Каноническая дискриминантная модель влияния генотипа отца на некоторые интерьерные показатели его потомков у свиней

Автор: Камалдинов Е.В.
Журнал: Вестник Красноярского государственного аграрного университета @vestnik-kgau
Рубрика: Животноводство
Статья в выпуске: 1, 2012 года.
Бесплатный доступ
Установлен ряд гематологических и биохимических показателей, по которым наблюдались наибольшие отличия между потомками разных производителей скороспелой мясной породы. Показана возможность использования стандартизированных и структурных коэффициентов для оценки влияния генотипа хряка на уровень лейкоцитов и общего белка у поросят в постотъемный период. Предлагаемый подход позволяет принимать во внимание сложную многомерную природу входных данных и учитывать многочисленные зависимости изучаемых количественных признаков.
Свиньи, порода, показатели крови, генотип, дискриминантный анализ, каноническая корреляция
Короткий адрес: https://sciup.org/14082009
IDR: 14082009 | УДК: 636.082:575.21:577.1
Canonical discriminant model of the father genotype influence on some interior indicators of its pig descendants
A number of hematological and biochemical indicators on which the greatest differences among the descendants of various early meat breed manufacturers are observed is determined. Possibility to use the standardized and structural coefficients in order to estimate a male pig genotype influence on the level of leukocytes and general protein in pigs in the post wean period is shown. The offered approach allows to take into consideration difficult multidimensional nature of the input data and to consider numerous dependences of the studied quantitative features.
Текст научной статьи Каноническая дискриминантная модель влияния генотипа отца на некоторые интерьерные показатели его потомков у свиней
Исследования в области гематологии и биохимии находят свое отражение в многочисленном количестве научных работ [1–4], в которых учитываются всевозможные корреляции, повторяемость или оценивается факториальная изменчивость. Вместе с тем известно, что такие показатели могут обладать высокой изменчивостью, в основе которой лежит многомерная природа их взаимодействий между собой и средовыми компонентами [5]. Таким образом, наследственная природа количественных признаков затрудняет понимание генетических и эволюционных факторов [6,7].
Некоторыми исследователями делаются предположения о генетической природе отличий животных по уровням ряда биохимических параметров [8]. Однако выраженность количественного признака может быть результатом накопительного эффекта, когда имеет место прямое или опосредованное воздействие одного или нескольких других факторов [9].
В этом случае не всегда представляется корректным определение коэффициентов повторяемости, применение критерия Стьюдента или проведение дисперсионного анализа, которые не позволяют в достаточной мере выделять наиболее выраженные факторы с учетом многомерного подхода. С этой целью предлагается использовать статистические подходы, позволяющие принимать во внимание такую многомерность входных данных [10,11]. Одним из широко распространенных методов выступает канонический дискриминантный анализ [12].
Целью наших исследований явилось определение отличий между потомками разных производителей по некоторым интерьерным показателям с последующей корректировкой полученных оценок с учетом существующих корреляционных взаимодействий.
Материал и методы исследований. Исследования проводились на поросятах возраста 19–21 день (n=288) и 2 недели после отъема (n=87) породы СМ-1. Для исследований использовалась база учебного хозяйства ГПЗ «Тулинское» Новосибирского государственного аграрного университета.
Определяли массу животных, базальную температуру, частоту дыхания и сердечных сокращений, скорость оседания эритроцитов и ряд биохимических, гематологических показателей в крови сразу после ее забора: уровень эритроцитов, лейкоцитов, гемоглобина, концентрацию гемоглобина в одном эритроците, цветовой показатель, лейкоцитарную формулу, коэффициент де Ритиса, активность общей кислой фосфатазы, общей и простатической кислой фосфатазы, аланинаминотрансферазы (АЛТ), аспартатаминотрансферазы (АСТ). Кальций-фосфорное отношение, уровень общего белка, холестерина, глюкозы, креатинина, хлоридов, кальция, фосфора и мочевины анализировали через 4 ч после взятия крови в течение первых суток и с учетом рекомендаций производителей наборов реагентов («Вектор-Бест» и «Biocon»). В качестве антикоагулянта применяли цитрат натрия и гепарин. Витамин С в пробах был стабилизирован с помощью раствора дитиотрейтола сразу после забора крови. Концентрацию лейкоцитов и эритроцитов определяли по общепринятой методике с применением камеры Горяева и фотометра. Содержание тяжелых металлов в щетине исследовали с помощью анализатора ТА-2. Всего в исследованиях проанализировано 36 параметров.
