К вопросу о маркировании локусов Pl, контролирующих устойчивость подсолнечника к возбудителю ложной мучнистой росы

Автор: Рамазанова С.А., Антонова Т.С.
Рубрика: Селекция и семеноводство сельскохозяйственных растений
Статья в выпуске: 1 (177), 2019 года.
Бесплатный доступ
Одной из самых вредоносных болезней подсолнечника является ложная мучнистая роса. К настоящему времени в мире обнаружено более 45 физиологических рас патогена. Эффективным методом контроля над возбудителем болезни является введение доминантных генов устойчивости к нему в растение-хозяина. Современные технологии генотипирования с помощью ДНК-маркеров позволяют контролировать наличие этих генов на каждом этапе селекции. Апробированы известные из литературных источников девять STS и три SSR-маркера генов Pl5 , Pl6 и Pl8 , контролирующих устойчивость к расам P. halstedii , для идентификации этих генов у линий-дифференциаторов устойчивости подсолнечника, входящих в международный тест-набор для идентификации рас P. halstedii . Был отобран и апробирован молекулярный STS-маркер НАР3, пригодный для идентификации локуса Pl6 , контролирующего устойчивость подсолнечника к возбудителю ложной мучнистой росы.
Днк-маркеры, мас, r-гены, устойчивость, подсолнечник
Короткий адрес: https://sciup.org/142220356
IDR: 142220356 | УДК: 631.523:633.854.78 | DOI: 10.25230/2412-608X-2019-1-177-17-23
To a question about marking of Pl loci controlling sunflower resistance to downy mildew pathogen
One of the most dangerous diseases on sunflower is downy mildew. By now, there are known more than 45 physiological races of the pathogen in the world. The effective method to control the pathogen is to introduce dominant gens of resistance into host-plant. The modern technologies of genetic typing using DNA-markers allowing controlling these genes present at the each stage of breeding process. Our research was conducted in the All-Russia Research Institute of Oil Crops by Pustovoit V.S., Krasnodar, at the laboratory of immunity and molecular marking. We approbated nine STS and three SSR-markers of the gens Pl5 , Pl6 and Pl8 well known from the literary sources which control sunflower resistance to P. halstedii races with purpose to identify these genes in lines-differentiators of sunflower resistance which are included into the international test-set for P. halstedii races identification. We selected and test a molecular STS-marker НАР3 which can be sued for identification of Pl6 locus controlling sunflower resistance to downy mildew pathogen.
Текст научной статьи К вопросу о маркировании локусов Pl, контролирующих устойчивость подсолнечника к возбудителю ложной мучнистой росы
Введение. Болезнь подсолнечника – ложная мучнистая роса, вызываемая облигатным паразитом – оомицетом Plasmopara halstedii (Farl.) Berl. et de Toni, является одной из самых вредоносных. При благоприятных условиях для развития возбудителя, болезнь вызывает значительное сокращение урожая семян подсолнечника и содержания в них масла [1]. К настоящему времени в мире обнаружено более 45 физиологических рас патогена и наблюдается постоянное возникновение новых, более вирулентных. Так, если до 2007 г. в Европе насчитывалось 13 рас патогена, в Америке – 20, в России в Краснодарском крае – 7, то уже к 2015 г. их число возросло до 24 в Европе и до 40 в Америке [1; 2]. В Краснодарском крае за последние 5 лет идентифицированы три новых расы: 334, 713 и 733 [4].
Наиболее эффективным методом контроля над возбудителем болезни является введение доминантных генов устойчивости к нему в растение-хозяина. Устойчивость к P. halstedii контролируется генами Pl. Впервые они были идентифицированы в культурном подсолнечнике еще в 70-х годах прошлого столетия, обозначены как Pl 1 и Pl 2 , контролировали устойчивость к расам 100 и 300 соответственно [5; 6]. Позднее они были картированы как относящиеся к восьмой группе сцепления LG на генетической карте SSR-локусов (Simple Sequence Repeat) [7]. Гены Pl 3 и Pl 4 контролируют устойчивость к этим же расам, локализация их на генетической карте подсолнечника пока не известна [8]. Гены Pl 5 и Pl 8 контролируют устойчивость к 16-и расам P. halstedii [9]. Эти два гена сцеплены и относятся к тринадцатой группе сцепления на генетической карте SSR-локусов. Ген Pl 6 так же, как Pl 1 и Pl 2 , расположен в восьмой группе сцепления LG и контролирует устойчивость к расам 100, 300, 700, 703, 710, 330, 770 и 730. К этой же группе сцепления относятся и кластеры генов Pl 7 , Pl 9 , Pl 10 [10]. Анализ публикаций, посвященных картированию генов устойчивости подсолнечника к P. halstedii , показал, что тип их расположения является одиночным диаллельным, то есть один локус имеет множество кодоминантных аллелей, контролирующих устойчивость к разным расам патогена [8; 9; 13; 16; 28]. Сгруппированы они, как минимум, в четырех геномных комплексах в трех группах сцепления (LG) [16].
