Конроль развития плода у самок черноморской афалины (Tursiops truncatus ponticus Barabash,1940) во второй половине беременности
 во второй половине беременности")
Автор: Семенов В.А., Родин И.А., Околелова А.И., Винокурова Д.П., Яковец М.Г., Шантыз А.Ю.
Журнал: Вестник Красноярского государственного аграрного университета @vestnik-kgau
Рубрика: Ветеринария и зоотехния
Статья в выпуске: 10, 2019 года.
Бесплатный доступ
Цель исследований - выявление беременности у самок черноморской афалины с помощью ультразвуковой диагностики (УЗИ) и изучение динамики линейных размеров головы и грудной клетки плода во второй ее половине, а также показателей крови беременных самок в зависимости от сроков беременности в случаях благополучного и неблагополучного родоразрешения. В период с 2004 по 2017 год проводились исследования в различных дельфинариях на побережье Чёрного моря. Было выявлено 30 случаев беременности черноморских афалин и проведен ряд исследований. На протяжении всего периода исследований имели случаи благополучного и неблагополучного родоразрешения, что послужило причиной формирования двух групп рожденных. В первую группу включили благополучно родившихся детенышей, вовторую группу - мертворожденных или погибших в первый день после рождения. Достоверных отличий между размерами головы и грудной клетки на одном и том же месяце беременности у этих двух групп детенышей не отмечалось. Метод ультразвуковой диагностики присутствия беременности у самок дельфинов в сравнении с другими методами хорошо зарекомендовал себя...
Ультразвуковое исследование, беременность, черноморская афалина, гематологические показатели
Короткий адрес: https://sciup.org/140245587
IDR: 140245587 | УДК: 619:618.2]:639.2:599.537
Fetal development control in female of black sea bottlenose dolphins (Tursiops truncatus ponticus Barabash, 1940) in the second half of pregnancy
The aim of the study was to identify pregnancy in females of black sea bottlenose dolphin by ultrasound diagnosis (UD) and to study the dynamics of linear dimensions of the head and chest of the fetus in its second half, as well as blood parameters of pregnant females, depending on its timing in cases both of safe and unsuccessful delivery. In the period from 2004 to 2017, the studies were conducted in various dolphinariums on the Black Sea coast. 30 cases of pregnancy of black sea bottlenose dolphins were detected and a number of studies were conducted. Throughout the period of studies, there were cases of successful and unsuccessful delivery, which caused the formation of two groups of newborns. The first group included safely born cubs, the second group-stillborn or dead on the first day after birth. There were no significant differences between the size of head and chest in the same month of pregnancy in these two groups of bottlenose calves. The method of ultrasonic diagnostics of pregnancy in female dolphins in comparison with other methods was well established...
Текст научной статьи Конроль развития плода у самок черноморской афалины (Tursiops truncatus ponticus Barabash,1940) во второй половине беременности
Введение . Период беременности у афалин длится приблизительно 12 ± 1 месяцев. К примеру, у индийской афалины ( T.t. Aduncus) с известными датами зачатия определяют срок 370 ± 11 дней [1]. Период лактации у афалин может продолжаться до 2 лет и даже более (у диких животных) [2]. Большинство самок впервые производят на свет детенышей в возрасте от 7 до 10 лет [3]. Полный репродуктивный цикл, или промежуток времени между родами, составляет 3–4 года. Продолжительность эстрального цикла у T. truncatus составляет от 21 до 42 дней [4– 6].
Все китообразные являются одноплодными животными, у которых случаи многоплодия очень редки и колеблются от 0,002 % у финвала (Balaenoptera physalus) до 0,5 % у белухи (Delphinapterus leucas). Одноплодность у китообразных связывают с необходимостью высокого уровня развития новорождённых и рассматривают как положительный эволюционно приобретенный признак, а редкие случаи многоплодия относят к атавистическим признакам [7]. По имеющимся данным [8] о многочисленных случаях родов у самок черноморской афалины в неволе, все они завершались рождением одного детеныша. В литературе упоминается всего два случая двуплодной беременности. Ни одна их них не имела благополучного исхода. В одном случае близнецы родились мёртвыми [9]; в другом – у самки произошли преждевременные роды, и спустя четыре месяца после её гибели, при вскрытии, у неё был обнаружен доношенный мацерированный плод [10].
