Качество семян и особенности их прорастания у Begonia grandis в условиях защищенного грунта

Автор: Буглова Л.В., Фершалова Т.Д.
Журнал: Вестник Красноярского государственного аграрного университета @vestnik-kgau
Рубрика: Агрономия
Статья в выпуске: 9, 2025 года.
Бесплатный доступ
Цель исследования – изучение особенностей семенного размножения Begonia grandis Dryand. в условиях защищенного грунта и искусственного опыления в Сибири. В опытах были задействованы пять генетически самостоятельных растений, не являющихся потомками одного клона. Для определения коэффициента семенификации использован оригинальный метод учета развитых микроскопических семян и абортированных семязачатков по микрофотографиям; этот показатель варьирует в пределах 33–66 %. Выделено четыре фракции семян по выполненности: выполненные; выполненные с незначительными отклонениями; слабо выполненные (складчатые); невыполненные. Нормально развитые зародыши находятся в первых двух фракциях семян, они продолговатоовальные, крупные относительно размеров семени: (306 ± 8) мкм длиной, (152 ± 5) мкм шириной. Семена B. grandis имеют размеры (423 ± 5)–(435 ± 5) мкм, (218 ± 6)–(237 ± 3) мкм соответственно и являются эмбриональными: для них характерен крайне низкий запас питательных веществ и слабо выраженный диффузный эндосперм. Уточнены морфологические особенности проростков. Установлено, что семена начинают прорастать на 5й день. На 14й день всхожесть достигает плато и составляет 54–78 %. У проростков своеобразные ростовые потенции: наиболее развит и первым приступает к росту гипокотиль, затем формируются корневые волоски, на следующем этапе сбрасывается семенная кожура и расправляются семядоли. Впервые установлено, что формирование конуса нарастания у B. grandis начинается после того, как проростки приступили к фотосинтетической активности. Зачатки семядолей служат, в силу малых размеров, только для фотосинтеза. Изза крайне малого запаса питательных веществ в семенах жизнь проростка зависит от скорейшего перехода к автономному питанию. Адаптивными приспособлениями для такого быстрого перехода служат отсутствие визуально диагностируемой плюмулы и раннее развитие корневых волосков на начальных этапах прорастания.
Begonia grandis, зародыш, защищенный грунт, коэффициент семенификации, проростки, прорастание, семена
Короткий адрес: https://sciup.org/140312339
IDR: 140312339 | УДК: 582.681.61+581.48+631.531 | DOI: 10.36718/1819-4036-2025-9-45-56
Seed quality and germination characteristics of Begonia grandis in protected ground conditions
The aim of the study was to investigate the characteristics of seed propagation of Begonia grandis Dryand. in protected ground conditions and artificial pollination in Siberia. Five genetically independent plants that were not descendants of the same clone were used in the experiments. To determine the seedification coefficient, an original method of accounting for developed microscopic seeds and aborted ovules using microphotographs was used; this indicator varies within 33–66 %. Four fractions of seeds were distinguished by their completeness: complete; complete with minor deviations; poorly complete (folded); uncompleted. Normally developed embryos are found in the first two fractions of seeds; they are oblongoval, large relative to the size of the seed: (306 ± 8) μm in length, (152 ± 5) μm in width. The seeds of B. grandis have the sizes of (423 ± 5)–(435 ± 5) μm, (218 ± 6)–(237 ± 3) μm, respectively, and are embryonic: they are characterized by an extremely low reserve of nutrients and a poorly expressed diffuse endosperm. The morphological features of the seedlings have been clarified. It has been established that the seeds begin to germinate on the 5th day. On the 14th day, the germination reaches a plateau and is 54–78 %. The sprouts have unique growth potential: the hypocotyl is the most developed and the first to start growing, then the root hairs are formed, at the next stage the seed coat is shed and the cotyledons straighten out. It was established for the first time that the formation of the growth cone in B. grandis begins after the sprouts have started photosynthetic activity. The cotyledon rudiments serve, due to their small size, only for photosynthesis. Due to the extremely small supply of nutrients in the seeds, the life of the sprout depends on the fastest possible transition to autonomous nutrition. The adaptive devices for such a rapid transition are the absence of a visually diagnosable plumule and the early development of root hairs at the initial stages of germination.
