Эмбриогенез кишечника гибридных свиней семиреченской породы

Автор: Давлетова Л.В.
Журнал: Сельскохозяйственная биология @agrobiology
Рубрика: Морфология отдельных органов и тканей
Статья в выпуске: 2 т.39, 2004 года.
Бесплатный доступ
По динамике морфофункциональных показателей проводили сравнительную оценку развития тонкого и толстого кишечника у эмбрионов гибридных свиней семиреченской породы и исходных родительских форм - дикого кабана европейского подвида и домашних свиней породы ландрас. Обсуждается характер обменных и пищеварительных процессов в кишечнике, а также выраженность типа пищеварения у гибридных животных.
Короткий адрес: https://sciup.org/142132971
IDR: 142132971 | УДК: 636.4:57.017.642:591.434
Intestine embryogenesis in pigs of the hybridous Semirechenskaya breed
On basis of dynamics of morphofunctional parameters the author compared the development of small and large intestines in pigs embryos of the hybridous Semirechenskaya breed and initial parental forms - wild boar of European subspecies and domestic pig of the Landras breed. It was shown, that hybrids have the intermediate type of intestine development with considerable individual variability and irregular growth of structure elements, and also pronounced intestinal-gastric type of digestion. Activities of metabolism and digestion in hybridous embryos conditioned by formation of vacuoles and cavities in villuses enterocytes, scalloped texture of their lateral surfaces, slowed transformation of colon villuses to crypts and also intensive development of duodenal glands. Because in newborn piglets as hybridous as domestic it was recommended to take into account a weakening of protective mechanisms of intestine functioning owing to partial conservation of embryos texture of villuses, participated in metabolism processes.
Текст научной статьи Эмбриогенез кишечника гибридных свиней семиреченской породы
Селекцию гибридных свиней семиреченской породы, созданных в Казахстане посредством скрещивания дикого кабана и домашних свиней разных пород, проводили с целью получения высокопродуктивных гибридов, приспособленных к условиям повышенной инсоляции. При этом добивались темной окраски гибридных животных (как у дикого кабана и свиней породы дюрок, принимавших участие в скрещиваниях), с тем чтобы предохранить их от жары и солнечной радиации (1). Приспособленность этих свиней к природно-климатическим условиям Казахстана обусловлена также морфофункциональными особенностями органов пищеварения.
В задачу нашей работы входила оценка морфофункциональных особенностей тонкого и толстого кишечника эмбрионов гибридных свиней семиреченской породы от раннеплодного периода до рождения по сравнению с родительскими формами.
Методика . Объектом исследования служили эмбрионы свиней семиречен-ской породы и породы ландрас, а также кабанов европейского подвида (в разные возрастные периоды число эмбрионов варьировало). Для оценки гистогенеза кишечника использовали гистологические, гистохимические и биохимические методы (5). Толщина слизистой и мышечной оболочек, слизистого и подслизистого слоев тонкого (двенадцатиперстная и подвздошная кишки) и толстого (ободочная кишка) кишечника представлена как в абсолютных, так и в относительных единицах (% от толщины стенки кишечника). Плотность расположения дуоденальных желез оценивали по их числу на 1 мм2 площади подслизистого слоя (% от 100 определений). О достоверности полученных данных судили по средним показателям с учетом индивидуальной изменчивости (5).
Результаты . Толстый и тонкий кишечник у гибридных свиней семиречен-ской породы (далее гибриды), как и у исходных форм, был представлен слизистой, мышечной и серозной оболочками, толщина которых увеличивалась по мере плодного развития. Мышечная оболочка образована двумя слоями гладких мышечных клеток, слизистая — соединительной тканью и эпителиальными клетками, дифференциация которых проходила по-разному в тонком и толстом кишечнике.
Д в е н а д ц а т и п е р с т н а я к и ш к а. У 40-суточных эмбрионов гибридных свиней семиреченской породы слизистая оболочка двенадцатиперстной кишки была представлена эпителиальными бугорками, состоящими из многорядного эпителия, а также первичными складками, имеющими соединительно-тканную основу, покрытую многорядным эпителием. При расщеплении первичных складок формировались первичные ворсинки, имеющие многорядный эпителий, и истинные ворсинки, у которых соединительно-тканная основа покрыта цилиндрическими эпителиальны-73
ми клетками, расположенными однослойно и имеющими каемку в апикальной части. В раннеплодный период у гибридов прослеживалась полная последовательность закладки и формирования ворсинок, в то время как у 40-суточных эмбрионов домашних и диких свиней — только закладка ворсинок в виде бугорков и первичных складок. Содержание гликогена в эпителиальных клетках у гибридов и исходных форм составляло соответственно 123 и 121-129 мг%.