Единственный, хорошо развитый детеныш рождается очень крупным – от 1/4 до 1/2 длины тела матери. Изредка в одной самке находят несколько зародышей. У синего кита однажды обнаружили семь, а у финвала и сейвала – по шесть зародышей. Это свидетельство былого многоплодия предков китообразных. Лишние зародыши обычно рассасываются, и только очень редко могут родиться двойни [11]. Гибель одного из эмбрионов у людей может привести к серьезным нарушениям здоровья второго или даже его гибели. По статистике, в случае смерти одного из эмбрионов в первой трети беременности вероятность нормального развития и рождения второго достигает 90 %. В этом случае происходит полное рассасывание эмбриона или его размягчение. Если смерть одного плода при двойне произошла на более поздних сроках, то у второго может развиться тяжелое поражение центральной нервной системы, внутренних органов или гибель [12].
Действие различных факторов окружающей среды на репродуктивный цикл свидетельствует о том, что процесс размножения находится под контролем нервных стимулов, передающихся в мозг. Основная часть передачи и преобразования этих стимулов, по-видимому, происходит в гипоталамусе и связанных с ним ядрах, нейроны которых секретируют кринотропные гормоны в портальную систему гипофиза [2]. Эти гормоны контролируют переднюю долю гипофиза, или аденогипофиза. К числу таких нейрогормонов относится гонадотропин-рилизинг-гормон (ГнРГ), который имеет первостепенное значение для регуляции эндокринных аспектов размножения млекопитающих. Активация ГнРГ-рецепторов в передней доле гипофиза вызывает секрецию в кровоток лютеинизирующего гормона (ЛГ) и фолликулостимулирующего гормона (ФСГ). По мере роста и созревания фолликула вырабатывается эстроген за счет паракринного взаимодействия между клетками внутренней оболочки (theca interna) фолликула и зернистого слоя (гранулозы), выстилающего фолликул. Вначале рост синтеза эстрогена ингибирует и ЛГ-, и ФСГ-секрецию в гипофизе. По мере приближения созревающего фолликула к преову-ляционной стадии эстрогены, достигшие максимального уровня секреции, начинают оказывать положительное обратное действие на частоту и амплитуду выбросов ГнРГ, что приводит, в свою очередь, к преовуляторному выбросу ЛГ. ЛГ стимулирует образование в фолликуле низкомолекулярного гликопротеина, так называемого ингибина. Ингибин не только подавляет секрецию ФСГ, но и увеличивает чувствительность клеток theca interna преовуляционного фолликула к ЛГ [13]. Данное сочетание увеличения числа рецепторов к ЛГ и повышения чувствительности к ЛГ обеспечивает соответствующую реакцию на выброс ЛГ и овуляцию.
После овуляции зернистые и обкладочные клетки преобразуются в большие прогестерон-секретирующие и малые лютеиновые клетки соответственно [14]. Прогестерон и в меньшей степени эстроген, продуцируемые желтым телом, подавляют лютеинизирующий и фолликулостимулирующий гормоны, снижая уровень гонадотропин-рилизинг-гормона, высвобождаемого из гипоталамуса.
У содержащихся в Atlantis Marine Park (Западная Австралия) самок афалин использовалось ультразвуковое обследование в В-режиме для диагностики беременности и измерения растущего плода [15]. Обследование проводили примерно с месячным интервалом, используя ультразвуковой аппарат AlokaSSD260 (компании Aloka, Токио, Япония) с линейным датчиком частотой 33-5 МГц (AlokaUST-5021). Преобразователь был заключен в тонкий пластиковый пакет (для защиты от контакта с соленой водой) и снабжен большим количеством геля, чтобы обеспечить хороший контакт с объектом. Проводились замеры поперечного сечения грудной клетки на уровне 2-го или 3-го межреберья, диаметра черепа, ширины ребер и шейных позвонков, а также длины рострума. Из-за непродолжительности процесса визуализации были получены надлежащие изображения грудной клетки и диаметра черепа (затылочно-лобной оси), изображения остальных структур плода не дали надежных результатов. Определение размеров структур плода проводились путем сопоставления их с масштабной шкалой на экране ультразвукового аппарата. В результате исследований выявлена линейная зависимость увеличения параметров развивающегося плода. Сердцебиение плода регулярно визуализировалось и указывало на его жизнеспособность.
Особый интерес представляют исследования динамики роста головы и грудной клетки плода у самок с успешным родоразрешением с одной стороны, и неудачными родами – с другой.
Цель исследований. Выявление беременности у самок черноморской афалины (Tursiops truncatus ponticus Barabash,1940) с помощью ультразвукового исследования и изучение динамики линейных размеров головы и грудной клетки плода во второй половине беременности, а также анализов крови беременных самок, в зависимости от её сроков в случаях благополучного и неблагополучного родоразрешения.
