоя переменному току характеризуется элементом R1. Цепочка параллельно включенных активного сопротивления и емкости описывает систему растительных клеток. Емкость С2 обусловлена электрической емкостью мембран клеток, асопротивление R2 – электрическим сопротивлением мембран и внутриклеточного пространства [4, 5].
Для данной эквивалентной схемы вели- чину активного сопротивления импеданса можно записать как:
Z r ( ro ) = R +
R 2
1 + ( ® R 2 C 2 )2
а реактивного (емкостного) сопротивления:
Z^. ( ro ) =
1 roR^ 2 C-^
ro C 1 1 + ( roR2 C 2 )2
Модуль электрического импеданса определяется как:
|z\ = \ zR + zC , (3)
а угол фазового сдвига:
Ф = arctan(Z C ) . (4)
ZR
Анализ выражений (1) и (2) позволяет сделать следующие выводы:
• для to ^ю
R 2
формулы значение
i (1) : при стремлении Z r ( to ) = R , т.к.
1 + ( to R 2 C 2 ) 2
^ 0;
• для формулы
to ^^ величина
функция Zc( to ) =
(2) : при
— ^ 0 .
toC1
ro R 2 C2
стремлении
При этом
1 + ( roR2 C 2 ) 2
имеет при
частоте to max
максимум, величина ко- R 2 C 2
торого Z C ( to ) = 0.5 ■ R 2. Определив из экспе
риментальных данных величины R2 и toma;r , 1
можно рассчитать С2 =------;
to pe3 R - 2
• для формулы (2) : при стремлении to ^ 0 соотношение слагаемых в формулы становится следующим
1 to R, 2 C2
--->>----2—2—- и величина toCx 1 + (toR2C2)
Таким образом, из анализа частотных зависимостей Zc ( to ) и ZR ( to ) , можно определить все элементы эквивалентной схемы растительной ткани.
В работе были экспериментально исследованы следующие объекты:
-
- образцы паренхимной ткани яблока в виде цилиндров диаметром 20 мм и длиной 20 мм;
-
- яблочный сок, полученный при механическом разрушении клеток методом центробежной сепарации;
-
- отжатая яблочная мезга (содержание сока не более 20 %), также полученная центробежной сепарацией, в которой разрушена система клеток, но присутствует фрагменты с клеточной структурой.
Измерения проводились при Т=296 K в диапазоне частот от 5 до 500 кГц. В качестве электродов использовались покрытые оловом (чистота 99,9 %) медные пластины.
На рисунках 2 и 3 представлены частотные зависимости модуля электрического импеданса | Z | и угла фазового сдвига ф для вышеуказанных образцов.
ZC (to) «-----. Тогда значения С 1 =------ to ■ с, zc ■ го
.
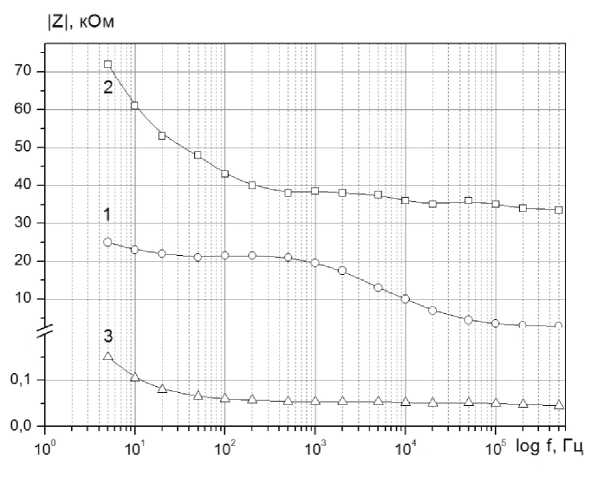
Рисунок 2. Частотная зависимость модуля электрического импеданса Z : 1- ткань яблока; 2 - яблочная мезга; 3 - яблочный сок
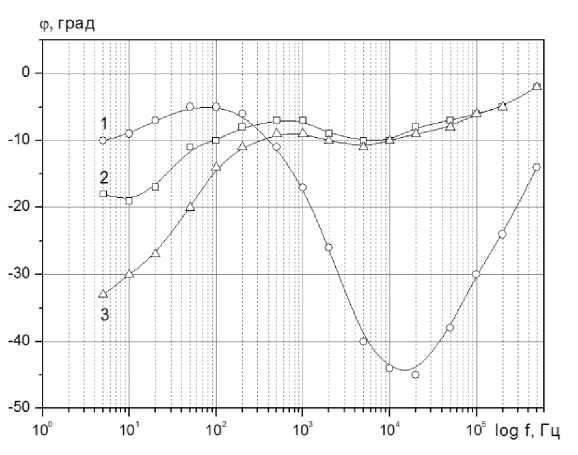
Рисунок 3. Частотная зависимость угла фазового сдвига ф : 1- ткань яблока; 2 - яблочная мезга; 3 - яблочный сок
Обращает на себя тот факт, что частотные зависимости фазового сдвига ф для яблочной ткани с одной стороны и яблочного сока и мезги качественно отличаются (рисунок 3, кривые 1 и 2, 3).Для растительной ткани с целостной системой клеток в области 103-105 Гц наблюдается ярко выраженный минимум угла фазового сдвига. В случае отсутствия клеток его величина существенно уменьшается. Отметим, что и в других экспериментах, когда производилась деструкция клеток тканей растительного происхождения (картофеля, моркови и других овощей и фруктов) путем термической обработки, наблюда- лось аналогичное изменение характера функциональной зависимости ф( f ) [5-8]. Кривые |Z( f) имеют схожий характер (рисунок 2 а) и отличаются лишь количественно. В области высоких частот (> 100 кГц), когда существенно снижается влияние двойного электрического слоя, сопротивление мезги больше сопротивления тканей яблока в 10 раз, а сопротивление сока меньше в 60 раз.