Текст научной статьи Качество семян и особенности их прорастания у Begonia grandis в условиях защищенного грунта
Funding : the study was carried out according to state assignments of the Central Siberian Botanical Garden of the Siberian Branch of the Russian Academy of Sciences: AAAA-A21-121011290025-2 "Analysis of biodiversity, conservation and restoration of rare and resource plant species using experimental methods"; AAAA-A21-121011290027-6 Theoretical and applied aspects of studying the gene pools of natural plant populations and conservation of plant diversity outside the typical habitat ( ex situ ).
Введение. Begonia L. (сем. Begoniaceae) – один из 10 крупнейших родов цветковых растений, включающий более 2100 видов [1]. В роде выделяют 70 секций, B. grandis является типовым видом секции Diploclinium [2]. Представители этого таксона распространены от 97.45° до 121.78° в.д. и от 22.98° до 40.67° с.ш. [3] и произрастают на высоте до 3400 м над уровнем моря. B. grandis имеет обширный естественный ареал, что обусловлено высокой экологической пластичностью его представителей, способных произрастать в довольно широком температур- ном диапазоне. Это самый холодостойкий вид рода и единственный, у которого клубни могут зимовать при отрицательных температурах [1]. Недавно обнаружены естественные популяции B. grandis на юге Тайваня [4]. Растения этого вида встречаются в долинах, преимущественно под пологом леса, по берегам рек, среди скал.
-
B. grandis имеет тысячелетнюю историю выращивания в Китае и Японии. Этот вид ценится за высокие декоративные качества, а также используется как лекарственное и пищевое растение [3, 5]. Метанольный экстракт Begonia grandis
показал гепатопротекторный эффект при гепато-токсичных повреждениях [6] B. grandis, рекомендован в качестве декоративной культуры для создания цветочного оформления объектов ландшафтной архитектуры в Западной Сибири [7].
-
B. grandis – малолетник с однолетними или малолетними клубнями почти шаровидной формы 8–12 мм в диаметре. Побеги в условиях интродукции достигают 28–60 см в высоту и почти лишены опушения [8]. В природе могут достигать высоты 1 м. Соцветие – открытый фрондозный тирс [9]. Цветонос имеет длину 10– 15 см, на нем образуются, как правило, 10–30 раздельнополых цветков. Сначала раскрываются мужские цветки, затем женские, составляющие 10–15 % от общего числа цветков. Завязь нижняя, образованная тремя плодолистиками, гинецей синкарпный. Плод – висячая, продолговатая коробочка, 10–12 × ~7 мм, неравномерно трехкрылая; абаксиальное крыло продолговатое или треугольно-продолговатое, около 1,8 см, боковые крылья узкотреугольные; реже все крылья узкотреугольные. После созревания плоды продольно растрескиваются, но не опадают. Семена B. grandis, как и других представителей семейства Begoniaceae, с тонкой семенной кожурой, характеризуются наличием поперечного кольца из так называемых воротничковых клеток, демонстрируют разнообразие по размеру, форме и микроморфологическим признакам. Одновременно с цветением особи в пазухах листьев закладываются специализированные пазушные почки, обеспечивающие вегетативное размножение. После цветения надземная часть растения отмирает. Растение входит в состояние покоя в виде подземного клубня.