Материалы и методы. Исследования проводились с 2004 по 2017 г. в различных дельфинариях на побережье Чёрного моря. Нами было выявлено 30 случаев беременности черноморских афалин у 15 самок в возрасте от 6,5 до 20 лет и проведено 203 исследования. В связи с отсутствием опыта по проведению ультразвукового обследования черноморских дельфинов на начальных этапах изучения данного вопроса не все исследования оказались корректными и достоверными, поэтому лишь 144 из них вошли в данную работу и подверглись биометрической обработке. Длина животных варьировала от 240 до 275 см, а вес в период отсутствия беременности или в начальной её стадии колебался от 172 до 283 кг.
Беременные животные во время обследования не вынимались из воды, а обследовались у бортика бассейна в боковом лежачем положении, которое позволяло в любой момент приподнять голову над уровнем воды и сделать беспрепятственно акт выдоха-вдоха. Кроме того, дельфины очень хорошо слышат ультразвуки, издаваемые ими же самими в воде, а также их отражения, которые легко проникают через тело этих животных и достигают внутреннего уха [7]. Поэтому требуется некоторое время, возможно, несколько сеансов с пищевым подкреплением, чтобы дельфин привык к ультразвуку, излучаемому датчиком ультразвукового аппарата, и при этом не проявлял бы беспокойства. Данные исследования были сконцентрированы на выявлении беременности с помощью ультразвукового обследования самок и изучении динамики дорсовентральных размеров головы и грудной клетки плода в сагиттальной плоскости во второй половине беременности. Голова измерялась по контурам костей, а грудная клетка – с учётом мягких тканей на уровне сердца. Срок беременности устанавливался ретроспективно от даты родов.
Все 30 случаев беременностей были разделены на две группы:
-
1) закончившиеся удачным родоразрешени-ем;
-
2) закончившиеся неудачным родоразреше-нием, характеризующимся появлением на свет мертворожденных детёнышей или погибших в первый день после рождения.
Биометрическая обработка промеров плода, а также результатов анализов крови самок в течение беременности проводилась как в целом во всех случаях исследований, так и по отдельности в каждой группе. Ультразвуковое обследование беременных самок осуществлялось с помощью аппарата SonoSite180 производства США с глубиной проникновения ультразвуковых волн до 22 см и конвексным преобразователем C60/5 – 2 MHz. Кровь для исследований брали из хвостовых вен – Superficial fluke vv . При этом клинический анализ крови выполнялся на автоматическом анализаторе «Cell-Dyn» (Abbott США) с использованием тест-систем фирмы Abbott США; биохимические исследования проводились на автоматическом анализаторе «ARCHITECT C-8000» (Abbott США) с использованием тест-систем фирмы Abbott США; концентрация прогестерона в крови определялась на автоматическом анализаторе «Architect i2000» (Abbott США) с использованием тест-систем фирмы Abbott США.
Результаты исследований и их обсуждение. Беременность во всех случаях в наших исследованиях завершалась родоразрешением через естественные родовые пути. Уровень плазменного прогестерона в период беременности колебался от 1,70 до 65,1 нг/мл и был в среднем равен 24,54 ± 1,882 нг/мл (число исследований равнялось 66), в то время как в отсутствии беременности он был достоверно ниже (P < 0,001), варьируя от 0 до 17,1 нг/мл, и был в среднем равен 2,14 ± 0,392 нг/мл (число исследований – 74). На протяжении второй половины беременности фиксировалось сердцебиение плода у всех беременных самок от 120 ударов в минуту на 6-м месяце и 117 ударов в минуту на
7-м месяце до 96 ударов в минуту в начале 12-го месяца, а затем до 76 в конце 12-го месяца за 5 дней до рождения.
Начиная с 6 до 9-го месяца беременности включительно размеры головы и грудной клетки плода каждый месяц достоверно (от P <0,05 до P <0,001) увеличивались в сравнении с предыдущим месяцем (табл. 1).
