Каталог биохимических паспортов сортов овса посевного сибирской селекции

Автор: Любимова Анна Валерьевна, Еремин Дмитрий Иванович, Мамаева Виктория Сергеевна, Брагин Николай Александрович, Белоусов Сергей Александрович, Брагина Мария Владимировна, Кочнева Дарья Алексеевна, Таутекенова Азия Кайсаровна
Журнал: Вестник Красноярского государственного аграрного университета @vestnik-kgau
Рубрика: Агрономия
Статья в выпуске: 5, 2022 года.
Бесплатный доступ
Цель исследования - составление биохимических паспортов сортов овса посевного сибирской селекции с применением метода электрофореза проламинов для защиты авторских прав и использования в селекционном процессе. Материалом для исследования послужили сорта овса посевного селекции НИИСХ Северного Зауралья - филиала ТюмНЦ СО РАН. Установлено, что сорта Мегион и Отрада состоят из двух биотипов соотношением 2:1. Остальные сорта гомогенные по компонентному составу авенина. На основании новых данных о характере наследования авенинов составлены генетические формулы исследованных сортов: Мегион - AvnA2+9 B7 C5, Тюменский голозерный - AvnA2 B7 C3, Талисман - AvnA11 B4 C2, Отрада - AvnA10+11 B4 С8, Фома - AvnA11 B11 С8, Тоболяк - AvnA4 B8 С2. Максимальную частоту встречаемости у исследованных сортов имеют аллели А11, В4, В7 и С8. Эти аллели перспективны для дальнейшего изучения и использования в качестве маркеров высококонкурентных ассоциаций генов, важных в природно-климатических условиях Западной Сибири. В результате кластеризации установлено, что в регионе существует две группы сортов местной селекции, для каждой из которых характерно наличие своего адаптивного генного комплекса. Новый сорт Тоболяк не вошел ни в один из кластеров, что может говорить о наличии у него новых сочетаний генов, дающих преимущества на современном этапе селекции. Среднее генетическое разнообразие составляет 0,82, что свидетельствует об отсутствии процессов генетической эрозии и грамотно организованной селекционной работе с культурой в регионе.
Овес посевной, паспортизация, электрофорез, авенин, проламинкодирующие локусы, блоки компонентов, генетическое разнообразие
Короткий адрес: https://sciup.org/140294705
IDR: 140294705 | УДК: 581.19:631.52:633.112.1 | DOI: 10.36718/1819-4036-2022-5-73-83
Siberian oat varieties' biochemical passports catalog
The purpose of the study is to compile biochemical passports of Siberian oat varieties using the prolamin electrophoresis method for copyright protection and use in the breeding process. The material for the study was the cultivars of oats bred at the Scientific Research Institute of Agriculture of the Northern Trans-Urals - a branch of the Tyumen Scientific Center of the Siberian Branch of the Russian Academy of Sciences. It has been established that varieties Megion and Otrada consist of two biotypes with a ratio of 2:1. The remaining varieties are homogeneous in terms of the component composition of avenin. Based on new data on the nature of the inheritance of avenins, genetic formulas of the studied varieties were compiled: Megion - AvnA2+9 B7 C5, Tyumensky golozerny - AvnA2 B7 C3, Talisman - AvnA11 B4 C2, Otrada - AvnA10 + 11 B4 C8, Foma - AvnA11 B11 C8, Tobolyak - AvnA4 B8 C2. Alleles A11, B4, B7 and C8 have the maximum frequency of occurrence in the studied varieties. These alleles are promising for further study and use as markers of highly competitive gene associations that are important in the climatic conditions of Western Siberia. As a result of clustering, it was found that in the region there are two groups of varieties of local selection, each of which is characterized by the presence of its own adaptive gene complex. The new variety Tobolyak was not included in any of the clusters, which may indicate that it has new combinations of genes that provide advantages at the present stage of breeding. The average genetic diversity is 0.82, which indicates the absence of genetic erosion processes and well-organized breeding work with the crop in the region.