К 45-суточному возрасту у эмбрионов как гибридов, так и исходных форм в слизистой двенадцатиперстной кишки наблюдалось формирование значительного количества истинных ворсинок. У основания ворсинок происходила закладка дуоденальных желез в виде скопления эпителиальных клеток, внедряющихся в соединительную ткань слизистой оболочки. Среди энтероцитов истинных ворсинок, имеющих каемку, появлялись бокаловидные клетки, которые секретировали нейтральные гликопротеины, выявляемые также в секрете дуоденальных желез и каемке. К возрасту 50 сут в слизистой двенадцатиперстной кишки эмбрионов гибридов и исходных форм значительно удлинялись ворсинки, на верхушках которых энтероциты отходили от соединительно-тканной основы, образуя полости, а также увеличивалось число бокаловидных клеток, в которых наряду с нейтральными были выявлены кислые гликопротеины. Последние были обнаружены в секрете дуоденальных желез эмбрионов гибридных и домашних свиней, но отсутствовали у таковых дикого кабана. Содержание гликогена в энтероцитах и мышечной оболочке двенадцатиперстной кишки эмбрионов гибридных, диких и домашних животных составляло соответственно 199,5; 400,0 и 522,4 мг%, что, видимо, свидетельствует о более интенсивной дифференциации слизистой оболочки у первых.
У основания ворсинок слизистой оболочки двенадцатиперстной кишки 60-суточных эмбрионов гибридных, диких и домашних свиней формировались крипты и мышечный слой этой оболочки, который был нечетко выражен из-за протоков дуоденальных желез, расположенных в подслизистом слое; количество гликогена составляло соответственно 291,5; 254,0 и 344,0 мг%. В каемке энтероцитов помимо гликопротеинов была выявлена щелочная фосфатаза, в бокаловидных клетках — липаза. У эмбрионов гибридов и кабана энтероциты на верхушках ворсинок, помимо полостей в базальной части, утрачивали каемку и образовывали вакуоли в апикальной части, чего не наблюдалось у свиней породы ландрас. Наличие полостей и вакуолизация энтероцитов на верхушках ворсинок свидетельствуют об участии этих структур в процессах обмена веществ.
В позднеплодном периоде у гибридов продолжалось развитие ворсинок крипт и дуоденальных желез, которые к возрасту 80-90 сут разветвлялись и заполняли подслизистый слой, открываясь протоками в крипты. В последних увеличивалось количество бокаловидных клеток, в которых не отмечено содержания гликогена. Узкие и длинные ворсинки были покрыты каемчатым эпителием и бокаловидными клетками, на верхушках таких ворсинок энтероциты образовывали вакуоли и полости. Боковые стороны отдельных ворсинок приобрели фестончатое строение, что увеличивало площадь поверхности (у исходных форм фестончатость отсутствовала).
Слизистая оболочка двенадцатиперстной кишки эмбрионов гибридов и кабана (в возрасте 100 сут) и домашних свиней (80-90 сут) приобретала складчатость, что сопровождалось снижением содержания гликогена — соответственно до 91,7; 154,1 и 125,3 мг%.
При морфометрической оценке оболочек двенадцатиперстной кишки эмбрионов было выявлено, что к началу плодного периода (40 сут) толщина слизистой и мышечной оболочек у гибридов была достоверно меньше (Р < 0,05), чем у домашних свиней породы ландрас, и незначительно отличалась (Р = 0,05) от таковой у диких кабанов (табл. 1). К концу раннеплодного периода (60 сут) толщина слизистой оболочки у эмбрионов гибридов была больше, чем у диких кабанов, и почти такой же, как у домашних свиней. Толщина мышечной оболочки только одного гибридного эмбриона превосходила таковую исходных форм; к 70 сут эти показатели выравнивались.
Толщина мышечной оболочки у плодов гибридов в позднеплодный период
(80-100 сут) находилась в пределах индивидуальной изменчивости, слизистой (в возрасте 80 сут) — была достоверно меньше (Р < 0,05), чем у исходных форм, и приближалась к показателям домашних свиней в возрасте 90 и 100 сут (см. табл. 1). Ко времени рождения толщина слизистой оболочки гибридных поросят семиреченской породы оказалась достоверно больше (Р < 0,05), чем диких кабанов, и незначительно больше, чем домашних свиней. Относительная толщина оболочки двенадцатиперстной кишки существенно варьировала в зависимости от возраста эмбрионов гибридов (рис. 1, а). При этом у эмбрионов гибридов и кабанов этот показатель варьировал в большей степени, чем у домашних свиней, и ко времени рождения составлял соответственно 79,9; 79,0 и 81,8 % .