Цель исследования – поиск ДНК-маркеров для идентификации генов, контролирующих устойчивость к P. halstedii , в селекционном материале ВНИИМК и создание на их основе системы маркеров с последующим использованием ее в селекционных программах, направленных на создание генотипов подсолнечника с комплексной устойчивостью к разным расам возбудителя ложной мучнистой росы.
Материалы и методы Объектом исследования послужили 14 линий-дифференциаторов устойчивости подсолнечника, входящих в международный тест-набор для идентификации рас P. halstedii (таблица).
Таблица 1
Линии-дифференциаторы устойчивости подсолнечника к поражению ложной мучнистой росой, содержащие известные гены Pl
|
Линия-дифференциатор |
Pl -ген |
Группа сцепления |
Литературный источник |
|
RHA-419 |
Pl arg |
LG1 |
Dussle et al., 2004; Wieckhorst et al., 2010; Imerovski et al., 2014 |
|
XRQ |
Pl 5 |
LG13 |
Bert et al., 2001; Radwan et al., 2004 |
|
83HR4RM |
Pl 7 |
LG8 |
Jocić et al., 2012 |
|
HIR-34 |
Pl 4 |
Не определена |
Jocić et al., 2012 |
|
PSC8 |
Pl 2 |
LG8 |
Gascuel et al., 2014 |
|
YVQ |
Pl 8 |
LG13 |
Gascuel et al., 2014 |
|
HA-335 |
Pl 6 |
LG8 |
Bouzidi et al., 2002; Panković et al., 2007 |
|
HA-R5 |
Pl 13 |
LG1 |
Mulpuri et al., 2009; Bertero de Romano et al., 2010 |
|
HA-R4 |
Pl 16 |
LG1 |
Liu et al., 2012 |
|
803-1 |
Pl 5 |
LG13 |
Gascuel et al., 2014 |
|
PM-17 |
Pl 5 |
LG13 |
Gascuel et al., 2014 |
|
DM-2 |
Pl 5, Pl 11, Pl 12 |
LG13 |
Rahim et al., 2002 |
|
RHA-274 |
Pl 2 , Pl 9, Pl 10 |
LG8 |
Gulya et al., 1991; Rahim et al., 2002; Molinero-Ruiz et al., 2002; Romano et al., 2010 |
|
RHA-265 |
Pl 1 |
LG8 |
Gedil et al., 2001; Rahim M., 2002 |
Для выделения ДНК применяли метод, основанный на использовании лизирующего буфера, содержащего гексадеци-лтриметиламмоний бромид (СТАВ) с модификациями [11; 12]. Концентрацию ДНК в полученном препарате определяли визуально по интенсивности свечения пробы объемом 10 мкл в ультрафиолетовом свете в 1 %-ном агарозном геле с добавлением 2 мкл бромистого этидия. Электрофорез растворов ДНК проводили при напряжении 100 V в течение 30 мин в трис-ацетатном буферном растворе (ТАЕ).
Для ПЦР-анализа использовали 12 праймеров (HAP1, HAP2 HAP3, Ha-P1, Ha-P2, Ha-P3, Ha-P4, Ha-P5, Ha-P6, ORS 37, ORS 166 и ORS 1043), разработанных для маркирования локусов Pl 5 Pl 6 Pl 8 [9; 13; 14; 15]. Полимеразную цепную реакцию выполняли в реакционной смеси (25 мкл) следующего состава: 67 мМ Трис-HCl (рН 8,8); 16,6 мM сульфата аммония; 1,5–3,0 мM MgCl 2 ; 0,01 % Tween 20; по 0,2
мM дезоксирибонуклеозидфосфатов; по 10 пМ праймеров; 10 нг матричной ДНК и 1 ед. рекомбинантной термостабильной ДНК-полимеразы (НПО «СибЭнзим», Россия). Реакции проводили в термоциклере S1000тм (BioRad, США) при следующих температурных режимах: начальная денатурация при 95 °С в течение 3 мин, далее 35 циклов с последовательной сменой температур: денатурация при 94 ºС в течение 10 сек, отжиг праймера при 58–63 ºС (в зависимости от праймера) – 30 сек, элонгация при 72 ºС – 1 мин 30 сек и заключительная элонгация 10 мин.