Таблица 1
Динамика изменений дорсовентральных размеров головы и грудной клетки (X ± m, см) плода в период беременности у самок черноморской афалины
|
Месяц беременности |
Дорсовентральные размеры головы (N=15, n=30) |
Дорсовентральные размеры грудной клетки (N=15, n=30) |
||
|
Количество исследований |
X ± m, см |
Количество исследований |
X ± m, см |
|
|
7-й |
10 |
6,5 ± 0,26 ** |
10 |
7,6 ± 0,21 ** |
|
8-й |
11 |
8,7 ± 0,50 * |
12 |
9,9 ± 0,45 *** |
|
9-й |
7 |
10,7 ± 0,12 * |
7 |
11,6 ± 0,29 ** |
|
10-й |
10 |
11,3 ± 0,24 |
10 |
12,9 ± 0,20 ** |
|
11-й |
11 |
12,0 ± 0,21 ** |
11 |
14,5 ±0,12 *** |
|
12-й |
8 |
14,1 ± 0,42 * |
5 |
14,8 ± 0,25 |
|
Итого |
57 |
55 |
||
Примечание. Здесь и далее : X – средняя арифметическая; m – стандартная ошибка для выборочной доли; N – число обследованных особей; n – число исследованных беременностей.
Достоверность различий между данным сроком беременности и предыдущим: * – P <0,05; ** – P <0,01; *** – P < 0,001
На 7-м месяце было выявлено увеличение грудной клетки плода до 7,6± 0,21 см, а головы – до 6,5 ±0,26 см (рис. 1).
На 8-м месяце диаметр грудной клетки увеличился до 9,9±0,45 см, а головы – до 8,7±0,50 см. На 9-м месяце рост грудной клетки достигает в среднем 11,6 ±0,29 см, а головы – 10,7±0,12 см.
На 10-м месяце грудная клетка плода продолжает достоверно (P <0,01) увеличиваться и достигает 12,9±0,20 см, а голова – 11,3±0,24 см.
На 11-м месяце диаметр грудной клетки (рис. 2) и головы также достоверно больше (P <0,001 и P <0,01), чем на 10-м месяце, и в среднем равен соответственно 14,5 ±0,12 и 12,0 ±0,21 см.
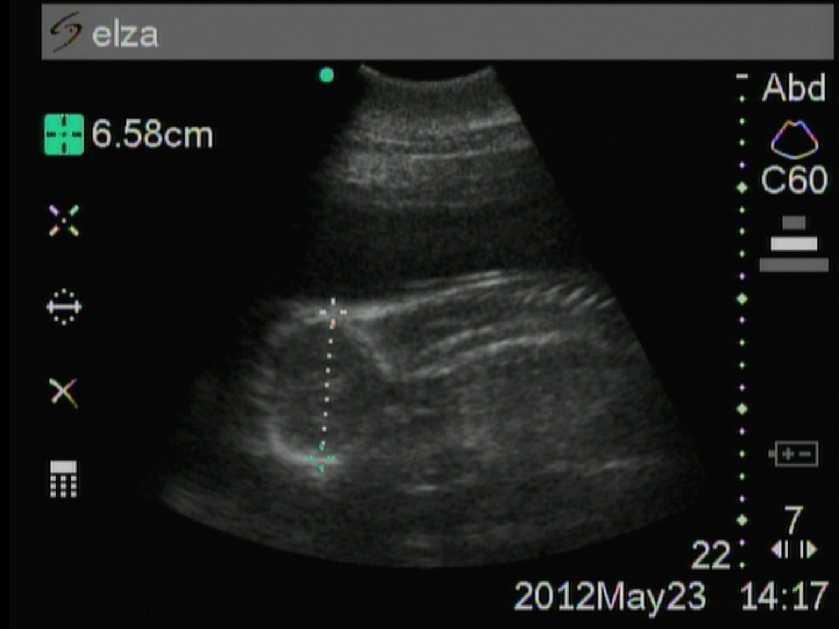
Рис. 1. Седьмой месяц беременности (голова)
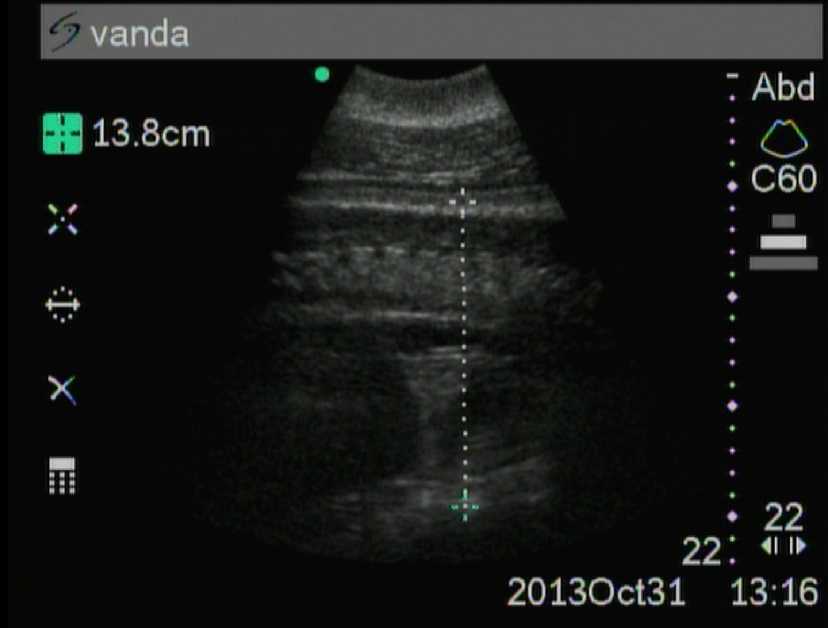
Рис. 2. Одиннадцатый месяц беременности (грудная клетка)
Далее на последнем, 12-м месяце беременности (рис. 3 и 4) голова плода достоверно (P <0,01) увеличивалась, что нельзя сказать о грудной клетке. Размер грудной клетки изменился незначительно (14,8 ± 0,25 см), в то время как размер головы увеличивается до 14,1 ± 0,42 см и почти достигает показателей грудной клетки.
