Квазидискретные особенности накопления биомассы зерен в колосе пшеницы

Автор: Бабицкий А.Ф., Русанов A.M., Канаровский Е.Ю.
Журнал: Известия Волгоградского государственного педагогического университета @izvestia-vspu
Рубрика: Физика и математика
Статья в выпуске: 4 (10), 2004 года.
Бесплатный доступ
Короткий адрес: https://sciup.org/148162674
IDR: 148162674
Текст статьи Квазидискретные особенности накопления биомассы зерен в колосе пшеницы
Дискретный характер явлений и процессов (квантование) проявляется в микросистемах на атомном и молекулярном уровнях. В макросистемах квантование не проявляется, имеет место континуальное распределение свойств элементов системы по кривой Гаусса, которая описывает аддитивное поведение множества независимых элементов, на которые действует множество независимых переменных. Однако характерная особенность биологических систем и состоит в том, что они составлены из множества элементов, находящихся в тесной взаимно коррелятивной связи и поэтому, как правило, не подчиняющихся кривой нормального распределения. На статистических кривых распределения процессов различной природы можно обнаруживать дискретные состояния в ходе флуктуаций в макроскопических процессах [1; 2]. Довольно часто встречаются многовершинные кривые распределения, которые свидетельствуют о квазидискретности биологических макросистем [3—8]. Изучение этой многовершинности и тонкой структуры статистических кривых распределения позволяет обнаружить целый ряд закономерностей взаимодействия и взаимовлияния элементов биологической системы между собой. Взаимосвязь между континуальной и дискретной изменчивостью описана теоретически [8]. Хорошо доказано, что рост и развитие биологических объектов происходят ритмически [9], что отражает ритмичность целого ряда физических и химических процессов [10], следовательно, и статистические кривые их распределения также должны в своей форме отражать эту ритмичность. О возможности на макроуровне системы выявления процессов, происходящих на более глубоких ее уровнях, свидетельствуют данные, представленные в нижеследующем сообщении.
Объект и методы исследования
В качестве объекта исследования использовали зерна пшеницы «Кооператорка». С площади 1 кв. м был срезан сноп из стеблей на стадии полной зрелости зерна. Из каждого колоса выделялись зерна и взвешивались на аналитических весах ВЛР=20 с точностью 0,1 мг. Всего было взвешено 2530 зерен. Полученные данные математически обработаны с помощью программ Excel и «Гусеница» (Caterpillar).
Экспериментальные данные и их обработка
Экспериментальная кривая распределения зерновок пшеницы по массе представлена на рис. 1 (кривая 1), которая обладает асимметричным смещением вправо и отрицательным эксцессом, т.е. значительно отличается от кривой нормального распределения и имеет тонкую структуру, отражающую взаимовлияние зерен в колосе. Для выявле- ния этого взаимовлияния из экспериментальной кривой распределения была вычтена теоретическая кривая нормального распределения (рис.1, кривая 2), соответствующая средней величине 46,219 мг и среднеквадратическому отклонению 11,412 мг. В результате получена разностная кривая (рис.1, кривая 3), из которой в программе Excel выделен тренд (рис.1, кривая 4). Анализ кривой распределения позволяет выделить три области масс зерновок: 1) малые - от 4 до 30 мг; 2) средние - от 30 до 49 мг; 3) крупные - от 49 до 65 мг. В области малых зерновок экспериментальная кривая распределения соответствует нормальной кривой, что свидетельствует о том, что в этом диапазоне масс зерновок нет заметной асимметрии во взаимовлиянии зерновок в колосе и наблюдается только тонкая структура распределения.
В области средних масс разностная кривая 3 имеет впадину, а в области больших масс наблюдается подъем. Это свидетельствует о том, что самое сильное взаимовлияние в колосе имеется между этими двумя группами зерновок: крупные зерновки поглощают питательные вещества не только со стержня колоса, но и из зерновок среднего размера. Следовательно, в процессе формирования крупных зерновок заметную роль играет перекачка питательных веществ из средних.
Вычитанием тренда (кривая 4) из разностной кривой 3 получена кривая 5, представляющая тонкую структуру взаимовлияния зерновок в колосе. Разложение кривой 5 на частотные составляющие выполнено по программе «Гусеница».