Электрофорез продуктов амплификации проводили в геле, содержащем 2 % агарозы и SВ-буфер, с использованием камеры для горизонтального электрофореза SE-2 (Хеликон, Россия) при напряжении ≈ 200 В, силе тока ≈ 100 мА, в течение 30 мин. Гели окрашивали бромистым этидием. Для визуализации и документирования результатов электрофореза применяли систему цифровой документации видеоизображения BIO-PRINT (Vil-ber Lourmat, Франция).
Результаты и обсуждение. По сообщениям многих авторов, каждая линия-дифференциатор устойчивости из международного тест-набора для идентификации рас P. halstedii содержит один или несколько генов Pl, которые обеспечивают их устойчивость к определенным расам патогена. В таблице представлены 14 линий подсолнечника, соответствующие им гены и группы сцепления, в которых они картированы на генетической карте SSR-локусов.
Для поиска ДНК-маркеров генов устойчивости Pl5, Pl6 и Pl8 к P. halstedii исследовали молекулярно-генетический полиморфизм девяти STS (Sequence-Tagged Site) и трех SSR (Simple Sequence Repeat) локусов ДНК. Результаты ПЦР ДНК линий подсолнечника показали, что только с парой праймеров HAP1 не было получено амплифицированных фрагмен- тов. По остальным 11 локусам были подобраны оптимальные параметры реакции амплификации и получен отжиг соответствующих праймеров на матрице. По четырем локусам (ORS 166, Ha-P3, Ha-P5 и Ha-P6) не было выявлено полиморфных фракций ДНК, поэтому эти маркеры были исключены из дальнейшего исследования.
По пяти локусам (ORS 37, ORS 1043, Ha-P1, Ha-P2 и Ha-P4) выявлен полиморфизм в виде наличия–отсутствия фрагментов ДНК. Для примера на рисунках 1 и 2 представлены фореграммы продуктов амплификации ДНК с праймерами ORS 37 и ORS 1043. В результате ПЦР с праймерами, фланкирующими эти микроса-теллитные локусы, не было получено фрагментов ДНК, характерных для линии НA-335, в генотипе которой присутствует ген Pl 6 ; так же, как и для линий XRQ, YVQ, и 803-1, PM-17, DM-2 – носителей кластеров генов Pl 5 /Pl 8.
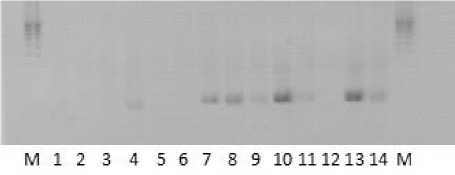
Рисунок 1 – Фореграмма продуктов амплификации ДНК линий-дифференциаторов подсолнечника с праймером ORS 1043. Дорожки:1 – RHA-419; 2 – XRQ;
3 – 83HR4RM; 4 – HIR 34; 5 – PSC8; 6 – YVQ; 7 – HA335; 8 – HAR-5; 9 – HAR-4; 10 – 803-1;
11 – PM-17; 12 – DM-2; 13 – RHA-274;
14 – RHA-265; M – маркер молекулярного веса

1 2 3 4 5 6 7 8 9 10 11 12 13 14 К" М
Рисунок 2 – Фореграмма продуктов амплификации ДНК линий-дифференциаторов подсолнечника с праймером ORS 37.
Дорожки: 1 – RHA-419; 2 – XRQ;
3 – 83HR4RM; 4 – HIR 34; 5 – PSC8; 6 – YVQ; 7 – Ha335; 8 – HAR-5; 9 – HAR-4; 10 – 803-1;
11 – PM-17; 12 – DM-2; 13 – RHA-274;
14 – RHA-265; M – маркер молекулярного веса
STS-локус HAP3 – один из трех, разработанных для маркирования локуса Pl 6 . Он представляет собой кластер из 13 генов, близко расположенных друг к другу, относящихся к TIR-NBS-LRR классу R-генов и картированных в группе сцепления LG8 на генетической карте SSR [13]. По данным авторов, эта праймерная пара дает четыре полиморфных фрагмента ДНК длиною от 988 до 1811 пар нуклеотидов (п.н.). Из них фракции длиною 988, 1119 и 1811 пар нуклеотидов выявлены у линий подсолнечника, устойчивых к расам 100, 300, 700, 703 и 710, а фрагмент длиною 1406 п. н. – у восприимчивого к этим расам образца. В нашем исследовании у линии НА-335 по локусу НАР3 получено два фрагмента ДНК в указанном диапазоне длин (рис. 3, дорожка 7). У линии РМ-17 (рис. 3, дорожка 8) не выявлено амплифицированных фрагментов ДНК, что может свидетельствовать об отсутствии у неё локуса Pl 6 . У остальных изученных двенадцати линий получены фрагменты ДНК разных длин, отличающиеся от таковых у линии НА-335 (рис. 3).