Величина диаметра грудной клетки опережает диаметр головы с 5-го и до 12-го месяца. При этом среди достоверных изменений размеров головы плода отмечалось наибольшее их увеличение в течение всего срока беременности на 8-м месяце, а именно на 33,8 % по сравнению с предыдущим, восьмым месяцем.
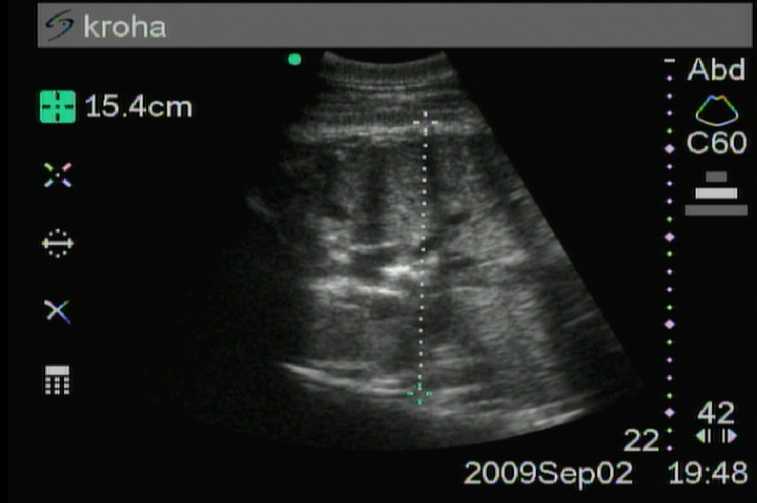
Рис. 3. Двенадцатый месяц беременности (грудная клетка )
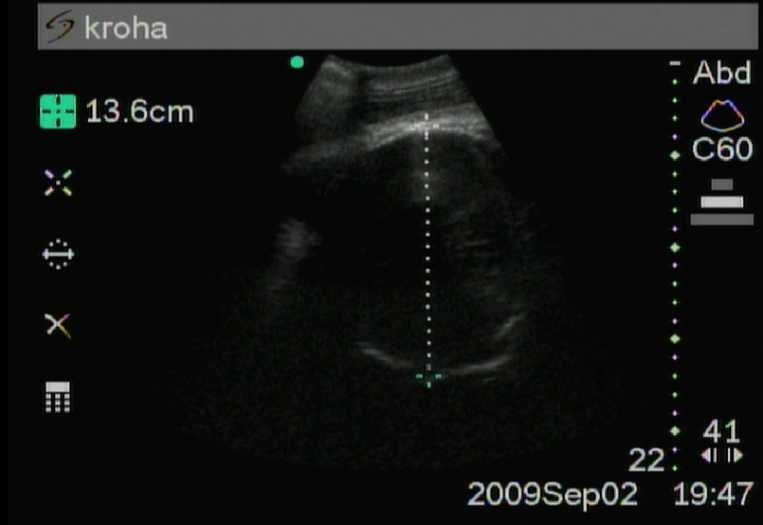
Рис. 4. Двенадцатый месяц беременности (голова)
На протяжении всего периода исследований имели случаи благополучного и неблагополучного родоразрешения, сформировавшие две группы рождённых. Особи первой группы благополучно родились и живут длительное время. Ко второй группе отнесли случаи, когда детёныши рождались мертвыми или погибали в первый день после рождения. Особый интерес составило не только сравнение промеров головы и грудной клетки детёнышей двух групп в пери- од беременности, но и показателей крови их матерей в этот момент жизни. Из таблиц 2 и 3 видно, что достоверные отличия между размерами головы и грудной клетки на одном и том же месяце беременности у этих двух групп детенышей афалины не отмечались.