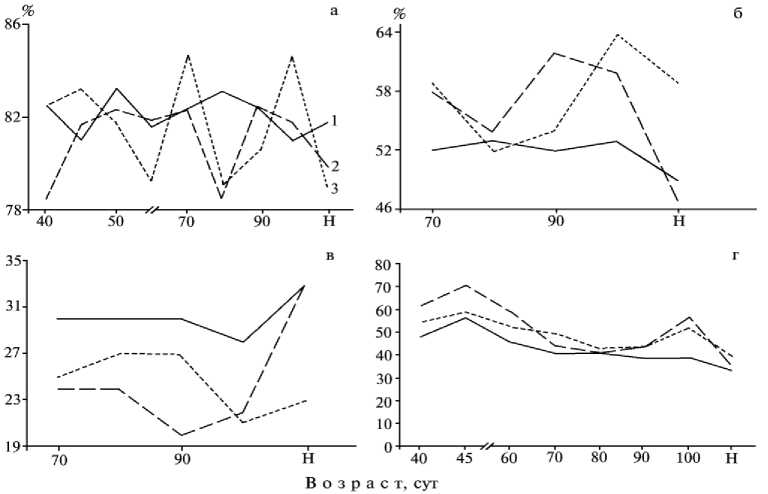
Рис. 1. Относительная толщина слизистой оболочки (а) , слизистого (б) и подслизистого (в) слоев, относительная длина ворсинок (г) двенадцатиперстной кишки эмбрионов разного возраста домашних, гибридных и диких свиней, а также новорожденных поросят (Н) : 1 — порода ландрас, 2 — гибриды семиреченской породы, 3 — кабан европейского подвида.
-
1. Толщина слизистой и мышечной оболочек, длина ворсинок двенадцатиперстной кишки эмбрионов гибридных свиней семиреченской породы и исходных форм в плодный и послеутробный периоды развития (мкм)
Возраст эмбриона, сут
Оболочка, ворсинки кишки
Домашние свиньи породы ландрас
Гибридные свиньи семиреченской породы
Дикий кабан европейского подвида
n
Толщина оболочки, длина ворсинок
От-до
n
Толщина оболочки, длина ворсинок
От-до
n
Толщина оболочки, длина ворсинок
От-до
40
Слизистая
2
258,0
254,2-261,8
6
157,1 ± 41,4
102,0-205,5
4
231,6 ± 35,7
212,5-263,5
Мышечная
54,4
51,8-57,0
42,5 ± 7,4
32,4-51,0
48,9 ± 17,8
34,0-59,5
Ворсинки слизистой
150,9 ± 48,2
144,5-157,2
125,1 ± 48,4
64,3-185,4
151,0 ± 29,7
136,0-178,5
45
Слизистая
6
246,8 ± 23,0
221,8-275,4
7
204,0 ± 19,9
170,0-236,4
3
294,7 ± 66,3
263,5-314,5
Мышечная
57,9 ± 7,1
50,2-68,0
45,1 ± 5,6
32,3-51,0
59,5 ± 6,8
51,0-68,0
Ворсинки слизистой
170,4 ± 13,2
150,4-183,6
175,1 ± 21,3
155,5-226,8
206,8 ± 55,2
187,0-229,5
50
Слизистая
2
272,0
249,0-295,0
3
267,6 ± 80,0
236,3-298,5
3
280,3 ± 44,2
263,5-297,5
Мышечная
54,8
51,0-58,6
57,1 ± 17,9
49,3-63,1
62,3 ± 22,1
51,0-68,0
Ворсинки слизистой
171,7 ± 32,6
167,4-176,0
207,5 ± 46,2
185,4-221,0
198,3 ± 22,1
187,0-204,0
60
Слизистая
7
334,8 ± 44,6
274,6-423,3
2
373,1
369,7-376,6
5
300,9 ± 34,0
263,5-331,5
Мышечная
75,5 ± 6,1
63,8-84,2
83,9
63,7-104,1
78,2 ± 8,5
68,0-85,0
Ворсинки слизистой
187,5 ± 20,7
153,0-222,7
267,4
265,0-269,9
198,9 ± 34,0
170,0-238,0
70
Слизистая
5
370,4 ± 34,9
336,5-406,3
4
330,7 ± 48,3
300,5-369,6
1
425,0
–
Мышечная
78,9 ± 8,1
71,4-87,6
70,9 ± 15,1
60,1-81,7
76,5
–
Ворсинки слизистой
182,1 ± 17,0
167,4-201,4
174,7 ± 46,4
136,0-202,3
246,5
–
80
Слизистая
7
400,9 ± 32,4
354,4-462,4
6
323,7 ± 18,1
307,8-353,2
3
487,3 ± 30,6
433,5-535,5
Мышечная
82,8 ± 12,7
70,6-113,0
88,2 ± 6,8
80,7-97,7
127,5 ± 0
127,5-127,5
Ворсинки слизистой
201,0 ± 22,8
166,0-242,2
166,7 ± 19,7
141,1-190,4
263,5 ± 88,4
229,5-297,5
90
Слизистая
4
487,3 ± 98,7
417,4-558,4
3
407,7 ± 115,9
365,5-454,7
3
631,8 ± 66,3
612,0-663,0
Мышечная
104,3 ± 23,8
90,1-124,1
85,4 ± 27,4
77,5-98,6
153,0 ± 77,3
119,9-178,5
Ворсинки слизистой
227,8 ± 32,7
198,0-244,8
215,0 ± 87,8
181,0-248,6
342,8 ± 88,4
314,5-382,5
100
Слизистая
5
524,1 ± 89,2
442,0-620,5
4
480,2 ± 80,8
421,5-537,0
3
688,5 ± 187,8
637,5-782,0
Мышечная
122,6 ± 20,0
96,0-136,0
105,3 ± 24,5
86,0-121,1
124,7 ± 55,2
102,0-144,5
Ворсинки слизистой
248,0 ± 54,8
204,8-314,5
331,3 ± 66,6
295,8-391,0
422,2 ± 88,4
399,5-467,5
Новорожденные
Слизистая
9
863,7 ± 64,2
762,4-10083,8
6
1060,7 ± 125,6
927,6-1241,8