Был изучен молекулярно-генетический полиморфизм локуса НАР3 у линий подсолнечника селекции ВНИИМК с целью маркирования кластера генов Pl 6 . Для исследования выбраны линии, отличающиеся по устойчивости к разным расам.
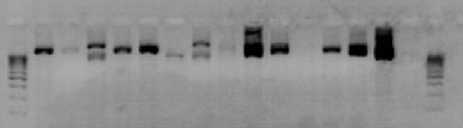
М 1 2 3 4 5 6 7 8 9 10 1112 13 14 К" М
Рисунок 3 – Фореграмма продуктов амплификации ДНК линий-дифференциаторов подсолнечника с праймером НАР3. Дорожки: 1 – RHA-419; 2 – XRQ;
3 – 83HR4RM; 4 – HIR 34; 5 – PSC8; 6 – YVQ; 7 – HА335; 8 – HAR-5; 9 – HAR-4; 10 – 803-1;
11 – PM-17; 12 – DM-2; 13 – RHA-274;14 – RHA-265; M – маркер молекулярного веса
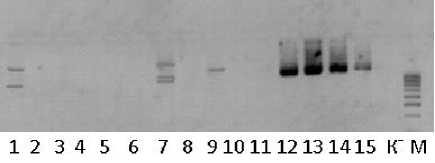
Рисунок 4 – Фореграмма продуктов амплификации ДНК линий подсолнечника селекции ВНИИМК с праймером НАР3.
Дорожки: 1 – НА-335; 2 – Л2018-1; 3 – Л689-15; 4 – Л693-15; 5 – Л697-16; 6 – Л678-15; 7 – Л665-15; 8 – Л700-15; 9 – Л699-15; 10 – ВК920; 11 – ВК917;
12 – СЛ 24 Б; 13 – СЛ 4 Б; 14 – ВК 935 Б;
15 – ВК 934 Б; К – отрицательный контроль; М – маркер молекулярного веса
На рисунке 4 показаны результаты амплификации ДНК этих линий с праймером НАР3. На первой дорожке фрагменты ДНК линии НА-335, далее на дорожках 2–11 представлены линии с разной устойчивостью к расам P. hals-tedii. На дорожке 2 – линия Л2018-1, устойчивая к расам 330, 334, 730 и 710. Далее на дорожках 3–9 представлены фрагменты ДНК линий, устойчивых к расам 330, 730 и 710, но восприимчивых к расе 334, а на дорожках 10–11 – устойчивых только к расе 330, на дорожках 12–14 – восприимчивых ко всем четырем расам. Результаты показали, что ни у одной изученной линии не выявлено точно таких фрагментов ДНК, как у линии НА-335. У линий Л2018-1, Л689-15, Л693-15, Л697-15, Л678-15, Л700-15, ВК 920, ВК 917 с устойчивостью к одной или нескольким расам (дорожки 2–6, 8, 10, 11) не получено амплифицированных фрагментов. Мы предполагаем, что в их геноме отсутствует локус Pl6, а устойчивость к P. halstedii контролируется генами, относящимися к другим группам сцепления. У линий
Л665-15 и Л699-15 получены амплифи-цированные фрагменты ДНК размером от 988 до 1811 п.н. (рис. 4; дорожки 7, 9). У четырех линий, восприимчивых ко всем четырем расам, получены четкие фрагменты одинакового размера, но отличающиеся длиной от линий НА-335, Л665-15 и Л699-15 (рис. 4). Это согласуется с данными из литературных источников, так как праймер НАР3 продуцирует четыре фрагмента ДНК, один из которых соответствует линиям, восприимчивым к расам 100, 300, 700, 703, 710, 330, 770 и 730.