Представители B. grandis являются модельными объектами для изучения морфологии, физиологии, фитохимии, возможностей селекции, микроклонального размножения в природе и при интродукции [10–17]. Несмотря на способность к вегетативному размножению, семенное воспроизводство бегоний представляет научный и практический интерес, поскольку особи, собранные в естественных местообитаниях, являются источником ценных признаков для селекции, в том числе устойчивости к абиотическим и биотическим стрессам [3]. У бегоний семена очень мелкие, их длина варьирует в широких пределах: от 220 до 2240 мкм. У большинства видов бегоний длина семян составляет от 300 до 600 мкм [11, 18,19]. Число семян на плод сущес- твенно варьирует: обычно оно находится в пределах 50–200 шт., однако в плодах B. schmi-dtiana Regel. формируется около 6500 семязачатков [10]. Данные о наличии запасающих тканей в семенах противоречивы. F.D. Boesewinkel и A. Lange [18] описали однослойный эндосперм, однако другие авторы отмечают его отсутствие при слабо или хорошо дифференцированном зародыше [20]. Крайне малые размеры семян значительно затрудняют исследования семенного воспроизводства бегоний, в частности данные по морфометрии семян и сведения о семенном воспроизводстве B. grandis отсутствуют.
В практике обычно используют различные методики проращивания семян бегоний: на фильтровальной бумаге либо свободно плавающими на поверхности воды. Контейнерами для проращивания могут быть чашки Петри и стеклянные трубки [10]. В лабораторных условиях прорастание обычно происходит в течение трех недель, а всхожесть может варьировать от 0 до 90 %. Для B. grandis subsp. evansiana приводятся данные о всхожести 60–70 % при 750 лк освещения и температуре 29 ºС [21]. R.S. Haba [10] установил, что оптимальные условия проращивания семян данного подвида в теплице – 24–27 °C при высокой влажности воздуха (60 %). Температурный интервал, в пределах которого сохраняются высокие показатели прорастания, составляет около 10 °C (от 20 до 29 °C). Семена данного подвида являются положительно фотобластическими: освещение 50 мкмоль·м – 2·с – 1 (300 лк) достаточно для их нормального прорастания. При длительном (2–52 недель) затемнении семена B. grandis subsp. evansiana способны к вторичному покою [10]. При хранении семян в темноте их общая всхожесть снижается уже через 2 недели, однако это не свидетельствует о непрерывной потере жизнеспособности, так как через 12 недель проращивание давало более низкие результаты, чем через 52 недели в аналогичных условиях. Через шесть месяцев хранения семян в темноте их всхожесть повышалась на 17 % по сравнению со свежесобранными, что указывает на необходимость дальнейших исследований влияния темной инкубации.
Наблюдения за прорастающими семенами бегонии показали, что первоначально гипокотиль, а не корешок, обеспечивает механическое давление для разрыва семенной оболочки [10, 15]. При этом микропилярная часть (кры- шечка) отделяется по предварительно сформированным линиям разрыва. Вскоре после удлинения гипокотиля начинают формироваться корневые волоски. Предполагается, что как только семядоли начинают фотосинтезировать, развивается корешок, который внедряется в субстрат, формируя корневую систему. Выход зародыша из семенной кожуры гипокотилем, а не корнем, как у большинства двудольных растений, является характерной чертой семян семейств Begoniaceae и Balsaminaceae [10].
Цель исследования – изучение особенностей семенного размножения Begonia grandis Dryand. в условиях защищенного грунта и искусственного опыления в Сибири.
Задачи: изучить морфометрические показатели семян; определить коэффициент семени-фикации; выделить фракции семян по их выполненности; определить пригодность семян для посева по морфологии и жизнеспособности зародышей; изучить особенности прорастания и морфогенетическую зональность проростков.
Объекты и методы. В качестве образцов для исследования использованы семена, собранные с 5 растений (генет) B. grandis, которые выращивались в условиях закрытого грунта в Центральном сибирском ботаническом саду СО РАН (г. Новосибирск). Исходный материал был получен из ботанического сада города Куньминь (Китай) в 2001 г. Семена для изучения собирали после однократного искусственного опыления, произведенного с помощью кисточки. Чтобы исключить случайное перекрестное опыление с другими видами бегоний, отобранные образцы содержались изолированно от общей коллекции бегоний. Во время созревания семян в оранжерее поддерживали температуру 20–27 °С, влажность воздуха 50–70 % и освещенность 1000 лк.
Семена были собраны в декабре 2023 г. (образец № 1) и в декабре 2024 г. (образцы № 2–5).