4
746,4 ± 149,5
612,0-825,6
Мышечная
194,8 ± 37,1
141,1-326,8
266,5 ± 117,3
176,3-469,6
201,1 ± 97,3
136,0-275,0
Ворсинки слизистой
343,5 ± 32,8
289,9-412,2
457,1 ± 118,9
333,2-612,7
364,4 ± 13,2
344,0-391,0
-
2. Толщина слизистого и подслизистого слоев двенадцатиперстной кишки эмбрионов гибридных свиней семиреченской породы и исходных форм в плодный и послеутробный периоды развития (мкм)
Возраст эмбриона, сут
Слой слизистой оболочки
Домашние свиньи породы ландрас
Гибридные свиньи семиреченской породы
Дикий кабан европейского подвида
n
Толщина слоя
1 От-до
n
1 Толщина слоя
От-до
n
1 Толщина слоя
От-до
70
Слизистый
5
235,3 ± 21,2
217,6-260,1
4
232,9 ± 10,7
225,2-240,6
1
297,5
–
Подслизистый
135,1 ± 21,3
112,9-155,6
98,5 ± 36,1
77,3-129,0
127,5
–
80
Слизистый
7
260,8 ± 18,1
232,0-292,4
6
225,4 ± 26,8
196,5-263,5
3
323,0 ± 121,5
280,5-374,0
Подслизистый
146,1 ± 14,7
122,4-171,7
98,3 ± 8,9
89,7-112,0
164,3 ± 33,1
153,0-178,5
90
Слизистый
4
312,0 ± 58,2
257,6-340,8
3
309,1 ± 113,7
273,7-361,2
3
422,1 ± 110,5
391,0-476,0
Подслизистый
175,3 ± 50,5
149,6-221,8
98,6 ± 24,3
91,8-110,5
209,7 ± 44,2
187,0-221,0
100
Слизистый
5
340,7 ± 55,7
292,4-403,8
4
351,5 ± 61,7
310,2-398,4
3
518,5 ± 121,5
484,5-578,0
Подслизистый
183,1 ± 35,3
146,2-216,8
128,7 ± 34,5
107,9-157,2
170,0 ± 77,3
144,5-204,0
Новорожденные
Слизистый
9
517,0 ± 30,6
437,8-590,9
6
623,9 ± 108,8
542,8-814,8
4
523,2 ± 64,1
476,0-567,6
Подслизистый
346,7 ± 47,4
255,8-493,0
436,8 ± 149,4
298,8-672,5
223,2 ± 85,4
136,0-258,0
-
4. Толщина слизистого слоя ободочной кишки эмбрионов гибридных свиней семиреченской породы и исходных форм в плодный и послеутробный периоды развития (мкм)
Возраст эмбриона, сут
Домашние свиньи породы ландрас
Гибридные свиньи семиреченской породы
Дикий кабан европейского подвида
n
1 Толщина оболочки
1 От-до
n
1 Толщина оболочки
От-до
n
Толщина оболочки
^ От-до
70
6
279,6 ± 93,0
177,6-394,4
6
228,6 ± 10,8
288,1-344,2
1
272,0
–
80
6
299,8 ± 55,0
236,3-364,6
4
302,5 ± 90,3
259,2-374,8
3
270,2 ± 109,5
224,0-306,0
90
5
238,8 ± 42,2
205,7-282,2
4
330,6 ± 120,2
232,0-385,9
3
289,0 ± 22,7
280,5-297,5
100
4
231,6 ± 85,7
168,3-278,0
5
280,4 ± 61,4
225,2-336,6
3
223,8 ± 34,1
212,5-238,0
Новорожденные
7
211,5 ± 12,5
195,5-230,4
6
243,0 ± 36,2
206,4-290,7
4
210,4 ± 19,9
195,5-221,0
В позднеплодный период при возникновении мышечного слоя слизистой оболочки увеличение толщины последней было связано с ростом слизистого и подслизистого слоев. Толщина слизистого слоя у эмбрионов гибридов и домашних свиней была приблизительно одинаковой и недостоверно меньше, чем у кабанов (Р > 0,05) (табл. 2). Ко времени рождения показатели у гибридов были выше, чем у исходных форм (без достоверных различий). Относительное утолщение слизистого слоя эмбрионов гибридов проходило более интенсивно, чем домашних свиней, одна- ко ко времени рождения относительная толщина этого слоя составляла соответственно 47,0 и 48,8 % (против 55,8 % у кабанов) (см. рис. 1, б).