В то же время, как видно из таблицы 4, различия между определёнными показателями крови беременных самок оказались существенными.
Таблица 2
Динамика изменений дорсовентральных размеров головы (X ± m, см) плода в период беременности у самок черноморской афалины
|
Месяц беремен-ности |
Нормальные родоразре-шения (N=11, n=20) |
Патологические родоразрешения (N=7, n=10) |
Все родоразрешения (N=15, n=30) |
|||
|
Количество исследований |
X ± m, см |
Количество исследований |
X ± m, см |
Количество исследований |
X ± m, см |
|
|
7-й |
9 |
6,5 ± 0,29 |
1 |
6,4 |
10 |
6,5 ± 0,26 |
|
8-й |
8 |
8,1 ± 0,43 |
3 |
10,3 ± 1,01 |
11 |
8,7 ± 0,50 |
|
9-й |
6 |
10,7 ± 0,13 |
1 |
11 |
7 |
10,7 ± 0,12 |
|
10-й |
8 |
11,3 ± 0,30 |
2 |
11,1 ± 0,10 |
10 |
11,3 ± 0,24 |
|
11-й |
6 |
12,1 ± 0,33 |
5 |
12,0 ± 0,27 |
11 |
12,0 ± 0,21 |
|
12-й |
5 |
13,4 ± 0,25 |
3 |
15,3 ± 0,46 |
8 |
14,1 ± 0,42 |
|
Итого |
42 |
15 |
57 |
|||
Таблица 3
Динамика изменений дорсовентральных размеров грудной клетки (X ± m, см) плода в период беременности у самок черноморской афалины
|
Месяц беременности |
Нормальные родоразреше-ния (N=11, n=20) |
Патологические родоразрешения (N=7, n=10) |
Все родоразрешения (N=15, n=30) |
|||
|
Количество исследований |
X ± m, см |
Количество исследований |
X ± m, см |
Количество исследований |
X ± m, см |
|
|
7-й |
9 |
7,9 ± 0,21 |
1 |
6,8 |
10 |
7,6 ± 0,21 |
|
8-й |
8 |
9,6 ± 0,55 |
4 |
10,5 ± 0,75 |
12 |
9,9 ± 0,45 |
|
9-й |
6 |
11,7 ± 0,34 |
1 |
11,3 |
7 |
11,6 ± 0,29 |
|
10-й |
8 |
13,0 ± 0,20 |
2 |
12,3 ± 0,50 |
10 |
12,9 ± 0,20 |
|
11-й |
7 |
14,5 ± 0,14 |
4 |
14,5 ± 0,26 |
11 |
14,5 ± 0,12 |
|
12-й |
4 |
14,8 ± 0,32 |
1 |
15 |
5 |
14,8 ± 0,25 |
|
Итого |
42 |
13 |
55 |
|||
Таблица 4
Гематологические показатели (X ± m, см) в период беременности у самок черноморской афалины
|
Показатель |
Нормальные родоразрешения (N=5, n=14) |
Патологические родораз-решения (N=4, n=7) |
Все родоразрешения (N=15, n=30) |
|||
|
Количество исследований |
X ± m, см |
Количество исследований |
X ± m, см |
Количество исследований |
X ± m, см |
|
|
Возраст, лет |
20 |
16,6 ± 1,20 |
10 |
9,8 ± 0,87 ** |
30 |
14,3 ± 1,03 |
|
Гемоглобин, г/л |
70 |
172,7 ± 1,36 |
33 |
162,3 ± 1,95 ** |
103 |
169,3 ± 1,21 |
|
Эритроциты, х 1012/л |
84 |
3,8 ± 0,05 |
45 |
3,7 ± 0,06 |
129 |
3,8 ± 0,04 |
|
Лейкоциты, х 109/л |
84 |
7,7 ± 0,20 |
45 |
7,3 ± 0,33 |
129 |
7,6 ± 0,17 |
|
Палочкоядерные нейтрофилы, % |
84 |
1,4 ± 0,12 |
45 |
2,2 ± 0,27 * |
129 |
1,7 ± 0,13 |
|
Сегментоядерные нейтрофилы, % |
84 |
54,1 ± 1,34 |
45 |
55,3 ± 1,30 |
129 |
54,5 ± 0.98 |
|
Эозинофилы, % |
84 |
19,3 ± 0,84 |
45 |
17,9 ± 1,21 |
129 |
18,8 ± 0,69 |
|
Лимфоциты, % |
84 |
21,6 ± 1,05 |
45 |
21,9 ± 1,45 |
129 |
21,7 ± 0,85 |
|
Моноциты, % |
84 |
3,3 ± 0,21 |
45 |
3,1 ± 0,33 |
129 |
3,2 ± 0,18 |
|
СОЗ, мм/ч |
75 |
2,9 ± 0,31 |
43 |
3,1 ± 0,57 |
129 |
3,0 ± 0,29 |
|
Прогестерон, нг/мл |
38 |
24,6 ± 2,67 |
28 |
24,4 ± 2,62 |
66 |
24,5 ± 1,88 |
|
Общий белок, г/л |
58 |
70,2 ± 0,89 |
