Камерная модель хищник-жертва на кольцевом ареале

Автор: Гончарова А.Б., Колпак Е.П., Гасратова Н.А.
Журнал: Бюллетень науки и практики @bulletennauki
Рубрика: Естественные науки
Статья в выпуске: 2 т.11, 2025 года.
Бесплатный доступ
В работе исследуется система хищник- жертва на линейном ареале. В модели учитывается нелинейный рост численности жертвы. Ареал представлен системой зон, между которыми происходит обмен особями. Для локальной модели определены неподвижные точки и условия их устойчивости. Для камерной модели построены численные решения с неоднородным распределением особей по камерам. Неоднородность стационарного распределения наступает при высокой подвижности хищника.
Особи, популяция, устойчивость, положение равновесия, подвижность
Короткий адрес: https://sciup.org/14131828
IDR: 14131828 | УДК: 577.3.01(0.75) | DOI: 10.33619/2414-2948/111/05
The predator-prey compartment model in the ring area
The work examines the predator-prey system in a linear range. The model takes into account the non-linear growth in the number of victims. The area is represented by a system of zones between which individuals are exchanged. Fixed points and their stability conditions are defined for the local model. Numerical solutions with an inhomogeneous distribution of individuals across the chambers are constructed for the chamber model. Heterogeneity of distribution occurs when the predator is highly mobile.
Текст научной статьи Камерная модель хищник-жертва на кольцевом ареале
Бюллетень науки и практики / Bulletin of Science and Practice
УДК 577.3.01(0.75)
Локальные модели взаимодействующих популяций не учитывают пространственное распределение особей популяций. В экосистемах могут образоваться группировки особей, проживающие на небольших близлежащих участках экосистемы. Сами экосистемы могут быть протяженными в пространстве [1, 2]. Примерами таких территорий могут быть транспортные пути, побережья рек и озер, системы островов. Побережье озера может представлять собой кольцевой ареал, на котором живут различные виды, как на суше, так и в водной системе [3]. Линейный ареал – транспортные пути, вдоль которых распространяются различные виды. На линейном и кольцевом ареале заселение территории не является равномерным. Поэтому локальная модель для таких ареалов дает усредненные характеристики численности популяций, не объясняя неравномерность распределения особей по территории. Модель хищник-жертва на билокальном ареале была предложена в работе А. Д. Базыкина [4]. В статье рассматривается модель хищник-жертва для многокамерной системы.
Локальная модель
Модель хищник-жертва, учитывающая квадратичный рост малочисленной популяции жертвы, разработана в [4]:
— = u2(1 — u) — uv, at
dv
-
-7- = —/(a — u)v,
dt где u — численность жертвы, v — численность хищника, α и γ — положительные параметры. Положение равновесия в системе уравнений (1) реализуются в следующих случаях:
-
1. u = 0, v = 0.
-
2. u = 1, v = 0.
-
3. и = a, v = a(1 — a)
Первое положение равновесия является неустойчивым [5] при любах значениях параметра a. Второе — устойчивым при a > 1, и неустойчивым при a < 1. Третье положение равновесия может реализовываться при выполнении неравенства a < 1, будет устойчивым только при a > 1/2.
При a < 1/2 все стационарные состояния (2) будут неустойчивыми. Функции u(t) и v(t) при значениях параметра a < 1/2 находятся в ограниченном фазовом пространстве, поэтому третье положение равновесия будет устойчивым по Лагранжу. Поскольку нулевая точка является неустойчивой, то при появлении у жертвы хищника могу возникать колебания численности жертвы и хищника [5]. На рисунке 1 для случая a = 0.45 показана зависимость функций u(t) и v(t) от времени. Начальная численность жертвы - u(t = 0) = 1.001a, и численность хищника - v(t = 0) = a(1 — a) приняты за малое отклонение от третьего положения равновесия. При малом возмущении третьего положения равновесия в (2) происходит потеря устойчивости и со временем начинаются периодические колебания в системе хищн
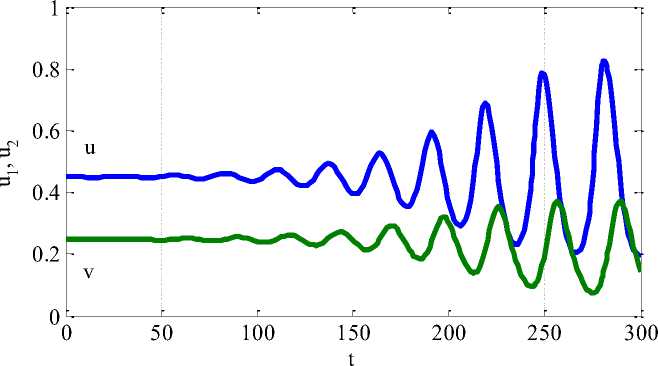
Рисунок 1. Изменение численности жертвы ( U ( t ) ) и хищника ( V ( t ) ) от времени
Камерная модель
Модель кольцевого ареала представлена последовательностью камер (зон), расположенных на замкнутой кривой единичной длины, расположенных на равных расстояниях друг от друга. Особи из одной камер могут перемещаться в любую из двух соседних. Для случая n камер модель кольцевого ареала на основе модели (1) имеет следующий вид:
du 1
— = F1(u1,v1) + cu(un
—
2u 1 + u2),
dV,
— = F2(u i ,V i ) + C v (V n — 2v i + v2); где F ( u , v ) = u (1 - u ) - uv , F ( u , v ) = —у ( a — u ) v .