Текст научной статьи Каталог биохимических паспортов сортов овса посевного сибирской селекции
Введение. Эффективность внедрения современных сортов сельскохозяйственных культур в производство напрямую зависит от потенциала их использования, связанного в том числе с показателями сортовой принадлежности и чистоты.
В соответствии сГражданским кодексом Российской Федерации сорта зерновых являются интеллектуальной собственностью и охраняются патентом. При этом патент выдается только тем селекционным достижениям, которые отвечают критериям охраноспособности: обладают новизной, отличимостью, однородностью и стабильностью (ст. 1412-1413 ГК РФ) [1]. Наиболее распространенный метод сортовой идентификации и оценки сортовой чистоты – полевая апробация – основан на морфологических признаках растений. В настоящее время набирает популярность метод ДНК-фингерпринтинга, основанный на использовании молекулярных маркеров для генетической паспортизации сортов, гибридов и селекционных линий ряда сельскохозяйственных культур [2–4]. Однако для большого количества культур на сегодняшний день система идентификации с применением ДНК-маркеров еще не разработана. Решением данной проблемы может послужить использование для паспортизации биохимических маркеров.
У зерновых культур для этих целей широко применяются запасные спирторастворимые белки зерна – проламины. Метод электрофореза проламинов успешно применяется для паспортизации таких культур, как пшеница, ячмень, тритикале и овес [5–8]. Благодаря высокому уровню полиморфизма, практически каждый сорт или линия имеют свой уникальный спектр запасных спирторастворимых белков. Это позволяет успешно выявлять сортовую примесь и контролировать постоянство биотипного состава гетерогенных сортов. Кроме того, оценка частоты встречаемости аллелей проламин-кодирующих локусов позволяет судить о внутривидовом генетическом разнообразии сортов, а также делать выводы о динамике этого показателя во времени и пространстве.
В условиях Северного Зауралья одна из основных возделываемых культур – это овес. Первый сорт местной селекции Мегион был районирован в Тюменской области в 1993 г. На сегодняшний день в посевах региона доля сортов овса селекции НИИСХ Северного Зауралья – филиала ТюмНЦ СО РАН составляет 100 %. При
Таблица 1
Проанализированные сорта овса посевного
|
Сорт |
Разновидность |
Год включения в реестр |
Происхождение |
|
Мегион |
Mutica |
1993 |
Нарымский 943 × Пшебуй II |
|
Тюменский голозерный |
Inermis |
2000 |
Популяция производственного посева в Синьдзян-Уйгурском автономном округе КНР |
|
Талисман |
Mutica |
2002 |
Flamingsnova × Метис |
|
Отрада |
Mutica |
2014 |
(WW 170079 × Pc 39) × (Мутика 600 × Risto) |
|
Фома |
Mutica |
2015 |
(WW 170079 × Pc 39) × (Мутика 600 × Risto) |
|
Тоболяк |
Mutica |
2020 |
Таежник × Орион |
Для анализа от каждого сорта методом случайной выборки отбирали по 100 зерен. Электрофорез авенинов проводили в 13,2 % полиакриламидном геле по ранее описанной методике [10]. Для электрофоретического разделения использовали вертикальные электрофоретические камеры с размерами формируемых пластин 17,8×17,8×0,1 см (VE-20, Helicon, Россия). Электрофорез проходил в течение 4,0–4,5 ч при постоянном напряжении 500 V. Аллели авенин-кодирующих локусов идентифицировали в соответствии с каталогом, разработанным В.А. Пор-тянко [9] с дополнениями А.В. Любимовой [11]. В качестве стандарта использовали смесь муки зерновок овса посевного сортов Вятский голо- этом в семеноводстве новых сортов активно применяется метод электрофореза проламинов овса – авенинов. Авенины овса наследуются блоками, синтез которых контролируется тремя полиморфными независимыми авенин-кодирую-щими локусами – Avn A, Avn B и Avn C [9]. Благодаря достаточно высокому уровню полиморфизма, эти белки позволяют различать большинство сортов и линий, типировать гетерозиготы, а также оценивать динамику генетического разнообразия популяции.