32 |
68,8 ± 1,12 |
90 |
69,7 ± 0,70 |
|
Мочевина, моль/л |
52 |
16,2 ± 0,64 |
30 |
15,7 ± 0,70 |
82 |
16,0 ± 0,48 |
|
Креатинин, мкмоль/л |
58 |
125,4 ± 3,67 |
29 |
125,2 ± 5,44 |
87 |
125,3 ± 3,03 |
|
Глюкоза, моль/л |
58 |
5,02 ± 0,16 |
29 |
4,8 ± 0,24 |
87 |
5,0 ± 0,13 |
|
АЛТ, ед/л |
62 |
61,9 ± 5,77 |
32 |
78,4 ± 12,35 ** |
94 |
67,5 ± 5,69 |
|
АСТ, ед/л |
63 |
312,8 ± 24,17 |
32 |
315,1 ± 51,34 |
95 |
313,6 ± 23,42 |
|
ГГТ, ед/л |
60 |
37,9 ± 2,12 |
30 |
36,5 ± 3,48 |
90 |
37,4 ± 1,82 |
|
ЩФ, ед/л |
52 |
543,9 ± 57,98 |
26 |
809,0 ± 74,09 * |
78 |
632,2 ± 47,76 |
|
Железо, мкмоль/л |
55 |
28,4 ± 1,36 |
33 |
25,1 ± 1,15 |
88 |
27,1 ± 0,96 |
Примечание . Достоверность различий между данным сроком беременности и предыдущим: * – P
< 0,01; ** – P < 0,001.
Из данной таблицы видно, что достоверные различия по гематологическим показателям между группой благополучно родивших самок и неблагополучно родивших отмечаются в первую очередь по концентрации гемоглобина. У благополучно родивших количество гемоглобина в крови оказалось достоверно выше (P <0,001), чем у самок другой группы. В то же время у первых оказался достоверно ниже уровень аланинтрансаминазы (АЛТ) (P <0,001), что указывает на отсутствие токсикоза, целостность гепатоцитов и нормальную функцию печени. При этом относительное содержание палочкоядерных нейтрофилов тоже оказалось достовер- но меньше (P <0,01), чем у неблагополучно родивших афалин, у которых отмечался более выраженный «сдвиг влево» лейкоформулы. Этот факт говорит о большей востребованности у второй группы животных фагоцитоза нейтрофилами тканевых обломков и уничтожения опсонизированных микроорганизмов. В первой группе афалин также отмечался более низкий уровень щелочной фосфотазы (ЩФ) (P <0,01). Однако, если разница по содержанию гемоглобина, АЛТ и палочкоядерных нейтрофилов имеет клиническую ценность, то разница по концентрации ЩФ, уровень которой с возрастом афалин снижается приблизительно от 1300 до
200 ед/л, скорее указывает на относительно молодой возраст неблагополучно родивших самок (9,8 ± 0,87 лет), который оказался достоверно (P < 0,001) ниже возраста нормально родивших (16,6 ± 1,20 лет). Таким образом, данный показатель также имеет важное значение в вопросе благополучного родоразрешения самок дельфинов.
Выводы. В результате проведённых исследований нам удалось определить сроки беременности у самок черноморской афалины во второй половине беременности с помощью их ультразвукового обследования в фетальный период, выявить динамические различия в размерах головы и грудной клетки плода. Использование ультразвука для контроля беременности у китообразных в неволе предоставляет ценные данные о морфологии, развитии и благополучии плода. При этом учет таких параметров, как размеры грудной клетки и головы, могут играть важную роль в установлении сроков беременности. Кроме того, оценка размеров грудной клетки и головы является важной частью обследования плода для контроля его правильного развития.
