Квантово-химическая модель гормонов щитовидной железы как доноров и переносчиков электронов

Автор: Кузнецова Н.Б., Кузнецов П.Е.
Журнал: Вестник Воронежского государственного университета инженерных технологий @vestnik-vsuet
Рубрика: Фундаментальная и прикладная химия, химическая технология
Статья в выпуске: 4 (58), 2013 года.
Бесплатный доступ
На основе рентгеноструктурных данных из Protein Data Base и квантово-химических расчетов взаимодействия 3,3',5'-трийодтиронина (Т 3) с активным центром тиреоидного рецептора, с молекулами кислорода и АТФ предложен механизм действия гормонов щитовидной железы с биомишенями. Он заключается в том, что при взаимодействии Т 3, Т 4 с биомишенями происходит перенос электрона, а Т 3, Т 4 могут выступать в качестве его доноров или акцепторов. Являясь переносчиками электронов, Т 3, Т 4 могут взаимодействовать с молекулами, находящимися в триплетном состоянии (О 2, АТФ, тиреоидный рецептор).
Трийодтиронин, тиреоидный рецептор, механизм действия, квантово-химическая модель
Короткий адрес: https://sciup.org/14040123
IDR: 14040123 | УДК: 544.18/577.29:577.17
Quantum-chemical model of thyroid hormones as electron donor and electron carriers
A new mechanism of interaction of thyroid gland hormones with biotargets is offered on the basis of X-ray structural data from Protein Data Base and quantum-chemical computation of 3,3',5'-triiodothyronine (T 3) interaction with the active center of thyroidal receptor as well as with oxygen molecules and ATP. The nature of the mechanism is that when T 3 and T 4 interact with biotargets, a transportation of electron occurs, and T 3 and T 4 can act as donors and acceptors of electron. Being electron carriers, T 3 and T 4 can interact with molecules that are in triplet state (O 2, ATP, thyroidal receptor).
Текст научной статьи Квантово-химическая модель гормонов щитовидной железы как доноров и переносчиков электронов
Экспериментальные данные показывают, что тиреоидные гормоны влияют на сердечнососудистую, нервную, дыхательную системы [7]. Анализ этих данных с позиций молекулярных механизмов указывает, что все они могут быть объяснены тремя взаимодействиями тиреоидных гормонов: с ядерными рецепторами, с биомишенями в окислительных реакциях и с мембранными белками. Опишем кратко все три вида взаимодествия.
Рецепторы Т 3 , Т 4 (тироксина) относятся к ядерным рецепторам. Взаимодействуя с лигандами, они регулируют экспрессию генов, управляя синтезом белков. Эффект Т 3 , Т 4 зависит от продолжительности их связывания с рецептором и числа связанных рецепторов [1]. Показано, что рецепторы Т 3 и ретиноевой кислоты взаимодействуют и образуют более активный гетеродимер [5]. Связывание Т 3 с рецептором блокирует его ингибирующее влияние [2].
Тиреоидные гормоны оказывают стимулирующее воздействие на синтез жирных кислот, при этом значительно увеличивается скорость поглощения кислорода. Увеличивается катаболическое окисление липидов и холесте-рола, что также требует больших затрат кислорода. Отмечается калоригенное действие
тиреоидных гормонов [7], что может быть связано с усилением процессов окисления и фосфорилирования в реакции окислительного фосфорилирования. Т 3 , Т 4 повышают потребление митохондриями кислорода, повышают в них теплообразование, обнаруживается «утечка» протонов через внутреннюю мембрану митохондрий, увеличивается синтез АТФ, митохондриальное дыхание может усиливаться в 2 раза, увеличивается площадь поверхности внутренних мембран митохондрий [1, 2]. Объясняются эти эксперименты влиянием Т 3 , Т 4 на липидный состав мембран, усилением эффективности трансмембранного насоса, усилением проницаемости мембран для ионов Са++, что активирует фермент малатдегидрогеназу и процессы дыхания. Однако отмечается, что неизвестны места связывания Т 3 , Т 4 в митохондриях.
Т 3 , Т 4 влияют на активность Na+/K+-насоса за счет усиления проницаемости мембран и увеличивают активность Na+/K+ - AТ-Фазы, которая гидролизует АТФ в присутствии Na+ и K+ (также окислительный процесс). Также увеличивается перенос в клетку кальция, что является результатом стимуляции гормонами Са++АТФазы [1, 2].