Количественные показатели семенной продуктивности имеют большое значение для оценки жизнеспособности вида в определенных условиях обитания или при интродукции. Но, поскольку семена B. grandis имеют микроскопические размеры, провести классический учет семенной продуктивности с определением реальной и потенциальной семенной продуктивности по методике И.В. Вайнагий [22], выполняемый «невооруженным глазом», крайне затруднительно. Микроскопические размеры семян и еще более мелкие абортированные се- мязачатки легко электризуются и разлетаются от малейшего движения воздуха, что приводит к ошибкам при их учете. Поэтому мы модифицировали методику определения коэффициента семенификации (КС), без определения реальной и потенциальной семенной продуктивности, методика учета которых не разработана для микроскопических семян. Коэффициент семе-нификации определяли с применением оригинальной методики: учитывали выполненные и невыполненные семена на микрофотографиях в трех полях зрения микроскопа. Для этого плоды бегоний на 2 часа помещались во влажную камеру – в чашку Петри с увлажненной фильтровальной бумагой, чтобы увеличить массу семян и уменьшить эффект электростатики. Далее семена высыпали в чашку Петри, потряхивая разломанный плод так, чтобы высыпались все семена, включая невыполненные (рис. 1), пинцетом убирали крупные части околоплодника и фотографировали семена под микроскопом. Эмпирически было установлено, что достаточным объемом выборки является учет в трех полях зрения микроскопа, при условии, что в каждое поде попадет не менее 25 семян. Учитывали семена 1–2 фракций (рис. 1) как аналог реальной семенной продуктивности, а все семена, включая усохшие семязачатки, – как аналог потенциальной семенной продуктивности. КС рассчитывали как отношение числа развитых семян к общему числу семяпочек, выраженное в процентах; в таблицу заносили среднее значение по трем повторностям. Этот параметр, подобно КС, вычисленному стандартным способом (по методике И.В. Вайнагий), показывает отношение развитых семян к общему числу семязачатков, но рассчитывается в полях зрения микроскопа, а не в абсолютных показателях реальной и потенциальной семенной продуктивности. Эмпирическим путем было установлено, что учет параметров в трех полях зрения является достаточным при условии, что в каждое поле зрения попало не менее 25 семязачатков.
Для изучения морфологии зародышей и начальных этапов прорастания, их извлекали из семенной кожуры после предварительного выдерживания в закрытой чашке Петри на увлажненной фильтровальной бумаге в течение двух часов. Далее их выдавливали легким нажатием препаровальной иглы на семенную кожуру сбоку. Для лучшей визуализации зародыши и про- ростки окрашивали в 1 % спиртовом растворе сафранина или в пропионовом кармине.
Для определения лабораторной всхожести семена раскладывали в чашки Петри диаметром 9 см с двумя слоями фильтровальной бумаги, смоченной 2 мл дистиллированной воды. В течение эксперимента ежедневно добавляли 1 мл воды, поддерживая влажность воздуха в чашках Петри на уровне 70–90 % при температуре 27 °С. Семена проращивали в двух вариантах освещенности: 1) при 1000 лк с фотопериодом 14 ч; 2) в полной темноте. Опыты закладывали в трех повторностях по 70 шт. семян в каждой, в таблицу заносили среднее значение.
При учете проросших семян проростки с отклонениями учитывали вместе с нормально развивающимися.
Разделение семян по фракциям выполняли с помощью микроскопа МСП1, микрофотографирование – с использованием микроскопа Primo Star iLED (Carl Zeiss Microscopy GmbH, Germany), оснащенного камерой AxioCamMRc (5 мп) и программой для обработки изображений AxioVi-sion 4.8 (Carl Zeiss Microscopy GmbH, Germany).
Результаты и их обсуждение . Период созревания семян B. grandis в научной коллекции ЦСБС СО РАН составил около 40 дней. В каждом плодолистике закладывалось 100–120 шт. семязачатков, соответственно, учитывая число плодолистиков, 300–360 шт. на плод.
