Late Pleistocene environments of East Kazakhstan (based on Ushbulak site materials)
")
Author: Anoikin A.A., Pavlenok G.D., Silaev V.I., Shnaider S.V., Koval V.V., Devyatova A.Y., Vasiliev S.K., Bocharova E.N., Kharevich V.M., Taimagambetov Z.K.
Journal: Archaeology, Ethnology & Anthropology of Eurasia @journal-aeae-en
Article in issue: 3 т.51, 2023.
Free access
We reconstruct environmental conditions at various stages of the Late Pleistocene and Early Holocene of East Kazakhstan. The reconstructions are based on materials from the stratified Ushbulak site in the Shilikta Valley, spanning a period from the Early Upper Paleolithic to the Bronze Age. Climatic changes were evaluated using natural science methods-mineralogical, ZooArchaeology by Mass Spectrometry (ZooMS), OSL- and AMS-dating, etc. Sevaral stages, relating to environmental changes, are evaluated. The fi rst period (~52–37 ka BP) was period of moderately warm and relatively humid climate, with predominantly forest-steppe, meadow-steppe, and semidesert landscapes. The second period (~25–21 ka BP) coincided with a transition from a moderately warm to a very cold and more arid climate dominated by steppes. The third period (~18–16 ka BP) was transitional from the glacial maximum to the postglacial interstadial, with a relatively cool and arid climate and mostly steppe and forest-steppe landscapes. The fourth period (~15–14 ka BP) was characterized by the warmest climate in the Late Pleistocene; steppe and forest-steppe vegetation dominated. During the latest, Early Holocene period, the climate was warm and humid, with savanna-like landscapes. The analysis of natural-climatic conditions allows us to conclude that the early stage of the site’s functioning, characterized by the highest intensity of settlement, was optimal for ancient man.
East Kazakhstan, paleoclimate, paleontology, ZooMS, litho-chemical analysis, mineralogical and geochemical analyses
Short address: https://sciup.org/145146925
IDR: 145146925 | DOI: 10.17746/1563-0110.2023.51.3.075-085
Text of the scientific article Late Pleistocene environments of East Kazakhstan (based on Ushbulak site materials)
Until recently, Late Pleistocene environments of East Kazakhstan were reconstructed on the basis of paleofaunal collections from several localities and of surface materials from the Bukhtarma and the Middle Irtysh valleys in the northern part of the region (Kozhamkulova, 1981; Kozhamkulova, Pak, 1988). The discovery of the stratified site of Ushbulak in the southern part of East Kazakhstan in 2016 enhanced paleogeographic studies of this region (Anoikin et al., 2019). In recent years, the study of the Ushbulak archaeological materials has been carried out through a complex of methods—mineralogical, geochemical, isotope, and zooarchaeology by mass spectrometry (ZooMS), as well as OSL- and AMS-dating. Correlation of the obtained results with the data on the main episodes of site occupation during the Upper Paleolithic provides base for reconstructions of the environmental and climatic conditions in the Shilikta Valley during MIS 3–1. The environmental changes, effects on the composition of faunal communities, and the specific economic activities of the ancient population have been considered.
General information
The site of Ushbulak is located at the foothills (1500 m above sea level) of the northeastern end of the Shilikta Valley, in the upper reaches of the Vostochny Stream (Fig. 1).
During the field work, two adjacent archaeological pits and 15 test pits covering a total area of ~50 m2 have been excavated at the site. The stratigraphic profile of the site was ~7 m thick and revealed eight main sedimentary layers, some of which were additionally subdivided into horizons (Fig. 2). The
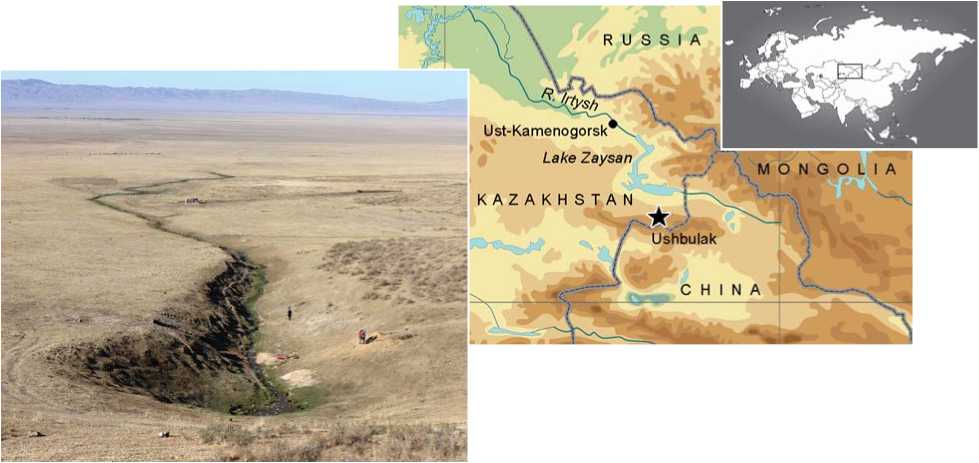
Fig. 1. Location of the site of Ushbulak.