Таким образом, на молекулярном уровне все многообразие экспериментальных данных можно объяснить тремя взаимодействиями Т 3 ,
Т 4 : с ядерным рецептором, с молекулой кислорода в окислительных реакциях и с молекулой АТФ мембранных белков. Исследованию этих взаимодействий и посвящена настоящая статья.
Наши исследования, основанные лишь на изучении электронной структуры, показали, что Т 3 , Т 4 обладают необычными свойствами – свойствами доноров и переносчиков электронов. Ниже приводятся расчетные данные по трем вышеперечисленным взаимодействиям гормонов Т 3 , Т 4 как доноров и акцепторов электронной плотности.
B Protein Data Base [5] содержатся определенные рентгеноструктурным анализом координаты атомов лиганд связанных гетеродимеров рецепторов Т 3 гормона и ретиноевой кислоты. Исследование структуры комплекса гормона Т3 показало, что его взаимодействие происходит с гистидином тиреоидного рецептора, т.к. его гидроксильная группа образует водородную связь с атомом водорода гистидина длиной 1,82 Å (рисунок 1). Соответственно, ретиноевая кислота в комплексе с ретиноевым рецептором образует водородную связь с аргинином-76.
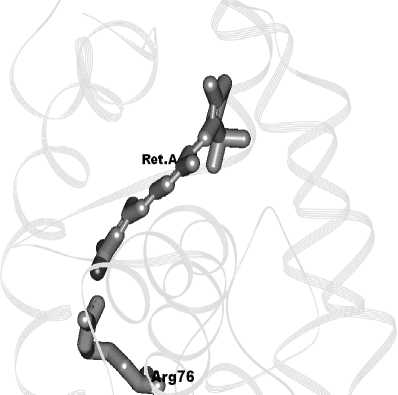
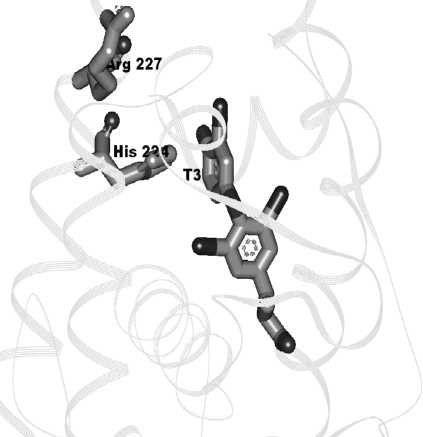
Рисунок 1 - Рентгеноструктурные данные комплекса (гетеродимера) Т 3 , тиреоидного рецептора и ретиноевой кислоты, ретиноевого рецептора
Активный центр тиреоидного рецептора с зарядом +2 содержит две активные аминокислоты – гистидин-224 и аргинин-227. Можно предположить, что активный центр является акцептором электронной плотности.
Исследования Protein Data Base показали, что для активации ядерного тиреоидного рецептора и последующей транскрипции с гена необходимо связывание ретиноевой кислоты с гетерорецептором [5].
Расчеты проводились программами Hy-perChem 7e [6], Discovery Studio Client v2.5.0.9164 [4]. Наши квантово-химические исследования комплекса активного центра тиреоидного рецептора и гормона Т 3 указывают, что при этом взаимодействия между ними и переноса электронной плотности не происходит. При этом учитывалось, что основное состояние активного центра – триплетное. Проведенные различными методами (РM 3 , Minimal (STO-3G)) расчеты, показали, что синглетное состояние активного центра рецептора в виде цвиттер-иона не существует, т.к. отсутствует сходимость в процедуре самосогласования.
Далее исследовалось влияние ретиноевой кислоты. Рассматривалась упрощенная модель, которая описывает взаимодействие Т 3 с гетеродимером и ретиноевой кислотой в виде комплекса ретиноевой кислоты с активным центром тиреоидного рецептора и гормоном Т 3 (рисунок 2). Модель комплекса сформирована в соответствии с рентгеноструктурными данными: активный центр рецептора находится в виде цвиттер-иона, Т 3 содержит концевые группы СОО- и NH 2 , рe-тиноевая кислота образует водородную связь с аргинином и содержит группу СООН. Расстояние между атомом кислорода гидроксильной группы и атомом водорода гистидина рецептора равно длине водородной связи.