Морфолого-анатомические особенности семян. Семена B. grandis светло-коричневого цвета. Семенная кожура подразделяется на 3 зоны: крышечка семени, клетки воротничка, клетки тесты (рис. 1). Теста и тегмен тонкие, однослойные, полупрозрачные, зародыш слегка просматривается через семенную кожуру. Размер семян: – (423 ± 5) – (435 ± 5) мкм длиной, (218 ± 6) – (237 ± 3) мкм шириной.
Нормально развитые зародыши относительно крупные – занимают около 3/4 размера семени, они (306 ± 8) мкм длиной, (152 ± 5) мкм шириной, удлиненно-овальной формы. Клеточная структура эндосперма отсутствует, но в увлажненном состоянии вокруг зародыша наблюдаются его остатки в виде рыхлой бесформенной массы. Семядоли короткие, составляют около 1/3 длины зародыша, а гипокотиль – корневая ось зародыша – соответственно 2/3 (рис. 2). Запасающие включения в клетках семядолей не обнаружены, в сочетании с их малыми размерами можно заключить, что запасающая функция у них отсутствует. При этом гипокотиль составляет основную часть длины зародыша, а корневая часть имеет всего 1 (возможно 2) слоя клеток. Плюмула не визуализируется, апикальная меристема не образует конуса нарастания в основании семядолей. По признакам: крупный зародыш, окруженный диффузным эндоспермом, семена B. grandis относятся к эндоспермальным.

Рис. 1. Дифференциация семян Begonia grandis по выполненности: 1 – выполненные; 2 – слабо выполненные; 3 – складчатые; 4 – невыполненные
Differentiation of Begonia grandis seeds based on the fullness property. 1. completed;
2. poorly completed; 3. folded; 4. not completed
Нами было выделено 4 фракции семян по степени выполненности, форме и размерам (рис. 1).
-
1. Выполненные семена овальной формы, светло-коричневые, нормально развитые.
-
2. Выполненные семена с незначительными отклонениями по форме (не овальные, а су-
- жающиеся к верху или с вмятинами) или по цвету (более темные).
-
3. Слабовыполненные складчатые семена, сравнительно крупные.
-
4. Невыполненные, мелкие, щуплые семена.
Наличие зародыша и степень его развития определяли отдельно для каждой фракции.
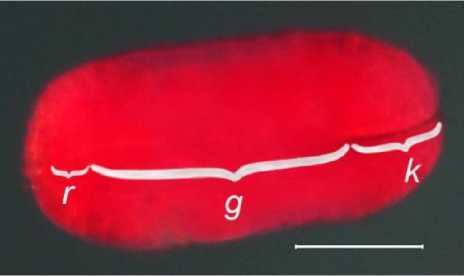
Рис. 2. Зародыш Begonia grandis, окрашенный спиртовым сафранином в течение 1 часа: k – семядоли, g – гипокотиль, r – радикула. Масштабная линейка: 100 мкм Begonia grandis embryo stained with alcoholic safranin for 1 hour: k – cotyledones, g – hypocotyl, r – radicula. Scale bar: 100 µm
В первой фракции 100 % семян содержали развитый зародыш естественной продолговатоовальной формы (см. рис. 2). Во второй фракции у 9 % семян зародыш отсутствовал, у 11 % наблюдался дегенерирующий (мелкий, темный) зародыш, а еще 32 % имели развившийся, но неправильной формы зародыш (обычно с семядолями, сужающимися кверху); на долю нормально развитых зародышей, приходится около половины – 48 %. Зародыши в семенах 3-й и 4-й фракций отсутствуют, четвертая фракция фактически представляет собой неоплодотворенные семязачатки, вокруг кото- рых, тем не менее, сформировалась семенная кожура. Следовательно, семена можно объединять в 2 категории: фракции 1 и 2 пригодны для посева, а фракции 3 и 4 можно отбраковывать как заведомо нежизнеспособные.
Коэффициент семенификации у 4 образцов довольно низкий, составляет 33–38 % и у одного образца существенно от них уклоняется, достигая 66,65 % (табл.). Доля прорастающих семян от общего числа заложившихся семяпочек у B. grandis в условиях защищенного грунта и искусственного опыления составляет 18,51–24,81 %.