При обработке экспериментальных данных использовали среду статистического программирования «R» и свободно распространяемые библиотеки сетевого репозитария «CRAN».
Применение дискриминантного анализа сопровождалось нахождением канонических дискриминантных функций, являющихся линейной комбинацией дискриминантных переменных:
dkm = β 0 + β 1 x 1km + ... + βpxpkm
где d km – значение канонической дискриминантной функции для m -го ( m = 1,… ,n ) объекта в группе k ( k=1, ... ,g ) с неизвестными коэффициентами дискриминантной функции β i ;
x jkm – значение j -го дискриминантного параметра для m -го наблюдения из k -го класса.
При оценке величин вклада переменных в значение дискриминантной функции производили преобразование нестандартизированных коэффициентов в стандартизированные по формуле ci=βi
wii
\ n - g
где n – общее количество наблюдений;
g – число групп.
С целью классификации существующих групп и выявления возможного совместного вклада нескольких переменных в дискриминантной модели применяли структурные коэффициенты. Структурные коэффициенты определяли коэффициентами взаимной корреляции между переменными и дискриминантной функцией по формуле sij
p
= ∑ rikckj k= 1
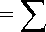
wikckj л/wiiwjj,
где s ij – структурный коэффициент, рассчитанный по i -й переменной и j -й функции;
r ik – внутригрупповые структурные коэффициенты корреляции между признаками i и k ; c kj – значения стандартизированных коэффициентов.
Коэффициенты канонической корреляции вычисляли по формуле [12]:
ri
λ i
\ 1 +λ i
,
где λ i – величина собственного значения.
Результаты исследований дисперсионный анализ позволил выявить гематологические и биохимические параметры крови поросят, по которым наблюдались отличия между потомками разных производите- лей. Получено 5 дискриминантых функций, характеризующих поросят-сосунов (группа 1), и 3 – поросят в возрасте 2 недели после отъема (группа 2) (табл. 1).
Результаты канонического дискриминантного анализа
Таблица 1
|
Дискриминантная функция |
Собственное значение |
Коэффициент канонической корреляции |
Λ -статистика Уилкса |
% |
|
Группа 1 |
||||
|
1 |
1,392 |
0,763 |
0,418 |
53,4 |
|
2 |
0,685 |
0,638 |
0,593 |
26,3 |
|
3 |
0,310 |
0,486 |
0,764 |
11,9 |
|
4 |
0,149 |
0,360 |
0,871 |
5,7 |
|
5 |
0,072 |
0,259 |
0,933 |
2,7 |
|
Группа 2 |
||||
|
1 |
2,634 |
0,851 |
0,275 |
72,8 |
|
2 |
0,583 |
0,607 |
0,632 |
16,1 |
|
3 |
0,403 |
0,536 |
0,713 |
11,1 |
Коэффициенты Уилкса в целом свидетельствовали о достаточном уровне детерминации дискриминантными функциями существующих отличий потомков разных хряков по ряду интерьерных показателей. Вместе с тем следует отметить, что более высокие значения Л- коэффициентов Уилкса установлены у поросят-сосунов. Это может объясняться наличием материнского эффекта и выражаться в изменении списка дискриминантных переменных для животных другой возрастной группы, что подтверждается данными таблицы 2. Обнаруженные особенности обусловлены, по всей видимости, поступлением в организм животных группы 1 элементов питания с молоком матери.