Распределение спиновой и зарядовой плотности в комплексе после взаимодействия показывает, что в каждой из областей, отмеченных на рисунке 2 пунктиром, находится на валентной оболочке по одному электрону с параллельными спинами. До взаимодействия электроны с параллельными спинами находились на рецепторе. Следовательно, произошло перераспределение электронов. Так, заряд на Т 3 до взаимодействия был равен (-1), а после – равен нулю.
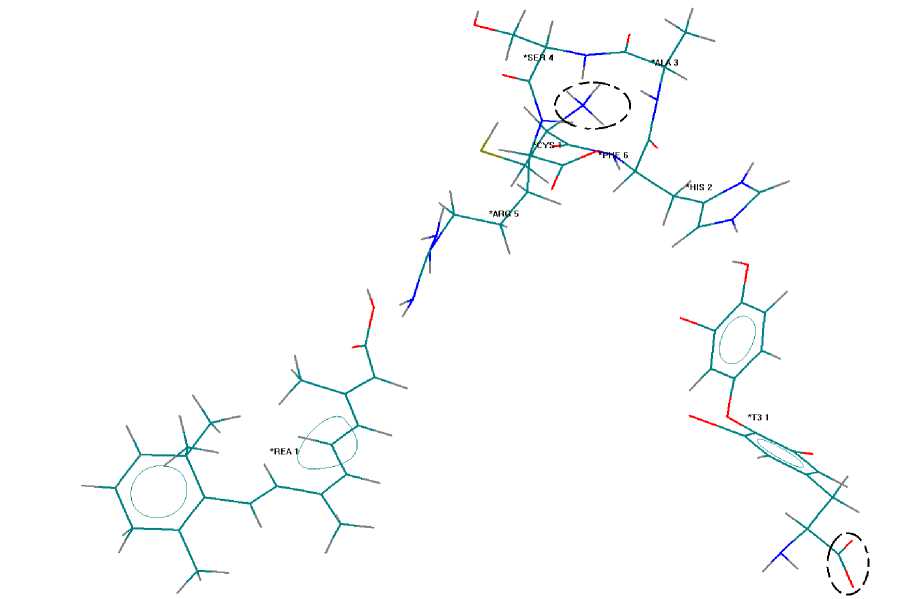
Рисунок 2 - Комплекс гормона Т3 с активным центром тиреоидного рецептора и ретиноевой кислотой
Следовательно, электрон перераспределился с гормона на рецептор. Соответственно, на активном центре рецептора до взаимодействия заряд был равен (+2), а после - (+1). Таким образом, присоединение к комплексу ретиноевой кислоты потенциирует перенос электрона с гормона Т3 на тиреоидный рецептор.
Таким образом, гормон Т3, как донор электрона, переносит электрон на рецептор и этим активирует его. Расчеты показали, что перенос электрона термодинамически выгоден.
Предложенный механизм действия тиреоидных гормонов как доноров электронов объясняет экспериментальные данные по ядерным тиреоидным рецепторам. Так, одновременное связывание Т3 и ретиноевой кислоты с ядерным рецептором активирует его путем переноса электрона, что ведет к последующему связыванию с соответствующим промотором и к активации экспрессии генов.
Взаимодействие с молекулой кислорода описано далее. Основное состояние молекулы кислорода - триплетное.
Известно, что Т3, Т4 стимулируют образование активных форм кислорода [1]. Проанализируем механизм взаимодействия гидроксила Т3 с кислородом на примере реакции окислительного фосфорилирования. Цитохромоксидаза в комплексе с кислородом является конечным звеном в цепи переноса электронов в реакции окис- лительного фосфорилирования и катализирует перенос четырех электронов на кислород с образованием двух молекул воды. Четыре иона водорода Н+ поступают из водной среды.
Таким образом, вполне вероятно, что гормоны щитовидной железы могут образовывать комплекс с кислородом цитохромоксидазы, а после присоединения протона к кислороду через водородную связь инициировать перенос электрона от молекулы гормона на молекулу кислорода, усиливая митохондриальное дыхание.
При этом молекула гормона рассматривается в качестве донора электронов, а молекула кислорода - в качестве акцептора.