Показатели качества семян и их прорастания у пяти образцов Begonia grandis (лабораторные условия, семена получены от искусственного опыления), % Seed quality and germination of five Begonia grandis samples (laboratory conditions,seeds obtained from artificial pollination)
|
Параметр |
Номер образцов |
||||
|
1 |
2 |
3 |
4 |
5 |
|
|
КС (в полях зрения микроскопа) |
66,65 |
37,96 |
32,77 |
34,11 |
36,61 |
|
Доля проросших семян на свету |
54,30 |
75,71 |
54,28 |
||
|
Доля проросших семян в темноте |
15,70 |
7,14 |
1,42 |
||
Проращивание семян. Набухание семян в лабораторных условиях отмечалось на третий день проращивания. К этому времени семена незначительно увеличились в размерах, но про- растания еще не наблюдалось. В этот период происходит дифференциация тканей: формирование проводящей системы, обособление клеток корешкового пояса и хлоренхимы. Отсоеди- нение крышечек семенной кожуры началось на 5-й день. Дальнейшее прорастание происходило стремительно – в течение суток удлинялся гипокотиль, в зоне перехода к корневой части развивался «корешковый пояс» – однослойный валик крупных клеток, быстро формирующих корневые волоски (рис. 3, А). Семядоли приобретают зеленую окраску до сбрасывания семенной кожуры. Примерно через 30 ч после отсоединения крышечки семядоли сбрасывали семенную кожуру и расправлялись. С этого момента растение переходило к ассимиляционной активности.
На рисунке 3, Б, левый проросток перед выходом семядолей из семенной кожуры, при этом плюмула редуцирована, апикальная меристема расположена горизонтальным диском под эпидермисом, стрелка указывает на центральную зону апикальной меристемы. У правого проростка семядоли вышли из семенной кожуры и развернулись, и только с этой стадии наблюдается выраженный конус нарастания. Центральная (осевая) зона апикальной меристемы побега порождает плюмулу, а периферическая в дальнейшем дает прирост коротких черешков семядолей. Корневой апекс также остается слабоактивным до сбрасывания семядолями семенной кожуры. Таким образом, рост апикальных меристем (корневой и побега) начинался только после разворачивания семядолей и начала автотрофного питания.
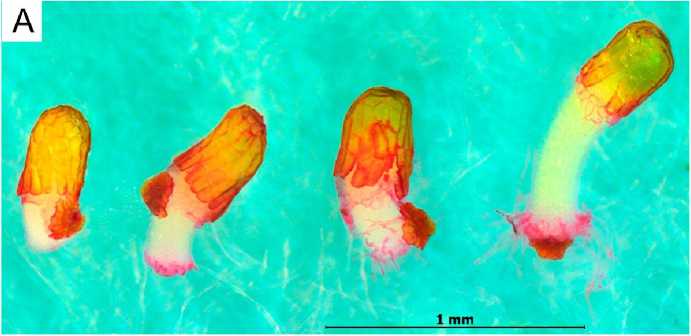
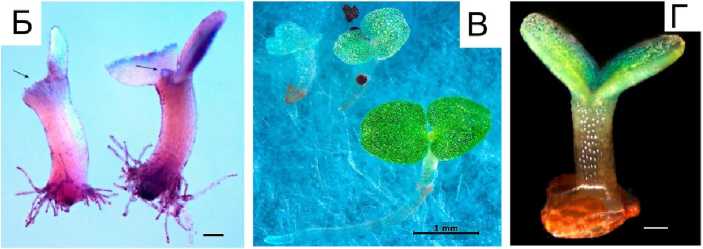
Рис.3. Проростки Begonoa grandis: А – начало прорастания: отделение крышечки, рост гипокотиля, рост корневых волосков, перед сбрасыванием семенной кожуры; Б – проросток, слева – перед сбрасыванием семенной кожуры с семядолей, справа – вскоре после сбрасывания семенной кожуры;
В – конгенитальный хлороз слева направо: полный, частичный; внизу справа – нормально развивающийся проросток; Г – проросток с неправильным выходом из семенной кожуры (А – подкрашивание сафранином 15 мин; Б – подкрашивание пропионовым кармином 15 мин;
В и Г – без окрашивания. Шкала: Б и Г – 100 мкм)
Begonia grandis seedlings, А – beginning of germination: operculum separation, hypocotyl growth, root hair growth, before seed coat shedding; Б – seedling on the left before seed coat shedding from cotyledons, on the right – soon after seed coat shedding; В – congenital chlorosis from left to right: complete, partial; bottom right - healthy seedling; Г – seedling with abnormal emergence from seed coat (А – stained with safranin for 15 minutes; Б – stained with propionic carmine for 15 minutes, В and Г – without staining. Scale bar: Б and Г – 100 µm)
Прорастание семян дружное: большая их часть во всех выборках прорастала на 5–8-й день, к 14-му дню увеличение числа проросших семян прекращалось (рис. 4). После этого срока семена больше не прорастали, и к 20-му дню все оставшиеся семена погибали.