В модели (3) cu параметр, характеризующий миграцию жертв в соседние камеры, а cV - хищников. cu и cV считаются разными для хищников и жертв, но одинаковыми для всех камер и постоянными во времени. F 1 и F2 - трофические функции одинаковые во всех камерах.
В модели (3) учитывается направленное движение не только хищника [6], но и жертвы. Миграция в соседние камеры считается постоянной [7] — cV и cu постоянные величины. Поскольку трофические функции во все камерах одинаковые, то распределение особей по камерам считается «свободным» [8] – допускается однородное распределение особей.
Система уравнений (3) имеет такие же стационарные точки, как и в случае одной камеры. При наличии миграции в системе уравнений (3) реализуется однородное положение равновесия такое, что u 1 = u2 =... = un = u * и v 1 = v2 =...= vn = v * , где (u * v * ) одно из положений равновесия (2). При а < 1/2 третья стационарная точка системы уравнений (3) неустойчивая - в ее окрестности возникают колебания. Для п камер система уравнений (3) может содержать и другие точки притяжения такие, что распределение особей по камерам не будет однородным.
Примеры неоднородного стационарного распределения хищника и жертвы для п = 20 камер приведены на Рисунке 2 и Рисунке 3. Символом «*» отмечены численности хищника и жертвы в камерах. Значения параметров: а = 0.45, у = 0.5. Предполагается, что миграция жертвы происходит медленно (cu = 0.01), а хищник перемещается по ареалу значительно быстрее, чем жертва: на рисунке 1 - cV = 10cV = 10 , на рисунке 2 - cV = 0.7 [4]. Пунктирными горизонтальными линиями на рисунке 2 и рисунке 3 отмечены однородные решения u i = а, v t = а(1 — а) (i = 1,2,...,п).
Вариант начальных данных u1 = 0.55, u2 = u3 =... = un = 0, v1 = 0.25, v2 = v3 =... = vn = 0
предполагает, что хищник и жертва в момент времени t = 0 появляются в первой зоне с последующим распространением по остальным зонам. Колебания во всех зонах со временем прекращаются – система переходит в устойчивое неоднородное по камерам положение равновесия. При низкой подвижности хищника возможно образование множества зон с малой численностью жертвы (Рисунок 3). Интенсивное движение хищника по ареалу может привести к возникновению лишь 2-3 группировок жертвы (Рисунок 2).
При низкой подвижности хищника (при cu < 0.5) колебания в системе сохраняются, но они будут происходить около положения равновесия собственного для каждой камеры, но с одинаковой частотой для всех камер. При большем числе камер могут образовываться и боле сложные распределения по камерам значений функций. К какой из нескольких точек притяжения со времен будет стремиться решение зависит от начального приближения. Полученные результаты согласуются с данными полевых наблюдений за системой хищник – две жертвы [1].
Численное моделирование осуществлялось в среде математического пакета Matlab. Высокая скорость вычислений (несколько секунд для системы из 50 камер) достигалась за счет векторизации вычислений. Использовалась схема повышенного порядка точности для уравнений (3), разработанная в [9].