В позднеплодный период толщина подслизистого слоя эмбрионов гибридов была меньше, чем исходных форм (разница достоверна в возрасте 80-90 сут) (см. табл. 2). Ко времени рождения различия между показателями нивелировались. Увеличение относительной толщины этого слоя проходило менее интенсивно у эмбрионов гибридов по сравнению с домашними свиньями и диким кабаном, однако ко времени рождения эти показатели составляли соответственно 32,9; 32,7 и 23,2 % (см. рис. 1, в).
Утолщение слизистой оболочки двенадцатиперстной кишки обусловлено развитием ворсинок. При значительной индивидуальной изменчивости длина ворсинок у эмбрионов гибридов в возрасте 40 сут была меньше, 45 и 50 сут — незначительно больше (Р > 0,05), 60 сут — достоверно больше (Р < 0,05), чем у исходных форм (см. табл. 1). В раннеплодный период относительная длина ворсинок слизистой двенадцатиперстной кишки у гибридов была больше, чем у исходных форм (см. рис. 1, г). В позднеплодный период (70-90 сут) длина ворсинок находилась в пределах индивидуальной изменчивости сравниваемых особей и у эмбрионов гибридов к возрасту 100 сут занимала промежуточное положение между наибольшими показателями кабанов и наименьшими свиней породы ландрас (см. табл. 1). Ко времени рождения средняя длина ворсинок у гибридов превосходила таковую у исходных форм (Р > 0,05).
Относительная длина ворсинок в этот период у всех эмбрионов была практи- чески одинаковой (см. рис. 1, г). Только у 100-суточных плодов гибридов и кабана интенсивность роста ворсинок была выше, чем у домашних свиней. Ко времени рож- дения относительная длина ворсинок у эмбрионов гибридных, домашних и диких свиней составляла соответственно 34,2; 32,8 и 39,4 %.
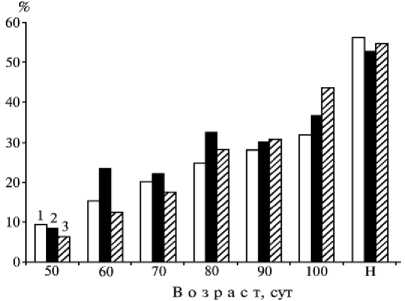
Рис. 2. Плотность расположения дуоденальных желез у эмбрионов разного возраста и новорожденных ( Н) свиней породы ландрас (1) , гибридов семиречен-ской породы (2) и кабана европейского подвида (3).
В подслизистом слое находятся дуоденальные железы, плотность расположения которых при закладке в возрасте 50 сут у плодов гибридов и домашних свиней была близкой (соответственно 8,9 и 9,7 %), а у кабанов — наименьшей (6,5 %). В 60-80-суточном возрасте этот показатель у эмбрионов гибридов увеличивался более интенсивно, чем у таковых домашних свиней и дикого кабана, а в 60 сут составлял соответственно 23,3; 15,5 и 12,5 % (Р < 0,05) (рис. 2). К возрасту 90 сут различия сглаживались, причем гибриды занимали промежуточное положение. При рождении у гибридных, диких и домаш- них поросят плотность расположения дуоденальных желез составляла соответственно 52,7; 54,6 и 56,1 %.
П о д в з д о ш н а я к и ш к а. Закладка ворсинок в подвздошной кишке происходила позже, чем в двенадцатиперстной, эпителиальные бугорки и первичная складчатость появлялись к 45 сут, истинные ворсинки — к 50 сут, единичные бока- ловидные клетки и крипты — к 60 сут эмбрионального развития; содержание гликогена, гликопротеинов и ферментов было аналогичным таковому в двенадцатиперстной кишке.
Верхушки ворсинок подвздошной кишки эмбрионов гибридов и дикого кабана были сильно вакуолизированы и имели обширные полости между энтероцитами и соединительной тканью; у первых выявлены также ворсинки, имеющие фестончатое строение боковых поверхностей. У эмбрионов домашних свиней наблюдалась в основном вакуолизация верхушек ворсинок. Обменные и пищеварительные процессы в кишечнике эмбрионов гибридов и дикого кабана проходили более активно, чем у домашних свиней: масса содержимого тонкого кишечника в возрасте 70 сут составляла соответственно 1,9; 1,5 и 1,0 г; 75-80 сут — 3,4; 2,7 и 2,6 г; к концу утробного развития — 8,9; 9,3 и 7,4 г.
В позднеплодный период у эмбрионов гибридов образовывались лимфатические фолликулы (не по всему периметру кишки), которые к 90- и 100-суточному возрасту приобретали соответственно шаровидную и грушевидную форму и располагались в подслизистом и слизистом слоях. Ворсинки укорачивались в местах расположения фолликулов, при отсутствии последних были длинными и частично фестончатыми, чего не наблюдалось у диких и домашних свиней.