Цель исследования – составление биохимических паспортов сортов овса посевного сибирской селекции с применением метода электрофореза проламинов для защиты авторских прав и использования в селекционном процессе.
Материалы и методы. Исследование проводили на базе лаборатории геномных исследований в растениеводстве Тюменского научного центра СО РАН в 2021 г. Материалом для исследования послужили сорта овса посевного селекции НИИСХ Северного Зауралья – филиала ТюмНЦ СО РАН (табл. 1).
зерный и Тюменский голозерный в соотношении 2:1 ( Avn A2 B4+7 C1+3 ) либо муку сорта овса посевного Astor ( Avn A2 B4 C2 ).
Генное разнообразие на локус (H) рассчитывали по формуле
И ·(1-∑KiPt}, где pi – популяционная частота i-го аллеля; k – количество аллелей локуса; n – объем выборки [12]. Для расчета среднего генного разнообразия (Н) усредняли количество аллелей на локус по всем локусам. Расчеты проводили с использованием программы Arlequin Ver 3.5.2.2 (Copyright 2015 L. Excoffier. CMPG, University of Berne).
Кластеризацию проводили методом Paired group, в качестве меры сходства использовали евклидовы расстояния, с проведением Bootstrap анализа для 100 повторностей. Расчеты проводили в программе PAST version 1.84 (Copyright Hammer and Harper 1999–2008).
Результаты и их обсуждение. В результате исследования установлено, что все проанализированные сорта отличаются друг от друга по компонентному составу авенина.
Сорт овса посевного Мегион гетерогенный и состоит из двух биотипов соотношением 2:1. Формула авенина сорта имеет вид Avn A2 + 9 B7 C5 . Блоки компонентов А9 и В7 идентифицированы нами в результате анализа характера наследования отдельных компонентов авенина в популяции гибридов F 2 . Более подробно эти исследования описаны в работе [11] (рис. 1).
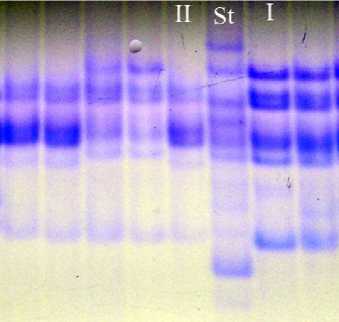
Рис. 1. Биохимический паспорт сорта овса посевного Мегион:
1 – эталонный спектр авенина; 2 – блоки компонентов авенина; I, II – биотипы; St – спектр-стандарт (Вятский голозерный +Тюменский голозерный)

Сорт овса посевного Тюменский голозерный гомогенный по компонентному составу авенина, от первого биотипа сорта Мегион он отличается только аллелем локуса Avn C. Формула авенина сорта Avn A2 B7 C3 (рис. 2).
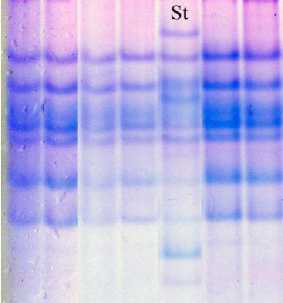
Рис. 2. Биохимический паспорт сорта овса посевного Тюменский голозерный: 1 – эталонный спектр авенина; 2 – блоки компонентов авенина;
St – спектр-стандарт (Вятский голозерный + Тюменский голозерный)
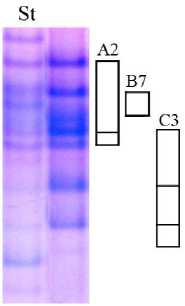
В результате исследования проламинов сорта овса Талисман выявлен один тип спектра. В наших ранних исследованиях в результате идентификации аллелей авенин-кодирующих локусов была составлена формула этого сорта
Avn A4 B4 C2. При этом установлено, что блоки компонентов, синтез которых контролируется аллелями локусов Avn A и Avn B сорта Талисман, идентичны таковым у второго биотипа сорта Отрада (ранее идентифицированная форму- ла сорта Avn A10+4 B4 С1). Однако в результате оценки характера наследования компонентов авенина в популяции F2 от скрещивания сорта Отрада с сортами Тюменский голозерный и Мегион было установлено, что в спектрах второго биотипа сорта Отрада присутствует не блок компонентов А4, а новый, ранее не описанный блок, которому присвоен номер А11. Следовательно, этот же блок компонентов характерен и для сорта Талисман.