На первом этапе исследований был изучен комплекс трийодтиронина с триплетным молекулярным кислородом. Установлено, что комплекс (трийодтиронин---3О=О) образуется с помощью водородной связи, но перераспределения спина в нем не происходит.
Рассмотрим, как преобразуется комплекс при присоединении к нему протона (трийодти-ронин---3О=О---Н+). На рисунке 3 представлена структура комплекса. В каждой из областей, отмеченных на рисунке пунктиром, находится на валентной оболочке по одному электрону с параллельными спинами. До взаимодействия электроны с параллельными спинами находились на молекуле кислорода.
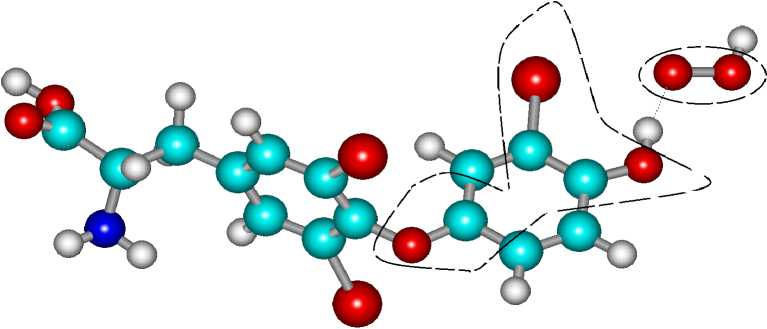
Рисунок 3 - Структура комплекса (трийодтиронин---3О=О---Н+)
Расчеты показали, что молекула трийод-тиронина трансформируется в катион-радикал (заряд - (+1), спин - (+1/2)), происходит перераспределение электронной плотности и перенос электрона на молекулу кислорода. Расчеты подтвердили термодинамическую разрешен-ность процесса.
Таким образом, продуктами переноса электрона с молекулы трийодтиронина на молекулярный кислород цитохромоксидазы являются катион-радикал трийодтиронина и пероксирадикал. В определенных условиях катион-радикал может восстанавливаться до трийодтиронина, вновь образовывать комплекс с кислородом цитохромоксидазы и инициировать перенос электрона на молекулу кислорода. Следовательно, молекула трийодтиронина может выступать катализатором описанной реакции, являясь, в сущности, переносчиком электронов. Пероксирадикал в результате реакций дисмутации может превращаться в другие активные формы кислорода [1].
Аналогичный механизм взаимодействия с молекулой кислорода может быть характерен и для гормона тироксина, что подтверждается аналогичными квантово-химическими расчетами. Расчеты показали, что Т3 быстрее переносит электрон на кислород, чем Т4, этим и объясняется меньшая активность Т4.
Предлагаемый биохимический механизм тироксина и трийодтиронина соответствует известным экспериментальным данным, в частности представлению о том, что гормоны щитовидной железы являются стимуляторами энергетического метаболизма, способствуя повышению скорости переноса электронов, а, следовательно, ускорению синтеза АТФ в реакции окислительного фосфорилирования. Гормоны щитовидной железы, взаимодействуя с кислородом цитохромоксидазы, инициируют перенос электронов и образование дополнительной цепи переноса электронов.
Известно, что гормоны вызывают индукцию набухания митохондрий [1], которая может быть инициирована формированием дополнительной цепи переноса электронов, стимулируемой гормонами щитовидной железы, и вследствие этого, увеличением числа компонентов мембраны.
Кроме того, предлагаемая схема объясняет увеличение образования активных форм кислорода, которые могут элиминироваться вследствие генерации пероксирадикала.
Таким образом, обсуждаемый возможный механизм биохимического действия гормонов щитовидной железы не противоречит известным экспериментальным данным. Более того, становятся объяснимыми такие факты, как стимулирование энергетического метаболизма, увеличение скорости переноса электронов в реакции окислительного фосфорилирования, образование активных форм кислорода, индукция набухания митохондрий, «утечка» протонов через внутреннюю мембрану митохондрий и другие данные.
Главный результат действия тиреоидных гормонов состоит в увеличении скорости основного обмена. До сих пор остается неизвестным, как тиреоидные гормоны регулируют скорость основного обмена. Изучение этой проблемы с позиций теоретической химии, электронного строения вещества и анализа биохимических свойств тиреоидных гормонов методами квантовой химии позволяет рассматривать данные соединения в качестве эффективных химических переносчиков электронов, увеличивающих скорость окислительных реакций.