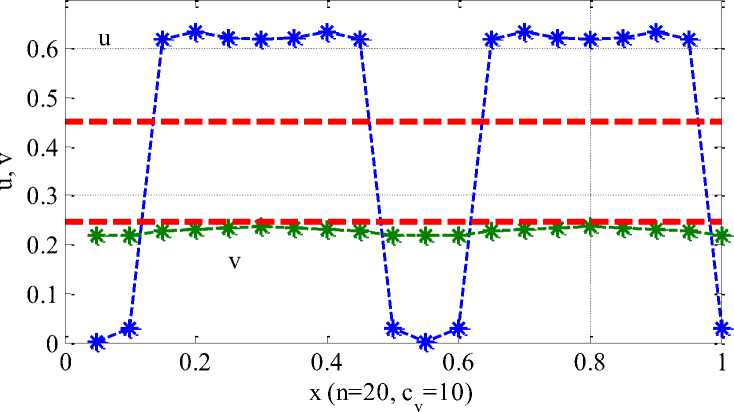
Рисунок 1. Распределение жертвы (и) и хищника (v) по 20 камерам в стационарной точке (c v = 10)
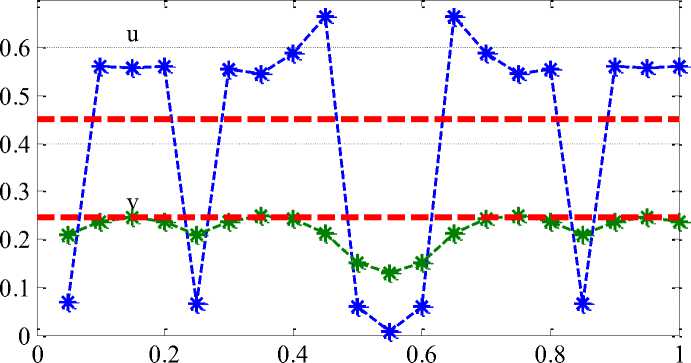
x (n=20, c =0.7)
Рисунок 2. Распределение жертвы (и) и хищника (v) по 20 камерам в стационарной точке (c v = 0.7)
Заключение
Камерная модель взаимодействия хищника и жертвы на линейном ареале может содержать большее число неподвижных точек, чем локальная. То есть распределение хищника, и жертвы по камерам может быть неравномерным. При высокой подвижности хищника его распределение на ареале стремится к равномерному распределению, а точка притяжения становится устойчивой. Жертва в отличие от хищника под его давлением может группироваться в отдельных зонах ареала.
Список литературы Камерная модель хищник-жертва на кольцевом ареале
- Данилов П. И., Каньшиев В. Я., Белкин В. В. Численность и распределение охотничьих животных в Карело-Мурманском крае в 2007 году. Петрозаводск, 2008. 30 с.
- Мысленков А. И., Волошина И. В., Шурыгина А. А., Керли Л. Ли. Местообитания, распространение и численность амурской рыси Lynx lynx stroganovi и дальневосточного лесного кота Prionailurus bengalensis euptilura в Лазовском заповеднике и национальном парке "Зов тигра" (Приморский край, Дальний Восток России) // Биота и среда природных территорий. 2023. Т. 11. №3. С. 27-52.
- Гончарова А. Б., Колпак Е. П., Гасратова Н. А. Модели антропогенного давления на экосистему. Казань: Бук, 2024. 102 с.
- Базыкин А. Д. Нелинейная динамика взаимодействующих популяций. М.: Институт компьютерных исследований, 2003. 368 с.
- Колпак Е. П., Гасратова Н. А., Селицкая Е. А Математическая модель хищник-жертва на ограниченной территории // Устойчивость и процессы управления: Материалы III международной конференции, 2015. С. 481-482.
- Гиричева Е. Е Влияние направленных перемещений хищника на формирование пространственных структур в модели трехвидового сообщества с учетом всеядности хищника // Компьютерные исследования и моделирование. 2023. Т. 15. № 6. С. 1617-1634.
- Курилова Е. В., Фрисман Е. Я. Моделирование динамики взаимодействующих популяций типа "хищник-жертва" при постоянной миграции особей с сопредельных территорий // Региональные проблемы. 2024. Т. 27. №1. С. 62-77.
- Зеленчук П. А., Цибулин В. Г. Математическая модель идеального свободного распределения в системе хищник-жертва // Современная математика. Фундаментальные направления. 2023. Т. 69. №2. С. 237-249.
- Нгуен Б. Х., Цибулин В. Г. Схема повышенного порядка точности для моделирования динамики хищника и жертвы на неоднородном ареале // Известия высших учебных заведений. Прикладная нелинейная динамика. 2024. Т. 32. №3. С. 294-304.