Таким образом, биохимическая формула сорта Талисман имеет вид Avn A11 B4 C2 (рис. 3).
St
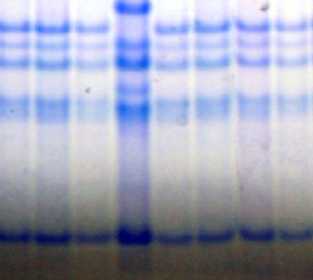
Рис. 3. Биохимический паспорт сорта овса посевного Талисман: 1 – эталонный спектр авенина; 2 – блоки компонентов авенина; St – спектр-стандарт (Astor)

Для сорта Отрада, помимо идентификации новых блоков компонентов, контролируемых аллелями локуса Avn А, был уточнен блок компонентов, контролируемый аллелем локуса Avn C. В данном случае использование в качестве стандарта сорта Astor с аллелем С2 локуса Avn C привело к ошибке в идентификации. Анализ с использованием в качестве стандарта сорта с аллелем C1 показал, что для сорта Отрада характерен новый блок, отличающийся от С2 и С1 меньшей подвижностью компонентов в зоне быстрых проламинов (отмечены красными маркерами на рисунке 4). Формула авенина сорта Отрада Avn A10+11 B4 С8 [11].
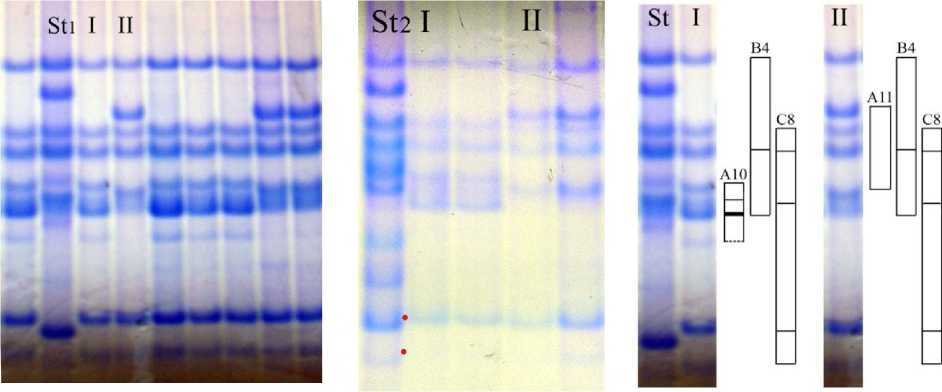
1 2
Рис. 4. Биохимический паспорт сорта овса посевного Отрада:
1 – эталонный спектр авенина; 2 – блоки компонентов авенина; I, II – биотипы; St1 – спектр-стандарт (Astor); St2 – спектр-стандарт (Вятский голозерный + Тюменский голозерный)
В результате анализа характера наследования компонентов авенина сорта Фома в популяциях F2 от скрещивания сорта Фома с сортами Отрада, Мегион и Тюменский голозерный установлено, что по аллелям локусов Avn A и Avn С сорт Фома идентичен второму биотипу сорта Отрада. Анализ популяции F2 позволил описать новый блок компонентов, контролируемый аллелем локуса Avn В, – В11 (рис. 5). Формула сорта Фома Avn A11 B11 С8.
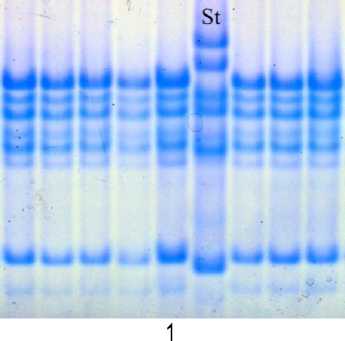
Рис. 5. Биохимический паспорт сорта овса посевного Фома:
1 – эталонный спектр авенина; 2 – блоки компонентов авенина; St – спектр-стандарт (Astor)

Сорт овса Тоболяк гомогенный по компонентному составу авенина. В результате идентификации блоков компонентов авенина для данного сорта был описан новый блок, синтез которого контролируется аллелем локуса Avn В, – В8 (рис. 6). Формула авенина сорта Тоболяк: Avn A4 B8 С2.