Двенадцатиперстная и подвздошная кишки новорожденных поросят при наличии всех элементов строения взрослых свиней утрачивали эмбриональное строение ворсинок тонкого кишечника. Прекращение у новорожденных плацентарного кровообращения и снижение доли участия кишечника в межуточном обмене веществ приводило к исчезновению вакуолей и полостей в энтероцитах на верхушках ворсинок, энтероциты приобретали каемку и базальную мембрану, в которых содержались гликопротеины и ферменты, принимающие участие в защитных функциях. Такая структурная перестройка ворсинок, обусловленная созреванием, была выражена в наибольшей степени у новорожденных диких свиней; у гибридов верхушки некоторых ворсинок в подвздошной кишке сохраняли эмбриональное строение, то есть были менее зрелыми. Об этом же свидетельствует и количество гликогена в подвздошной кишке, которое уменьшалось вследствие перестройки ворсинок ко времени рождения у диких кабанов, домашних и гибридных свиней соответственно до 77,0; 122,0 и 199,5 мг%.
О б о д о ч н а я к и ш к а. Эмбриогенез толстого кишечника оценивали по развитию ободочной кишки, в которой, как и в тонком кишечнике, формируются ворсинки. У 40-суточных эмбрионов гибридов и диких кабанов внутренняя поверхность ободочной кишки была представлена эпителиальными бугорками и первичными складками, из которых образуются ворсинки; у таковых домашних свиней эта поверхность выстлана только многорядным эпителием. Посредством преобразования эпителиальных бугорков и первичных складок к 45-50-суточному возрасту формировались первичные ворсинки, покрытые многорядным эпителием, содержание гликогена в котором у эмбрионов домашних свиней и гибридов составляло соответственно 270,0 и 433,0 мг%. К возрасту 60 сут обра-
|
ОО о |
3. Толщина слизистой и мышечной оболочек, длина ворсинок ободочной кишки эмбрионов гибридных свиней семиреченской породы и исходных форм в плодный и послеутробный периоды развития (мкм) |
|
Возраст эм- Оболочка, ворсин- Домашние свиньи породы ландрас Гибридные свиньи семиреченской породы Дикий кабан европейского подвида |
|
|
бриона, сут ки кишки n Толщина оболочки, длина От-до n Толщина оболочки, длина От-до n Толщина оболочки, От-до |
|
|
ворсинок ворсинок длина ворсинок |
|
|
40 |
Слизистая 2 98,2 85,8-110,5 3 140,8 ± 63,6 110,5-158,4 5 139,4 ± 37,5 110,5-178,5 |
|
Мышечная 29,8 27,2-32,3 31,4 ± 7,9 28,9-34,8 32,3 ± 9,4 25,5-42,5 |
|
|
Ворсинки слизистой – – 81,3 ± 56,8 56,1-98,6 97,8 ± 18,8 76,5-110,5 |
|
|
45 |
Слизистая 4 158,9 ± 66,4 127,5-212,5 6 141,3 ± 8,0 131,7-150,4 6 167,2 ± 18,2 153,0-195,5 |
|
Мышечная 37,6 ± 24,5 23,8-55,2 25,4 ± 2,2 23,8-28,9 38,2 ± 3,6 34,0-42,5 |
|
|
Ворсинки слизистой 114,8 ± 26,6 98,6-132,6 86,5 ± 7,3 76,5-93,5 117,6 ± 18,2 102,0-144,5 |
|
|
50 |
Слизистая 3 190,4 ± 111,1 162,4-245,6 2 264,3 243,9-284,7 2 165,8 161,5-170,0 |
|
Мышечная 42,2 ± 9,0 38,2-45,0 31,0 29,7-32,3 34,0 34,0-34,0 |
|
|
Ворсинки слизистой 138,6 ± 68,1 119,8-170,8 168,2 145,3-191,2 127,5 127,5-127,5 |
|
|
60 |
Слизистая 7 261,6 ± 36,1 214,2-315,4 3 388,1 ± 159,0 319,6-438,6 5 275,4 ± 28,1 246,5-297,5 |
|
Мышечная 59,1 ± 10,0 45,0-73,1 44,l ± l2,4 39,1-48,4 62,9 ± l4,0 51,0-76,5 |
|
|
Ворсинки слизистой 172,6 ± 21,8 137,7-198,9 239,6 ± 93,1 204,8-274,5 218,4 ± 46,8 187,0-272,0 |
|
|
70 |
Слизистая 6 370,9 ± 117,8 258,4-533,0 6 370,1 ± 54,2 305,9-423,3 1 323,0 – |
|
Мышечная 85,0 ± 47,7 52,7-164,0 46,4 ± 8,8 34,8-55,5 59,5 – |
|
|
Ворсинки слизистой 207,4 ± 70,7 138,6-303,4 214,6 ± 75,3 148,7-312,8 229,5 – |
|
|
80 |
Слизистая 6 375,0 ± 64,2 325,6-475,2 4 379,4 ± 115,5 309,3-457,2 3 355,2 ± 64,1 326,0-374,0 |
|
Мышечная 85,0 ± 18,3 68,8-111,4 62,8 ± l3,3 53,5-70,5 107,7 ± 90,8 85,0-153,0 |
|
|
Ворсинки слизистой 204,8 ± 53,6 151,3-276,2 230,5 ± 124,1 179,3-338,3 184,2 ± 79,5 153,0-212,5 |
|
|
90 |
Слизистая 5 330,8 ± 83,4 266,9-418,2 4 410,7 ± 139,4 300,0-478,5 3 371,2 ± 34,1 357,0-382,5 |