Таблица 2
Стандартизированные и структурные коэффициенты в различные периоды онтогенеза у свиней
|
Переменная |
Стандартизированный коэффициент |
Структурный коэффициент |
||||
|
Функция |
Функция |
|||||
|
1 |
2 |
3 |
1 |
2 |
3 |
|
|
Группа 1 |
||||||
|
Витамин С |
0,117649 |
0,061434 |
0,07081 |
0,26551 |
0,11077 |
-0,024800 |
|
Гемоглобин |
0,394409 |
-0,355518 |
-0,69078 |
-0,44359 |
-0,18651 |
-0,634556 |
|
Мочевина |
-0,201302 |
0,972918 |
0,13676 |
-0,34534 |
0,91350 |
-0,120471 |
|
АЛТ |
0,067447 |
-0,067420 |
0,99186 |
-0,65985 |
-0,30684 |
0,635736 |
|
АСТ |
-0,941983 |
-0,133681 |
-0,38812 |
-0,92745 |
-0,12181 |
0,273482 |
|
Группа 2 |
||||||
|
Лейкоциты |
-0,713132 |
-0,75823 |
0,12718 |
-0,58403 |
-0,80496 |
0,104594 |
|
Кальций |
0,075502 |
0,12386 |
1,15919 |
-0,47552 |
0,43180 |
0,766435 |
|
Общий белок |
-0,984034 |
0,39035 |
-0,58636 |
-0,85090 |
0,52500 |
-0,018364 |
Поросята-сосуны отличались между собой преимущественно по активности АСТ, которая характеризовалась высокими значениями стандартизированного и структурного коэффициентов. Проведенный корреляционный анализ позволил установить связь этого параметра с 21 другими гематологическими и биохимическими показателями (r= -0,291± – 0,594±), включая концентрацию витамина С, гемоглобина, мочевины и активность аланинаминотранферазы в сыворотке крови (r=-0,231±0,062*** – 0,751±0,041***). Наряду с этим, следует также обратить внимание на отличия по уровню мочевины и активности АСТ. Все установленные различия обусловлены накопительным эффектом. В результате происходит повышение степени сопряженности переменной дискриминантной функции со значением этой функции. Таким образом, применение ак- тивности АСТ и уровня мочевины у животных рассматриваемого возрастного диапазона в качестве биохимических маркеров, характеризующих генотип производителя в дальнейших исследованиях, представляется необоснованным.
Исследования крови поросят, отнятых от матери и успевших адаптироваться после этого стресса, позволили установить влияние генотипа хряка на некоторые интерьерные показатели. Так, согласно стандартизированным и структурным коэффициентам, генетическая компонента в большей степени прослеживается в отношении концентрации лейкоцитов (функции 1 и 2) по сравнению с содержанием общего белка в крови и другими признаками (см. табл. 2). Корреляционный анализ позволил выявить связь уровня лейкоцитов с содержанием хлоридов (r=0,459±0,115***), глюкозы (r=0,371±0,120**) в сыворотке крови и активностью АЛТ (r=-0,292±0,130*) и щелочной фосфатазы (r=0,390±0,134**). В то же самое время, уровень общего белка коррелировал с 10 показателями из 36 (r=-0,491±0,105*** – 0,569±0,099***). Таким образом, структурные коэффициенты дают адекватную оценку вклада сопряженности изучаемых гематологических и биохимических показателей в степень выраженности интересующих зависимых признаков.
Для характеристики потомства хряков визуализированы канонические оценки, позволяющие судить об изменчивости комплекса установленных интерьерных показателей у поросят-сосунов (рис.1). Особый интерес представляют сведения, полученные по потомкам Салюта-517, Сатурна-477, Светлого-1645, Соболя – 145, Соболя – 231 и Сома-69. Обнаруженные выбросы подчеркивают многомерную природу переменных дискриминантной модели и продиктованы действием, как генотипа хряка, так и материнским эффектом. Наименее консолидированным оказалось потомство Салюта-517, Совета-1813 и Сома-69 по сравнению с остальными. Наряду с изложенным, поросята таких производителей, как Саян-53 и Совет-1813 в наибольшей степени отличались по биохимическим параметрам от своих сверстников в подсосный период.