Основная доля проростков (93–97 %) развивалась нормально, но около 3–7 % имели отклонения в развитии.
-
1. Конгенитальный хлороз семядолей: семядоли проростков имеют беловатую или бледнозеленую окраску (рис. 3, В ). Такие растения в дальнейшем отставали в развитии и всегда погибали. Случаев пестролистности семядолей не наблюдали.
-
2. Аномальный выход из семенной кожуры, заключающийся в том, что оболочка оставалась не на семядолях, а на основании гипокотиля (рис. 3, Г , норма на рис. 3, А ). У таких проростков резко замедлялся рост корневых волосков и всего растения. Если сбрасывание семенной кожуры в дальнейшем не происходило, то проростки погибали.
В условиях проращивания в темноте средний показатель всхожести существенно ниже, чем при освещении, составляет в среднем 8 %, при этом разброс всхожести весьма существенный – 1,4–15,7 % (см. табл.).
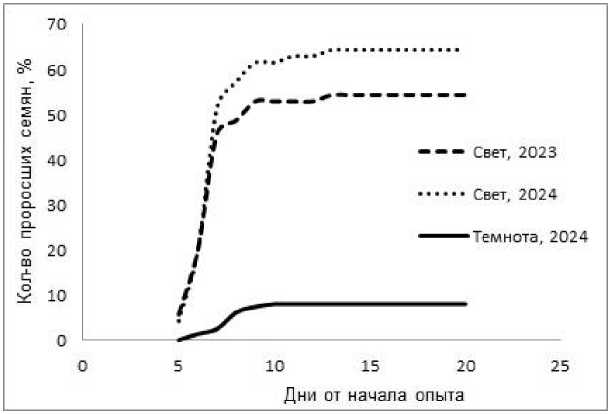
Рис. 4. Энергия прорастания семян Begonia grandis Begonia grandis seed germination energy
Семена B. grandis показали достаточно высокую всхожесть и дружное прорастание при освещении (рис. 4). Первые всходы появились на 5-й день, основная масса семян прорастала 6–8-й день, на 14-й день общая всхожесть составила 54,0–77,7 % на свету и 1,4–14,7 % в темноте.
Учет коэффициента семенификации (КС) по микрофотографиям выполнен нами впервые, он может быть удобен для видов с еще большим числом пылевидных семян по сравнению с Begoniaceae, например для представителей семейства Orchidaceae. При выращивании растений в условиях защищенного грунта и искусственном опылении реализация семенной продуктивности, определяемая по значению КС, учтенного на микрофотографиях, весьма изменчива у разных образцов: 33–66 %.
Благодаря определению КС по микрофотографиям можно оценить репродуктивный успех на стадиях от начала формирования семян до появления проростков. В данном опыте он определяется как произведение КС и доли проросших семян и составляет 18,51–24,81 %.