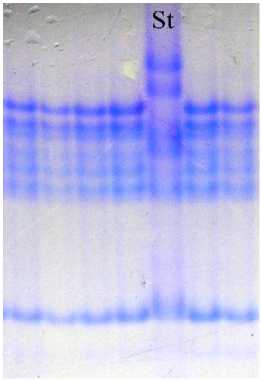
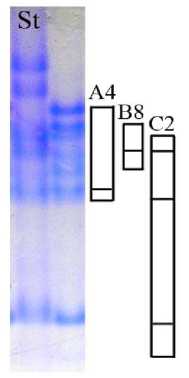
Рис. 6. Биохимический паспорт сорта овса посевного Тоболяк: 1 – эталонный спектр авенина; 2 – блоки компонентов авенина; St – спектр-стандарт (Astor)
На основании данных об аллельном составе сортов была проведена кластеризация. При этом каждый биотип рассматривался как отдельный образец. Как видно из дендрограммы, представленной на рисунке 7, большая часть образцов разделилась на два крупных кластера, сорт Тоболяк занял отдельностоящее положение. В первый кластер вошли биотипы сорта Мегион и сорт Тюменский голозерный, во второй – сорта Фома, Отрада и Талисман.
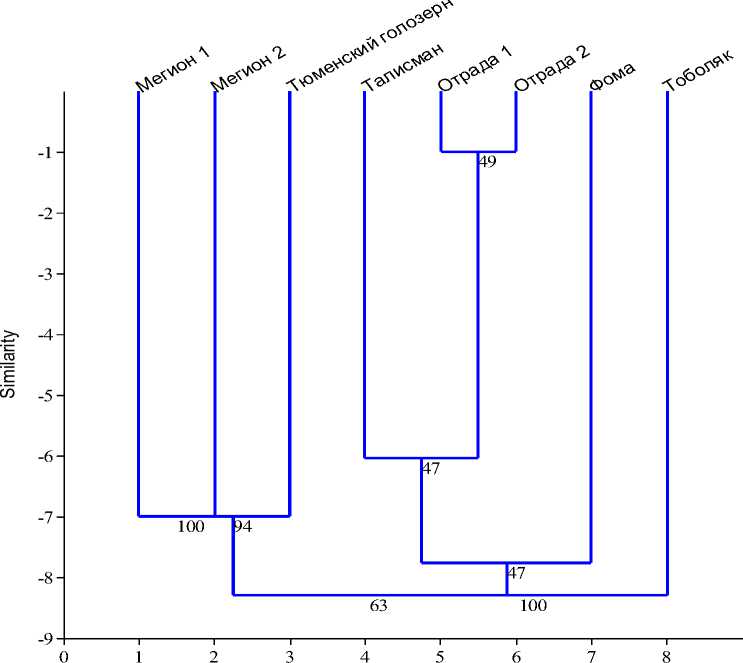
Рис. 7. Кластеризация сортов овса посевного сибирской селекции по аллельному составу авенин-кодирующих локусов (на ветвях указаны значения бутстеп-идндексов)
Объединение сортов в кластеры может свидетельствовать о наличии у них сходных селекционно ценных и адаптивно значимых признаков. Адаптивный характер полиморфизма запасных спирторастворимых белков показан на большом количестве примеров, в т. ч. для таких культур, как пшеница, ячмень и овес. Под действием отбора в определенных природноклиматических условиях формируются генотипы, несущие устойчивые сочетания генов – адаптивные генные комплексы. Связь проламинов с такими комплексами дает возможность выделять генотипы, наиболее приспособленные к конкретным условиям среды, основываясь на данных об аллельном составе проламин-кодирующих локусов. Кроме того, повышенная частота встречаемости конкретных аллелей АКЛ в сортах селекции определенного селекционного центра может свидетельствовать об их сцепленности с хозяйственно значимыми признаками. В результате в ходе селекционной работы при отборе генотипов с наилучшим сочетанием генов в популяции сортов происходит увеличение частоты встречаемости аллелей АКЛ, маркирующих эти сочетания [13, 14].