Канонические оценки
Структурные коэ
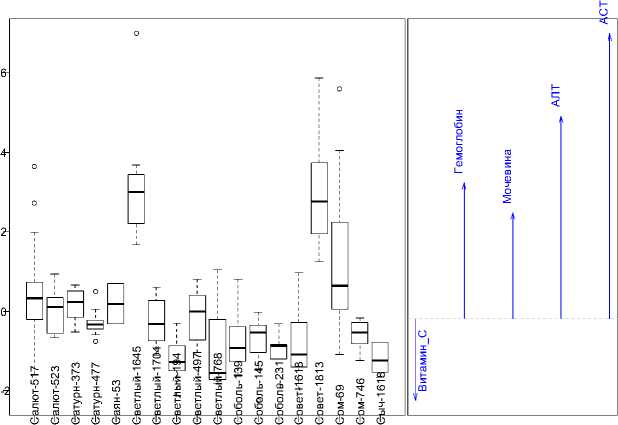
Рис.1. Отличия потомков производителей по гематологическим и биохимическим показателям крови в возрасте 19–21 день
Проведена оценка групповых средних уровней лейкоцитов, кальция и общего белка в крови потомков некоторых хряков через две недели после отъема (рис. 2). Характерной особенностью являлось отсутствие выбросов по всем изучаемым группам и изменение списка переменных канонической дискриминантной функции. С одной стороны, это объясняется нивелированием материнского эффекта группы 2 по сравнению с группой 1, где питательные элементы поступали в организм животных с молоком матери, с другой – особенностями обмена веществ растущего организма. В постотъемный период происходит интенсивный рост мышечной и костной ткани, что объясняет наличие таких переменных, как «кальций» (Ca) и «общий белок» (ОБ), которые также сопряжены со значением дискриминантной функции. Присутствие в модели переменной «лейкоциты» (Le) объясняется активным становлением иммунной системы свиней рассматриваемой возрастной группы.
Канонические оценки
Структурные коэ
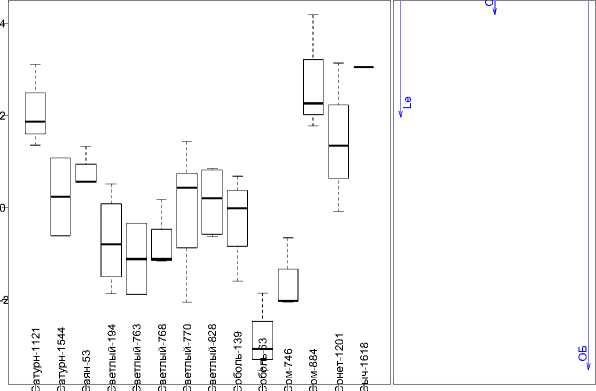
Рис. 2. Отличия потомков производителей по гематологическим и биохимическим показателям крови в возрасте 2 недели после отъема
Интересным представляется расположение групповых средних в верхнем и нижнем квадрантах разных периодов онтогенеза. Так, каноническая оценка потомства Саяна-53, Светлого-194, Светлого-768, Соболя-139 и Сома-746 не показала изменения знака рассматриваемых показателей. В то же время данной закономерности не наблюдалось по потомству Сыча-1618, которое также в значительной мере отличалось от средних показателей по группе.
Принимая во внимание представленные данные о структурных и стандартизированных коэффициентах, предполагаемая доля влияния генотипа хряка с исключением доли, определяемой корреляционными взаимодействиями, составила по уровню лейкоцитов 0,296, общего белка 0,147. По остальным переменным подобные коэффициенты приближались к нулю.
Таким образом, использование канонического дискриминантного анализа позволило выявить интерьерные признаки, по которым наблюдаются максимальные отличия с точки зрения многомерной природы их взаимодействий.
Выводы
Установлены интерьерные показатели, по которым наблюдается наибольшие отличия потомков разных хряков породы СМ-1 в разные периоды онтогенеза. В подсосный период потомки разных хряков отличались по активности АСТ, которая зависела от множества других факторов. В противоположность этому, поросята в возрасте 2 недели после отъема характеризовались отличиями, в меньшей степени диктуемыми существующими корреляционными взаимодействиями.
Использование активности АСТ и концентрации мочевины в крови поросят-сосунов мало информативно ввиду высокой коррелированности с множеством интерьерных показателей. Обнаружено влияние генотипа отца на уровень лейкоцитов (0,296) и общего белка (0,147) у поросят в постотъемный период.