Заключение. В результате проведенного исследования был подобран и апробирован молекулярный STS-маркер НАР3, пригодный для идентификации локуса Pl 6 , контролирующего устойчивость подсолнечника к расам P. halstedii 100, 300, 700, 703, 710, 330, 770 и 730. Этот маркер может стать одним из звеньев системы ДНК маркеров для идентификации тесно сцепленных генов и проведения маркер-вспомогательной селекции на устойчивость подсолнечника к возбудителю ложной мучнистой росы.
Список литературы К вопросу о маркировании локусов Pl, контролирующих устойчивость подсолнечника к возбудителю ложной мучнистой росы
- Virányi F., Gulya T.J., Tourvieille de Labrouhe D. Recent changes in the pathogenic variability of Plasmopara halstedii (sunflower downy mildew) populations from different continents//Helia. -2015 -V. 38 -P. 149-162.
- Gulya T.J. Distribution of Plasmopara halstedii races from sunflower around the world//Proc. 2nd Int. Downy Mildew Symposium «Advances in downy mildew research». Olomouc, Czech Republic, 2007. -V. 3. -P. 121-134.
- Spring O., Zipper R. New highly aggressive pathotype 354 of Plasmopara halstedii in German sunflower fields//Plant Protection Science. -2018. -V. 54 (2). -P. 83-86 DOI: 10.17221/99/2017-pps
- Iwebor M., Antonova T., Araslanova N., Saukova S. To the question of sunflower resistance to the downy mildew pathogen in the Krasnodar region of the Russian Fedeation//Proc. Intern. Congress on oil and protein crops. Chisinau, Republic of Moldova. -2018. -P. 133.
- Zimmer D.E., Kinman M.L. Downy mildew resistance in cultivated sunflower and its inheritance//Crop Sci. -1972. -V. 12 (6). -P.749-751. DOI: 10.2135/cropsci1972.0011183X001200060009x.
- Zimmer D.E. Physiological specialization between races of Plasmopara halstedii in America and Europe//Phytopathol. -1974. -V. 64. -P. 1465-1467
- DOI: 10.1094/Phyto-64-1465
- Liu Z, Gulya T.J., Seiler G.J., Vick B.A., Jan C-C. Molecular mapping of the Pl16 downy mildew resistance gene from HA-R4 to facilitate marker-assisted selection in sunflower//Theor. Appl. Genet. -2012. -V. 125. -P. 121-131.
- Jocić S., Miladinović D., Imerovski I., Dimitrijević A., Cvejić S., Nagl N., Kondić-Špika A. Towards sustainable downy mildew resistance in sunflower//Helia. -2012. -V. 35. -No 56. -P. 61-72.
- Radwan O., Bouzidi M.F., Vear F., Philippon J., Tourvieille de Labrouhe D., Nicolas P., Mouzeyar S. Identification of non-TIR-NBS-LRR markers linked to the P15/P18 locus for resistance to downy mildew in sunflower//Theoretical and Applied Genetics. -2003. -V. 106. -P. 1438-1446
- DOI: 10.1007/s00122-003-1196-1
- Gulya T.J., Sackston W.E., Viranyi F., Masirević S., Rashid K.Y. New races of the sunflower downy mildew pathogen (Plasmopara halstedii) in Europe and North and South America//Journal of Phytopathology. -1991. -V. 132 (4). -P. 303-311
- DOI: 10.1111/j.1439-0434.1991.tb00125.x
- Zolan M.E., Pukkila P.J. Inheritance of DNA methylation in Corpinus cinereus//Mol. Cell Biol. -1986. -Т. 6. -No 1. -Р. 195-200.
- Saghai-Maroof M.A., Soliman K.M., Jorgensen R.A., Allard R.W. Ribosomal DNA spacer-length polymorphisms in barley: Mendelian inheritance, chromosomal location, and population dynamics//PNAS USA. -1984. -V. 81. -P. 8014-8018.
- Bouzidi M.F, Badaoui S., Cambon F., Vear F., Tourvieille de Labrouhe D., Nicolas P., Mouzeyar S. Molecular analysis of a major locus for resistance to downy mildew in sunflower with specific PCR-based markers//Theoretical and Applied Genetics -2002. -V. 104. -P. 592-600.