Рис.1. Распределение зерен пшеницы «Кооператорка» по массе
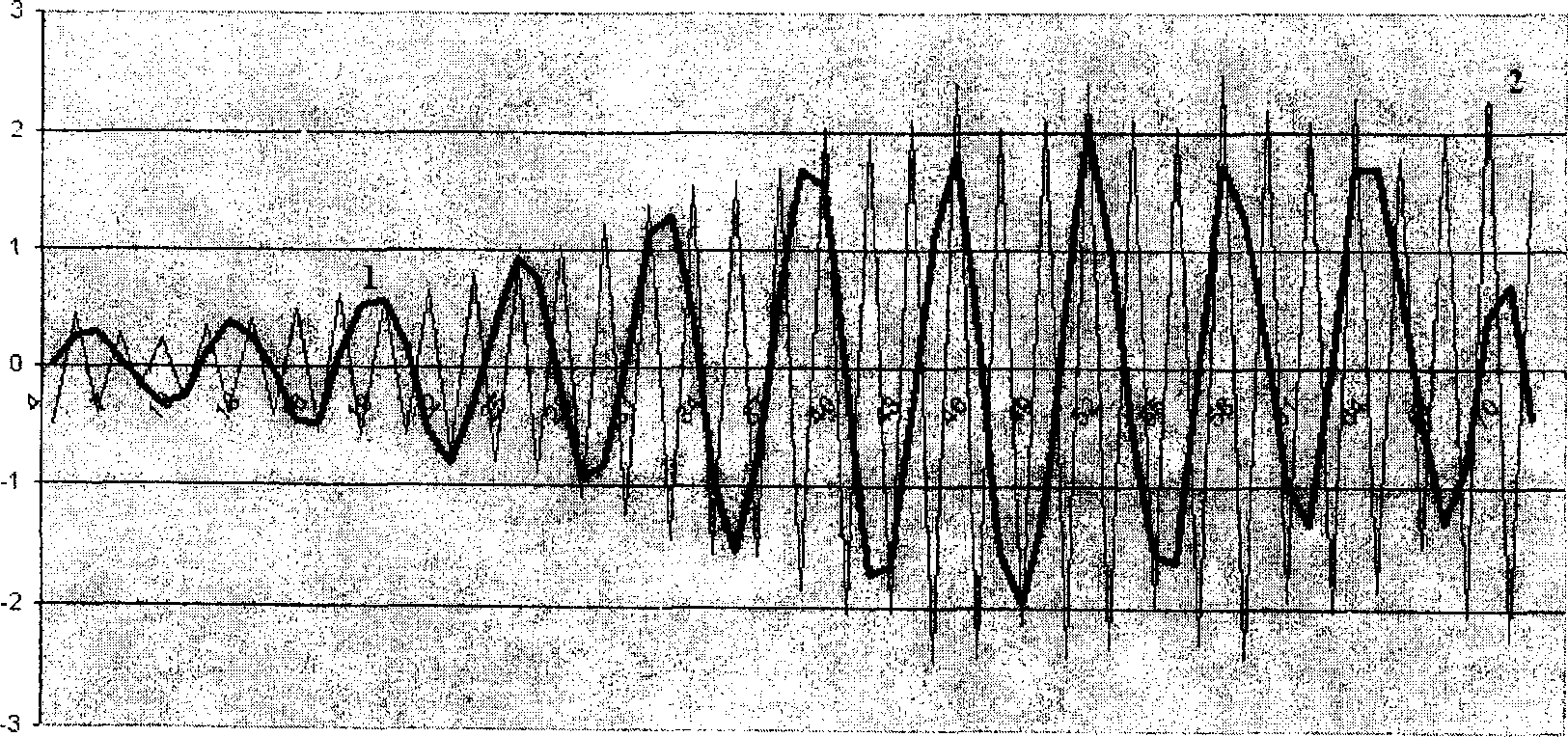
Рис.2. Основные периодические составляющие распределения зерен пшеницы «Кооператорка» по массе
В результате выделен целый ряд периодических составляющих, основные из которых представлены на рис. 2. Периодическая составляющая 1 имеет период 8 мг, по амплитуде возрастает от области мелких зерновок и достигает максимума в области перехода от средних зерновок к крупным, затем уменьшается. Вторая периодическая составляющая с периодом 1 мг также минимальна в области мелких зерновок и плавно возрастает в области средних зерновок, затем сохраняет свою амплитуду неизменной вплоть до района крупных зерновок. Остальные гармонические составляющие имеют меньший вклад.
Комментарий
Представленные нами обширные экспериментальные данные (более 2500 измерений), обработанные с использованием вычислительных программ, свидетельствуют о том, что биологические системы не подчиняются кривым нормального распределения. Этого и следовало ожидать, поскольку между элементами этой системы всегда существуют различного рода направленные взаимовлияния, поэтому вопрос об использовании в биологии статистических методов, в основе которых лежит кривая нормального распределения, нужно решать в каждом конкретном случае, учитывая степень детализации изучаемого явления. Так, К.Пирсон статистические кривые распределения биологических объектов и процессов предложил описывать несколькими типами специфических кривых [11].
Тонкая структура гистограммы статистических распределений позволяет из макроуровня выделить дискретные ритмические взаимовлияния, происходящие на микроуровне. Наибольшая амплитуда этих ритмических взаимовлияний наблюдается в области ингибирования роста средних зерновок более крупными в области впадины (кривые 3, 4), соответствующей максимуму кривой нормального распределения (кривая 2). В этой области кривой располагаются зерновки из средней по длине части колоса, где в колосках формируются самые крупные и самые мелкие добавочные зерна, которые вследствие ингибирования более ранними по времени образования крупными зернами не могут набрать достаточную массу и созревают в виде мелких зерен. На самих мелких зернах из кривой распределения ритмичность экспериментально просматривается менее заметно.
Отрицательное влияние многоцветковости колосков в средней части колоса на продуктивность пшеницы объясняет безуспешные попытки выведения практиками-селекционерами высокопродуктивных ветвистых сортов пшеницы. Наиболее продуктивные формы пшеницы и кукурузы имеют правильно и регулярно расположенные в колосе и початке зерновки, что позволяет свести к минимуму ингибиторные взаимодействия между зернами.
Анализируя периодические кривые, предварительно можно принять, что периодическая компонента 1 мг может быть интерпретирована как суточная компонента периодического накопления биомассы зерновкой. Эта суточная компонента может отражать периодичность накопления биомассы за счет процесса деления клеток и притока органического вещества из листьев в периодическом дневном процессе фотосинтеза. Компоненту с периодом 8 мг можно приписать недельной компоненте, являющейся результатом межклеточного взаимодействия в процессе дифференцирования вновь образовавшихся клеток при формировании в них цитоскелета [12]. Окончательные выводы возможны только после распространения этих исследований на обширный ряд сортов пшеницы различной продуктивности. Из наших данных следует, что дискретные особенности, проявляющиеся в экспериментальной кривой распределения, можно связать с особенностями накопления биомассы в процессе роста при взаимовлиянии зерен в колосе.