Таким образом, по результатам кластеризации можно сделать вывод о том, что в Тюменской области существует две группы сортов местной селекции, для каждой из которых характерно наличие своего адаптивного генного комплекса. Сорт Тоболяк создан недавно и не вошел ни в один из кластеров, указывает на наличие у него новых сочетаний генов, дающих преимущества на современном этапе селекции.
Согласно таблице 2, наибольшую частоту встречаемости в популяции сортов овса сибирской селекции имеют аллели А11, В4, В7 и С8. Эти аллели могут выступать в роли маркеров высококонкурентных ассоциаций генов, важных в природно-климатических условиях Западной Сибири.
Частота встречаемости аллелей авенин-кодирующих локусов и генетическое разнообразие (Н) исследованных сортов овса
Таблица 2
|
Локус |
Аллель |
Частота встречаемости, % |
Н±h |
Среднее генное разнообразие ( Н ) |
|
Avn A |
2 |
25,0 |
0,86±0,108 |
0,82 |
|
4 |
12,5 |
|||
|
9 |
12,5 |
|||
|
10 |
12,5 |
|||
|
11 |
37,5 |
|||
|
Avn B |
4 |
37,5 |
0,78±0,113 |
|
|
7 |
37,5 |
|||
|
8 |
12,5 |
|||
|
11 |
12,5 |
|||
|
Avn С |
2 |
25,0 |
0,82±0,101 |
|
|
3 |
12,5 |
|||
|
5 |
25,0 |
|||
|
8 |
37,5 |
Необходимо отметить, что преобладание определенных аллелей АКЛ в популяции сортов свидетельствует о процессе сужения внутривидового генетического разнообразия. Это связано с введением в селекцию одних и тех же генотипов, характеризующихся наличием конкретных хозяйственно ценных признаков. Генетическая эрозия приводит к снижению устойчивости популяции сортов к различным фитопатогенам, а также отрицательно сказывается на их способности к адаптации [15, 16].
Расчет величины генетического разнообразия показал, что по всем АКЛ этот показатель составляет более 0,70, что свидетельствует об отсутствии процессов генетической эрозии в популяции сортов овса посевного и говорит о грамотно организованной селекционной работе с культурой (см. табл. 2).
Заключение
-
1. С применением метода электрофореза проламинов составлены биохимические паспорта для сортов овса посевного сибирской селекции. Сорта Мегион и Отрада являются гетерогенными и состоят из двух биотипов соотношением 2:1. Генетические формулы авенина исследованных сортов имеют вид: Мегион – Avn A2+9 B7 C5, Тюменский голозерный – Avn A2 B7 C3, Талисман – Avn A11 B4 C2, Отрада – Avn A10+11 B4 С8, Фома – Avn A11 B11 С8, Тоболяк – Avn A4 B8 С2.
-
2. Сорта овса посевного по аллельному составу авенин-кодирующих локусов разделились
-
3. Сорта селекции НИИСХ Северного Зауралья – филиала ТюмНЦ СО РАН характеризуются высоким генетическим разнообразием (0,82), что связано с введением в селекционные программы генетически разнообразного материала.
на две группы, каждая из которых, вероятно, характеризуется определенным устойчивым сочетанием генов, дающих несущим их генотипам преимущества в природно-климатических условиях региона. Сорт Тоболяк занял отдельное положение, что свидетельствует о наличии в его генотипе новых сочетаний генов, ценных на современном этапе селекции.
Список литературы Каталог биохимических паспортов сортов овса посевного сибирской селекции
- Гражданский кодекс Российской Федерации (часть четвертая) от 18.12.2006 № 230-Ф3 (ред. от 11.06.2021) (с изм. и доп., вступ. в силу с 01.01.2022). М., 2022.