- Panković D., Radovanović N., Jocić S., Satović Z., Škorić D. Development of co-dominant amplified polymorphic sequence markers for resistance to sunflower downy mildew race 730//Plant Breeding. -2007. -V. 126 (4) -P. 440-444
- DOI: 10.1111/j.1439-0523.2007.01376.x
- Tang S., Yu J.-K., Slabaugh M.B., Shintani D.K., Knapp, S.J. Simple sequence repeat map of the sunflower genome//Theoretical and Applied Genetics. -2002 -V. 105 (8) -P. 1124-1136
- DOI: 10.1007/s00122-002-0989-y
- Gascuel Q., Martinez Y., Boniface M.-C., Vear F., Pichon M., Godiard L. The sunflower downy mildew pathogen Plasmopara halstedii//Molecular Plant Pathology. -2014. -V. 16 (2). -P. 109-122
- DOI: 10.1111/mpp.12164
- Qi L.L., Talukder Z.I., Hulke B.S., Foley M.E. Development and dissection of diagnostic SNP markers for the downy mildew resistance genes Plarg and Pl8 and marker-assisted gene pyramiding in sunflower (Helianthus annuus L.)//Molecular Genetics and Genomics. -2017. -V. 292 (3). -P. 551-563
- DOI: 10.1007/s00438-017-1290-8
- Dußle C.M., Hahn V., Knapp S.J., Bauer E. Plarg from Helianthus argophyllus is unlinked to other known downy mildew resistance genes in sunflower//Theoretical and Applied Genetics. -2004. -V. 109 (5). -Р. 1083-1086
- DOI: 10.1007/s00122-004-1722-9
- Imerovski I., Dimitrijević A., Miladinović D., Jocić S., Dedić B., Cvejić S., Surlan-Momirović G. Identification and validation of breeder-friendly DNA markers for Plarg gene in sunflower//Molecular Breeding -2014. -V. 34 (3). -P. 779-788
- DOI: 10.1007/s11032-014-0074-7
- Wieckhorst S., Bachlava E., Dußle C.M., Tang S., Gao W., Saski C., Bauer E. Fine mapping of the sunflower resistance locus PlARG introduced from the wild species Helianthus argophyllus//Theoretical and Applied Genetics. -2010. -V. 121 (8). -Р. 1633-1644
- DOI: 10.1007/s00122-010-1416-4
- Bert P.F., Tourvieille de Labrouhe D., Philippon J., Mouzeyar S., Jouan I., Nicolas P., Vear F. Identification of a second linkage group carrying genes controlling resistance to downy mildew (Plasmopara halstedii) in sunflower (Helianthus annuus L.)//Theoretical and Applied Genetics. -2001. -V. 103 -P. 992-997.
- Bertero de Romano A., Romano C., Bulos M., Altieri E., Sala C. A new gene for resistance to downy mildew in sunflower//Proc. Intern. Symposium "Sunflower Breeding on Resistance to Diseases", Russia, Krasnodar, 2010, June 23-24. -P. 141-146.
- Mulpuri S., Liu Z., Feng J., Gulya T. J., Jan C.-C. Inheritance and molecular mapping of a downy mildew resistance gene, Pl13 in cultivated sunflower (Helianthus annuus L.)//Theoretical and Applied Genetics. -2009. -V. 119 (5). -P. 795-803
- DOI: 10.1007/s00122-009-1089-z
- Liu Z., Gulya T.J., Seiler G.J., Vick B.A., Jan C-C. Molecular mapping of the Pl16 downy mildew resistance gene from HA-R4 to facilitate marker-assisted selection in sunflower//Theoretical and Applied Genetics. -2012. -V. 125. -P. 121-131.
- Rahim M., Jan C.C., Gulya T.J. Inheritance of resistance to sunflower downy mildew races 1, 2 and 3 in cultivated sunflower//Plant Breed. -2002. -V. 121. -P. 57-60.
- Molinero-Ruiz M.L., Domínguez J., Melero-Vara J.M. Races of isolates of Plasmopara halstedii from Spain and Studies on their virulence//Plant Disease -2002. -V. 86 (7). -P. 736-740
- DOI: 10.1094/pdis.2002.86.7.736
- Gedil M.A., Slabaugh M.B., Berry S., Johnson R., Michelmore R., Miller J., Gulya T., Knapp S.J. Candidate desease resistance genes in sunflower cloned using consereved nucleotide-binding site motifs: Genetic mapping and linkage to the downy mildew resistence gene Pl1//Genom. -2001. -V. 44 (2). -Р. 205-212.
- Ahmed S., Tourvieille de Labrouhe D., Delmotte F. Emerging virulence arising from hybridisation facilitated by multiple introductions of the sunflower downy mildew pathogen Plasmopara halstedii//Fungal Genetics and Biology -2012. -V. 49 (10) -P. 847-855
- DOI: 10.1016/j.fgb.2012.06.012