Для удобства анализа результатов измерений представим их по формулам (3) и (4) в виде функций ZR ( to ) и Zc ( to ) (рисунки 4, 5).
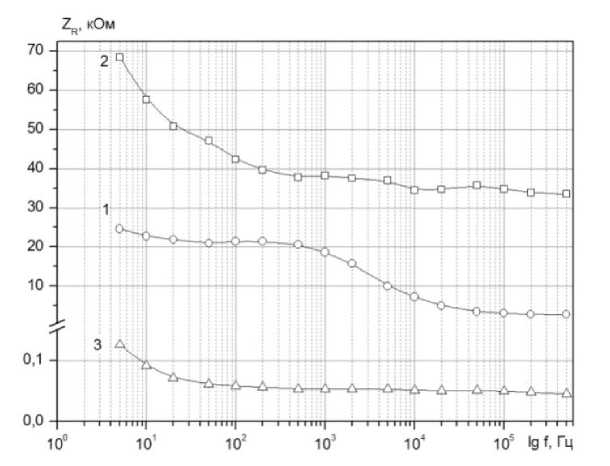
Рисунок 4. Частотная зависимость активного ZR сопротивления импеданса: 1- ткань яблока; 2 - яблочная мезга; 3 - яблочный сок
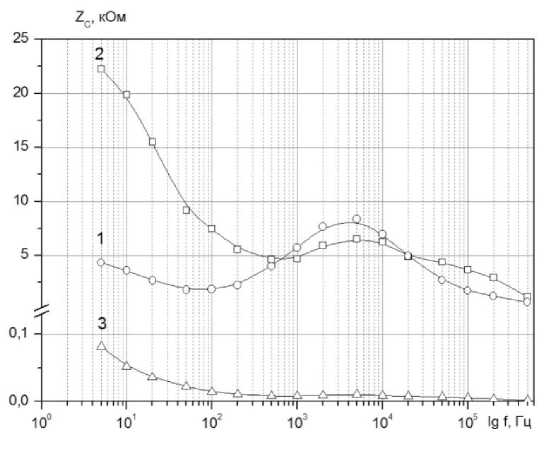
Рисунок 5. Частотная зависимость реактивного Zc сопротивления импеданса: 1- ткань яблока; 2 - яблочная мезга; 3 – яблочный сок
На частотных зависимостях Zc (to) наблюдается характерный экстремум, обусловленный электрофизическими параметрами клеточной ткани. Для яблочной мезги высота пика менее выражена. Важно, что максимумы для кривых 1 и 2 (рисунок 3, б) реализуются при одной и той же частоте f = 4000 Гц, которая является характеристической для данной яблочной ткани.
Используя вышеописанную методику анализа формул (1) и (2), были определены для трех объектов измерений значения элементов эквивалентной электрической схемы (таблица 1).