|
Мышечная 99,6 ± 24,8 79,0-124,1 7I,6 ± 33,2 47,6-90,1 110,5 ± 22,7 102,0-119,0 |
|
|
Ворсинки слизистой 149,2 ± 42,7 99,4-176,8 246,2 ± 57,7 202,3-276,2 164,3 ± 22,7 153,0-170,0 |
|
|
100 |
Слизистая 4 302,8 ± 102,2 219,3-350,2 5 374,9 ± 43,6 332,3-411,4 3 272,0 ± 56,8 255,0-297,5 |
|
Мышечная 95,2 ± 35,8 70,6-116,4 87,7 ± 16,4 73,1-102,8 116,2 ± 79,5 93,5-153,0 |
|
|
Ворсинки слизистой 136,0 ± 54,5 106,2-176,0 147,7 ± 53,9 102,8-200,6 80,8 ± 8,8 76,5-85,0 |
|
|
Новорожденные Слизистая 4 298,4 ± 26,7 255,0-329,8 6 34I,l ± 78,9 263,2-447,2 4 276,3 ± 39,8 246,5-297,5 |
|
|
Мышечная 152,7 ± 24,3 127,5-195,5 165,l ± 20,0 139,3-185,8 155,1 ± 26,5 136,0-170,0 |
|
зовывались истинные ворсинки, покрытые цилиндрическими энтероцитами, расположенными однорядно, среди которых располагались бокаловидные клетки; содержание гликогена составляло 622,6; 866,0 и 498,2 мг% (соответственно свиньи породы ландрас, дикие кабаны и гибриды), что обусловлено разной степенью дифференциации структурных элементов слизистой оболочки эмбрионов. В связи с этим можно предположить, что интенсивность формообразовательных процессов у эмбрионов гибридов семиреченской породы была выше, чем у исходных форм.
К возрасту 70 сут слизистая оболочка ободочной кишки была разделена на слизистый и подслизистый слои мышечным слоем, содержащим гликоген. Слизистый слой был представлен криптами и ворсинками; у эмбрионов гибридов отмечено образование ворсинок, боковые поверхности которых имели фестончатое строение, что увеличивало площадь поверхности. Энтероциты на верхушках ворсинок у эмбрионов дикого кабана, домашних свиней и гибридов были вакуолизированы и содержали полости между эпителием и соединительной тканью, что свидетельствует об усилении обменных процессов в толстом кишечнике, масса содержимого которого к 70-суточному возрасту составляла соответственно 0,37; 0,30 и 0,18 г (у гибридов к 75 сут увеличивалась до 0,93 г), а к концу утробного развития — соответственно 4,0; 3,3 и 3,1 г.
У эмбрионов в возрасте 80 сут ворсинки слизистого слоя ободочной кишки укорачивались и утолщались, постепенно преобразуясь в крипты. В апикальной части ворсинок уменьшалась вакуолизация энтероцитов, исчезали полости у основания последних, снижалось содержание гликогена, которое у 100-суточных эмбрионов домашних свиней, гибридов и кабанов составляло соответственно 134,1; 162,3 и 177,0 мг%.
К возрасту 90 сут появлялась складчатость слизистой оболочки, в слизистом слое которой формировались шаровидные лимфатические фолликулы, заполняющие к возрасту 100 сут также подслизистый слой. У новорожденных поросят слизистая оболочка ободочной кишки приобретала складчатость и была представлена криптами, однако между складками встречались септы, похожие по конфигурации и вакуолизации энтероцитов на ворсинки, не трансформированные в крипты. Содержание гликогена у новорожденных свиней породы ландрас, гибридов и кабанов составляло соответственно 117,0; 135,8 и 43,0 мг%.
При морфометрической оценке оболочек ободочной кишки показано, что толщина слизистой оболочки 40-суточных эмбрионов гибридов была больше, чем домашних свиней (Р > 0,05), и почти такой же, как кабанов (табл. 3). Дальнейшее утолщение слизистой оболочки у гибридов к возрасту 45 сут было меньшим, а к концу раннеплодного периода (50-60 сут) — б о льшим, чем у исходных форм (при значительных индивидуальных колебаниях). Толщина слизистой оболочки эмбрионов гибридов в возрасте 90 и 100 сут (Р = 0,05) и при рождении была больше, чем у исходных форм (при значительной индивидуальной изменчивости). По относительной толщине слизистой оболочки ободочной кишки эмбрионов гибриды превосходили исходные формы в б о льшей степени в раннеплодный и в меньшей — в позднеплодный период (рис. 3, а). Этот показатель у новорожденных домашних, гибридных и диких поросят составлял соответственно 66,2; 67,3 и 64,0 %.