Помимо этого, выявлены достоверные различия гематологических показателей между группой благополучно родивших самок с одной стороны, и неблагополучно родивших – с другой. Клиническую ценность имели различия по концентрации гемоглобина (P <0,001), аланин-трансаминазы (P <0,001) и относительному содержанию палочкоядерных нейтрофилов (P <0,01). Выявлен относительно молодой возраст (9,8±0,87 лет) неблагополучно родивших самок, который оказался достоверно (P <0,001) ниже возраста (16,6 ± 1,20 лет) благополучно родивших дельфинов.
Таким образом, становится ясно, что использование метода ультразвуковой диагностики актуально для определения беременности у черноморских афалин, её сроков и наблюдения за развитием плода. В то же время гематологические показатели беременных самок дают объективную оценку вероятности физиологически нормального родоразрешения.
Список литературы Конроль развития плода у самок черноморской афалины (Tursiops truncatus ponticus Barabash,1940) во второй половине беременности
- Brook F. The Use of Diagnostic Ultrasound in Assessment of the Reproductive Status of the Bottlenose Dolphin, Tursiops aduncas, in Captivity and Applications in Management of a Controlled Breeding Programme: Ph.D. thesis. - Hong Kong, Hong Kong Polytechnic University, Kowloon, 1997. - 339 p.
- Robeck T.R., Atkinson S.K.C., Brook F. Reproduction. In: Dierauf LA, Gulland FMD. = CRC Handbook of Marine Mammal Medicine. Second Edition. Boca Raton, 2001. - P. 193-226.
- Duffield D.A., Shell D., Dudley M. Demographic analysis of breeding bottlenose dolphins in North American zoological facilities: 1976-1998, in Report from the Bottlenose Dolphin Breeding Workshop, Duffield D.A. and Robeck T.R. (Eds.). - American Zoological Association Marine Mammal Taxon Advisory Group, Silver Spring, MD, 2000. - P. 139-156.
- Sawyer-Steffan J.E., Kirby. V.L. A study of serum steroid hormone levels in captive female bottlenose dolphins, their correlation with reproductive status, and their application to ovulation induction in captivity. - National Technical Information Service, PB80-177199, Springfield, VA, 1980.
- Kirby V.L., Ridgway S.H. Hormonal evidence of spontaneous ovulation in captive dolphins (Tursiops truncatus and Delphinus delphis) // Rep. Int. Whaling Comm. - Spec. Issue 6, 1984. - P. 459-464.
- Schroeder J.P. Reproductive aspects of marine mammal. - In CRC Handbook of Marine Mammal Medicine, Dierauf L.A. (Ed.), CRC Press, Boca Raton, FL, 1990. - P. 353-369.
- Яблоков А.В., Белькович В.М., Борисов В.И. Киты и дельфины. - М.: Наука, 1972. - 472 с.
- Ожаровская Л.В. Морфология репродуктивной системы черноморской афалины // Черноморская афалина Tursiops truncatus ponticus: морфология, физиология, акустика, гидродинамика / под ред. В.Е. Соколова, Е.В. Романенко. - М.: Наука, 1997. - С. 102-113.
- Ridgwey S.H., Benirschke K. Breeding dolphins: Present status, suggestions for the future: Final rep. to US Marine Mammals Comm. - Wash. (D.C.), 1977. - P. 308.
- Gray K.N., Conclin R.H. Multiple births and cardiac anomalies in the bottlenosed dolphin // Wildlife Diseases. - 1974. - Vol. 10. - P. 155-157.
- Томилин А.Г. Китообразные // Звери СССР и прилежащих стран. - М.: АН СССР, 1957. - Т. 9. - С. 756.
- Цивцивадзе Е.Б., Новикова С.В. Многоплодная беременность: современный взгляд на проблему ведения беременности и родов (обзор литературы) // Регулярные выпуски "РМЖ". - 2014. - № 1. - С. 16.
- Baird D.T., Smith K.B. Inhibin and related peptides in the regulation of reproduction, in Oxford Review of Reproductive Biology, Milligan, S.R. (Ed.). - Oxford University Press, Oxford, U.K., 1993. - P. 191-232.
- Hendricks D.M. Biochemistry and physiology of the gonadal hormones, in Reproduction in Domestic Animals, Cupps, P.T. (Ed.). - Academic Press, San Diego, CA, 1991. - P. 81-118.
- Williamson P., Gales N.J., Lister S. Use of real-time B-mode ultrasound for pregnancy diagnosis and measurement of fetal growth rate in captive bottlenose dolphins (Tursiops truncatus) // J. Reprod. Fertil. - 1990. - 88. - P. 543-548.