Т а б л и ц а 1
Величины элементов эквивалентной схемы для ткани яблока, яблочного сока и яблочной мезги
|
Объект исследования |
R1, кОм |
С1, мкФ |
R2, кОм |
С 2 , нФ |
|
Ткань яблока |
2,1 |
1,4 |
16 |
2,45 |
|
Яблочная мезга |
33 |
1,5 |
12,5 |
3,15 |
|
Яблочный сок |
0,045 |
390 |
0,081 |
- |
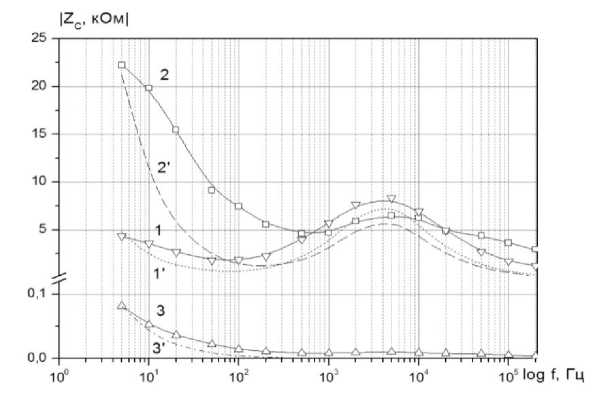
Рисунок 6. Экспериментальные (сплошные линии) и расчетные (прерывистые линии) частотные зависимости реактивного Zc сопротивления импеданса: 1, 1 ‘- ткань яблока; 2, 2‘ - яблочная мезга; 3, 3‘ - яблочный сок
На рисунках 6 и 7 показаны экспериментальные и расчетные значения для зависимостей а ZR( а ) и Z ( а ) .
Видно, что совпадение экспериментальных и расчетных данных в области частот а > 103 Гц удовлетворительное, в области меньших частот имеют место отличия. Это может быть связано с особенностями поведения двойного электрического слоя. Вместе с тем, в области частот, где проявляются электрические свойства клеточной структуры исследуемой ткани совпадение хорошее, что свидетельствует о справедливости для этого случая рассматриваемой эквивалентной схемы.
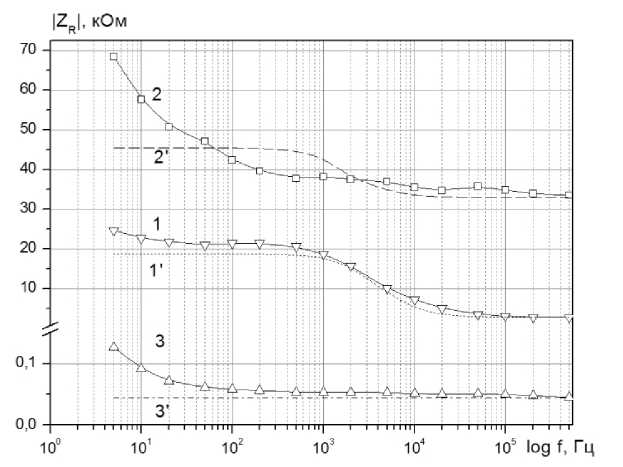
Рисунок 7. Экспериментальные (сплошные линии) и расчетные (прерывистые линии) частотные зависимости активного ZR сопротивления импеданса: 1, 1 ‘- ткань яблока; 2, 2‘ - яблочная мезга; 3, 3‘ - яблочный сок
Нами были проведены измерения и соот- всех элементов электрической эквивалентной ветствующие расчеты характеристических ча- схемы (таблица 2).
стот максимума на зависимостях Zc ( а ) и
Т а б л и ц а 2
Величины элементов эквивалентной схемы для ткани различных плодов
|
Объект исследования, ткани |
Частота характеристическая, Гц |
R 1 , кОм |
С 1 , мкФ |
R 2 , кОм |
С 2 , нФ |
|
Томата |
1900 |
0,075 |
12,3 |
3,2 |
26,4 |
|
Черешни |
2600 |
1,96 |
9,4 |
42,0 |
14,6 |
|
Яблока |
4000 |
2,1 |
1,4 |
16 |
2,45 |
|
Картофеля |
5000 |
0,73 |
14,6 |
1,28 |
25,0 |
|
Огурца |
9000 |
0,14 |
8,2 |
1,7 |
10,6 |
Обращает на себя внимание, что для исследуемых объектов характеристические частоты отличаются в 1,25-4,5 раз. Величины R1, С1, R2 и С2 индивидуальны для каждого объекта исследований, что позволяет проводить идентификацию растительных тканей, имея библиотеку значений.
В заключение отметим, что величина комплексного электрического сопротивления растительной ткани в диапазоне частот от 103 до 105 Гц позволяет получить электрические параметры клеточной структуры: характеристическую частоту, электрические емкость и сопротивление, которые на практике могут быть использованы для идентификации продукции и оценки степени влияния различных внешних факторов на ее качество.