Толщина мышечной оболочки ободочной кишки у эмбрионов гибридов увеличивалась медленнее и была меньше, чем у исходных форм. При этом различия были достоверными (Р < 0,05) между эмбрионами гибридов и кабанов в возрасте 45, 50 и 80 сут (см. табл. 3). Относительная толщина этой оболочки в период плодного развития у гибридов также была меньше, чем у исходных форм. У новорожденных поросят породы ландрас, гибридов семиреченской породы и диких кабанов этот показатель составлял соответственно 33,8; 32,7 и 36,0 % (см. рис. 3, б).
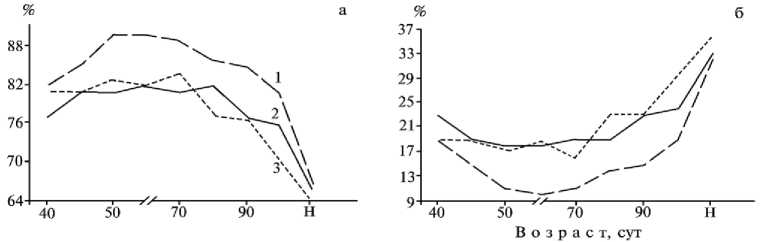
Рис. 3. Относительная толщина слизистой (а) и мышечной (б) оболочек, слизистого слоя (в) ободочной кишки эмбрионов разного возраста домашних, гибридных и диких свиней, а также новорожденных поросят (Н) : 1 — гибриды семи-реченской породы, 2 — порода ландрас, 3 — кабан европейского подвида.
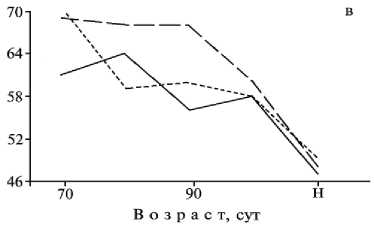
Увеличение толщины слизистой оболочки в позднеплодный период в основном обусловлено приростом слизистого слоя, толщина которого в среднем была наибольшей у гибридов (при значительной индивидуальной изменчивости) (табл. 4). Относительная толщина этого слоя у эмбрионов гибридов в возрасте 70, 80 и 90 сут также была больше, чем у исходных форм (см. рис. 3, в). Ко времени рождения интенсивность прироста слизистого слоя у гибридов снижалась и составляла 48,0 %, то есть была промежуточной между таковой у домашних свиней и диких кабанов (соответственно 46,9 и 48,9 %).
Увеличение толщины слизистой оболочки и слизистого слоя обусловлено трансформацией ворсинок в крипты. Длина ворсинок у эмбрионов гибридов и кабанов в возрасте 40 сут варьировала от 56,1 до 110,5 мкм (см. табл. 3). Дальнейшее удлинение первичных складок и первичных ворсинок у гибридов находилось в пределах индивидуальной изменчивости сравниваемых особей (в возрасте 45 сут меньше, 50 сут — больше, чем у исходных форм). К концу раннеплодного периода длина истинных ворсинок эмбрионов гибридов была больше, чем свиней породы ландрас и кабанов (Р = 0,05). Это преимущество сохранялось и в позднеплодный период, что свидетельствует о замедлении трансформации слизистой оболочки у гибридов. Свободные верхушки бывших ворсинок и септ, напоминающих по конфигурации ворсинки, у новорожденных гибридных поросят обеспечивают б о льшую, чем у исходных форм, продолжительность и активность обменных процессов в толстом кишечнике.
Таким образом, при оценке возрастных изменений морфометрических показателей структурных элементов кишечника у эмбрионов гибридных свиней семире-ченской породы наряду с промежуточным типом наследования выявлена значительная индивидуальная изменчивость по толщине оболочек и слоев, размеру ворсинок и крипт, а также плотности расположения дуоденальных желез. Более высокая активность обменных и пищеварительных процессов у гибридов по сравнению с дикими кабанами европейского подвида и домашними свиньями породы ландрас обусловлена образованием вакуолей и полостей в энтероцитах на верхушках ворсинок, фестон-чатостью строения боковых поверхностей последних, значительной продолжительностью функционирования ворсинок толстого кишечника, а также интенсивным формированием дуоденальных желез. При этом в секрете желез у поросят гибридов семиреченской породы и породы ландрас наряду с нейтральными присутствуют и кислые гликопротеины, не обнаруженные у дикого кабана. Активный формообразовательный процесс в кишечнике эмбрионов гибридов сопровождается значительным уменьшением содержания гликогена в тканях, а также большим количеством содержимого в тонком и толстом кишечнике. Следовательно, кишечно-желудочный тип пищеварения, формирующийся в процессе утробного развития, у гибридных животных выражен в большей степени, чем у исходных форм. При этом у новорожденных гибридных и домашних поросят частично сохраняется эмбриональное строение ворсинок, участвующих в обменных процессах, что ослабляет защитные механизмы при функционировании кишечника. Это необходимо учитывать при кормлении и содержании новорожденных животных.
Л И Т Е Р А Т У Р А
Институт проблем экологии и эволюции им. А.Н. Северцова, 119071, Москва, Ленинский просп., 33