Таким образом, объясняется уникальность эффекта тиреоидных гормонов, которые осуществляют мощную регуляторную функ- цию, присутствуя в биообъектах в малых концентрациях. При этом выраженность эффекта объясняется тем, что молекулы тиреоидных гормонов являются катализаторами переноса электронов, когда одна молекула инициирует перенос потока электронов.
Опишем взаимодействие с мембранными белками. Известно, что Т 3 стимулируют Na+/K+ - АТФазу и Са++АТФазу [1]. С позиций молекулярных механизмов это можно объяснить взаимодействием Т 3 с АТФ. Расчеты показали, что триплетное состояние АТФ с зарядом (-4) устойчивее синглетного состояния. Таким образом, при взаимодействии АТФ с Т 3 возможен перенос электрона. На рисунке 4 представлен комплекс [Т 3 -H+ – 3АТФ], образованный с помощью водородной связи между
NH3+-группой протонированного Т3 и NH2-группой АТФ. Из анализа распределения спиновой плотности в этом компл ексе следует, что в комплексе происходит перенос электрона с молекулы АТФ на гормон Т3. Термодина- мически такой процесс является разрешенным.
В результате молекула АТФ трансформируется в реакционноспособный анион-радикал (что ускоряет гидролиз), a Т 3 является акцептором электрона. В результате Т 3 выступает в качестве катализатора в реакции гидролиза АТФ АТФазой. Более того, известно, что гидролиз АТФ Са-АТФазой невозможен без присоединения двух ионов Са2+ [3], которые, видимо, являются акцепторами электронов как и Т 3 в данном случае. Т.е. Т 3 выполняет роль Са2+ и ускоряет реакцию гидролиза АТФ.
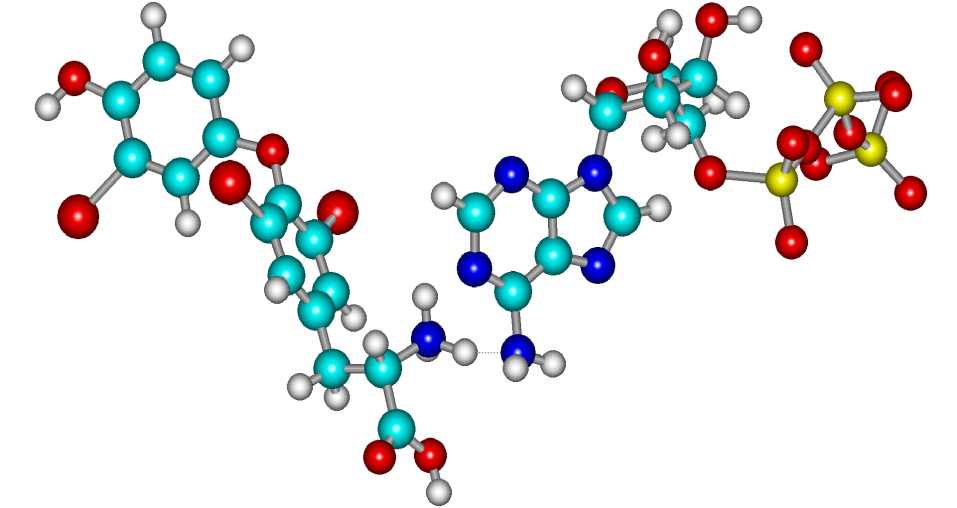
Рисунок 4 - Структура комплекса [Т3-H+ – 3АТФ]
Таким образом, гормоны щитовидной железы могут выступать как в качестве донора, так и в качестве акцептора электронов в процессах переноса электронов, являясь по сути переносчиками электронов. С ними могут взаимодействовать молекулы, находящиеся в триплетном состоянии: ядерные рецепторы Т 3 , Т 4 ; молекулы кислорода и АТФ и т.д. Этим и объясняется все многообразие воздействия этих гормонов на организмы, причем одна молекула Т 3 или Т 4 может множество раз катализировать различные реакции переноса электрона, при этом ее структура остается неизменной. Этим и объясняется гормональные эффекты Т 3 , Т 4 , когда их малое количество оказывает выраженное воздействие на клетки.