- Идентификация и паспортизация сортов кормовых трав (клевера лугового, люцерны изменчивой, посевной и хмелевидной) на основе ДНК-маркеров / И.А. Клименко [и др.]. М.: Угреша Т, 2020. 35 с. DOI: 10.33814/978-5-6043194-9-9.
- Шамустакимова А.О., Мавлютов Ю.М., Клименко И.А. Применение SRAP-маркеров для ДНК-идентификации российских сортов люцерны // Генетика. 2021. Т. 57, № 5. С. 536543. DOI: 10.31857/S0016675821050118.
- SSR marker-based DNA fingerprinting of Sub1 introgressed lines in the background of traditional rice varieties of Assam India / A. Gau-tam [et al.] // Indian Journal of Biochemistry and Biophysics. Vol. 59(3). March 2022. P. 350-356.
- Ярова Э.Т., Тоболова Г.В. Использование электрофореза запасных белков для селекции яровой тритикале // Вестник КрасГАУ. 2021. № 5 (170). С. 96-102. DOI: 10.36718/1819-4036-2021-5-96-102.
- Genetic diversity of gliadin-coding alleles in bread wheat (Triticum aestivum L.) from Northern Kazakhstan / M. Utebayev [et al.] // Peer J. 2019. 7:e7082. DOI: 10.7717/peerj. 7082.
- The development of the educational and applied database in MS Access for selection and genetic research / L.N. Shevtsova [et al.] // IOP Conference Series: Earth and Environmental Science: conference proceedings, Krasnoyarsk, Russia, 13-14 ноября 2019 года / Krasnoyarsk Science and Technology City Hall of the Russian Union of Scientific and Engineering Associations. Krasnoyarsk, Russia: Institute of Physics and IOP Publishing Limited, 2020. T. 421. P. 42014. DOI: 10.1088/1755-1315/ 421/4/ 042014.
- Ostapenko A.V., Tobolova G.V. Polymorphism of avenin species A. satva L., A. byzantina C. Koch. and A. strigosa Schreb // Plant Genetics, Genomics, Bioinformatics and Biotechnology : The 3rd International Conference. Abstract book, Novosibirsk, 17-21 июня 2015 года. Novosibirsk: Федеральный исследовательский центр Институт цитологии и генетики Сибирского отделения Российской академии наук, 2015. P. 39.
- Генетический контроль авенинов и принципы их классификации / В.А. Портянко [и др.] // Генетика. 1987. № 23 (5). С. 845853.
- Динамика генетического разнообразия сортов овса в Тюменской области по авенин-кодирующим локусам / А.В. Любимова [и др.] // Вавиловский журнал генетики и селекции. 2020. Т. 24 (2). С. 123-130. DOI: 10.18699/VJ20.607.
- Любимова А.В. Изучение характера наследования компонентов авенина у гибридов F2 от скрещивания овса посевного сибирской селекции // Аграрный вестник Урала. 2022. № 02 (217). С. 48-59. DOI: 10.32417/ 1997-4868-2022-217-02-48-59.
- Nei M., Li W. Mathematical model for studying genetic variation in terms of restriction endo-nucleases // Proc. Natl. Acad. Sci. USA, 1979, vol. 76. pp. 5269-5273.
- Генетическое разнообразие современных российских сортов яровой и озимой твердой пшеницы по глиадинкодирующим локусам / А.М. Кудрявцев [и др.] // Генетика. 2014. Т. 50, № 5. С. 554-559.
- Взаимосвязь между составом глиадинов, морфологией колоса и качеством зерна у яровой пшеницы / А.В. Фисенко [и др.] // Достижения науки и техники АПК. 2018. Т. 32, № 9. С. 24-27. DOI: 10.24411/02352451-2018-10906.
- Гончаренко А.А. Экологическая устойчивость сортов зерновых культур и задачи селекции // Зерновое хозяйство России. 2016. № 3. С. 31-37.
- Спектры проламинов в агроэкологической оценке коллекционного материала ячменя / Н.В. Зобова [и др.] // Достижения науки и техники АПК. 2018. № 32 (5). С. 45-47. DOI: 10.24411/0235-2451-2018-10511.