Сроки всхожести семян B. grandis в основном совпали с данными по семенам B. grandis subsp. evansiana [10]. Минимальные различия касаются начала прорастания – на 3-й день по R.S. Haba [10], в отличие от наших наблюдений – на 5-й день и в чуть более высокой всхожести. Эксперименты по изучению всхожести можно прекращать на 14-й день от начала проращивания, что согласуется с данными R.S. Haba [10], у которого к этому сроку кривая всхожести также выходила на плато. В ходе эксперимента была подтверждена фотобластичность семян.
Для бегоний была установлен интересный морфогенез проростков: первым приступает к росту гипокотиль [10], в то время как у большинства двудольных растений первым удлиняется корень. При этом биполярные потенции осевых структур зародыша и проростка являются обычными для двудольных растений. Наш вклад в изучение прорастания семян бегоний заключался в уточнении последовательности активации ростовых процессов в начальные стадии их прорастания. Рост апикальных меристем с формированием конусов нарастания (корневого и побега) начинается поздно: после разворачивания семядолей и начала автотрофного питания, после того, как растения приступили к фотосинтетической активности, а не на стадии роста корневых волосков, как у большинства других видов. Так, например, у Begonia fischeri , судя по рисункам R.S. Haba [10, 43], формирование конуса нарастания у проростков начинается на 1 стадию развития раньше – сразу после появления корневых волосков. Мы считаем, что зафиксированное автором еще более раннее появление плюмулы у этого вида – непосредственно после начала роста гипокотиля, до появления корневых волосков, может являться артефактом наблюдения, поскольку на микрофотографии [10, стр. 43] проростка у предполагаемой плюмулы не выявляется клеточное строение.
Дискуссионным вопросом является происхождение корешкового пояса. Морфологически близкая к нему многослойная структура у Streptocarpus glandulosissimus была описана как имеющая гипокотилярное происхождение [23]. Тем не менее, поскольку функциями клеток корешкового пояса у B. grandis является образование корневых волосков, всасывающая и прикрепление к субстрату, не исключается их корневое происхождение. Ответ на этот вопрос требует дополнительных гистохимических исследований.
R.S. Haba [10] высказывает обоснованное предположение, что механизм прорастания гипокотилем – это адаптация к условиям высокой влажности, особенно на склонах вблизи водопадов. Такая адаптация включает также переход к крайне низкому запасу питательных веществ (со скудным эндоспермом и маленькой семядолей). Таким проросткам для дальнейшего развития жизненно необходимо как можно быстрее перейти к автономному питанию. Поэ- тому особенности морфологии зародышей, которые могут быть определены как недоразвитие (отсутствие визуализируемой плюмулы), на самом деле являются вторичным признаком, возникшим как приспособление для максимально быстрого перехода к автотрофному питанию. В практическом плане для повышения выживаемости таких эмбриональных семян целесообразно вносить дополнительное питание от самых ранних стадий развития.
Начальные этапы морфогенеза разных таксонов двудольных растений различны и адаптированы к разным условиям среды обитания. Их изучение позволяет уточнить типы прорастания [24]. У представителей семейства Bego-niaceae в рамках надземного двудольного прорастания реализован своеобразный подтип прорастания гипокотилем.
Заключение. В условиях закрытого грунта возможно получение семенного потомства B. grandis от искусственного опыления, так как реализация семенной продуктивности в этих условиях может достигать 66 %. При проращивании семян B. grandis на свету показатели всхожести (54,0–77,7 %) и темпы прорастания (в течение 14 дней после посева) достаточно высокие для данного вида.
Предложенная модификация метода определения коэффициента семенификации позволяет оптимизировать его учет для очень мелких (пылеобразных) семян.
Известно, что у B. grandis выделен особый подтип прорастания гипокотилем; нами впервые установлено, что визуализация плюмулы происходит на поздних стадиях развития проростка – после разворачивания семядолей. После завершения дифференциации первичных тканей, для его стабильного развития, необходимо вносить дополнительное питание, чтобы сохранить максимальное количество всходов, поскольку у семян и зародышей крайне низкий запас питательных веществ.