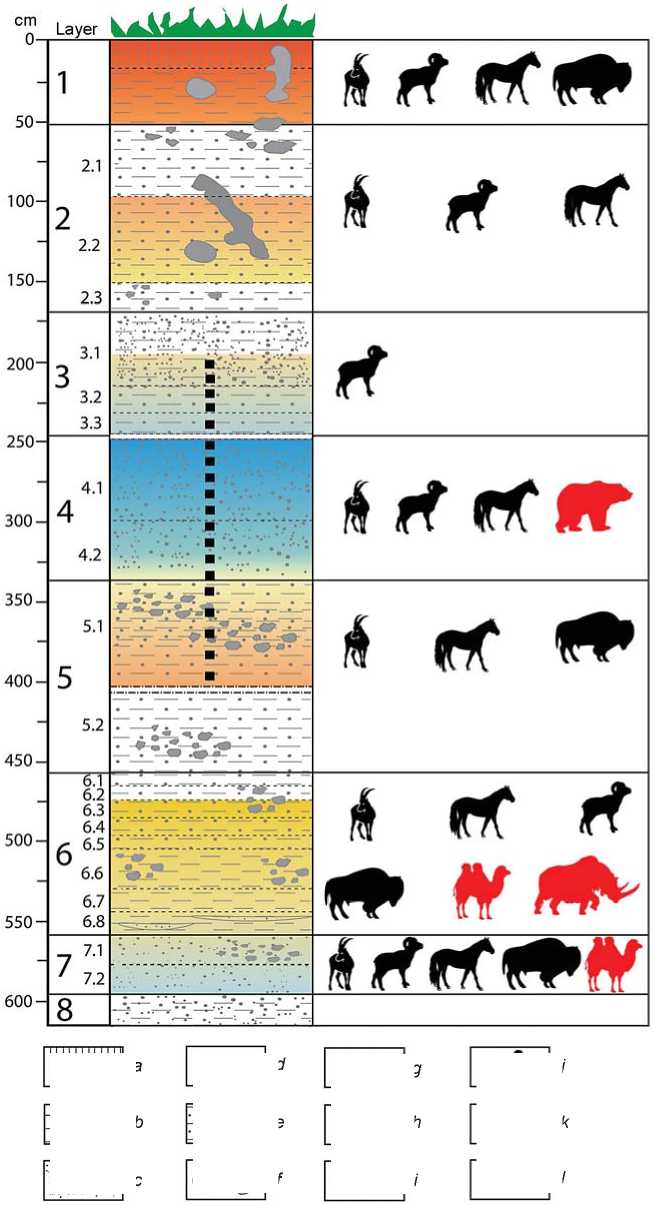
Fig. 2. Scheme of climatic changes in the Ushbulak area, reconstructed by the set of the sciencebased data.
a – heavy loams; b – loams; c – sand; d – breccia; e – sandy loam; f – sink holes; g – sedimentation hiatus; h – borderlines between layers, horizons; i – relative fluctuations in annual average temperatures; j – animal species previously identified at the site; k – animal species found at the site for the first time; l – points of collecting samples for X-ray fluorescence analysis.

lowermost layer 8 is composed of granules and grus of proluvial-mudflow origin, with inclusions of cobbles and heavy loamy matrix. Layer 7 (7.2, 7.1) is a granule and grus stratum with sandy-loam matrix. Layer 6 (6.8–6.1) consists of two sedimentary units. The lower unit is heavy loams with lenses of coarsegrained sands; the upper one is heavy sandy loams with inclusions of light humic loams. The formation of layers 7 and 6 was is connected with the activity of a small stream, which reworked proluvial-slope material from the nearby areas. The upper part of the profile (layers 5–2 (5.2, 5.1; 4.2, 4.1; 3.3–3.1; 2.3–2.1)) consists of proluvial and subaerial sediments, with a significant contribution of eolian material— sands and sandy loams with grus-sand interlayers. Layer 1 is a modern soil profile (Ibid.).
Archaeological material was recorded in layers 7–1. The collection of artifacts from layers 7.2– 6.1 includes more than 16,000 items representing a single lithic industry (Kharevich et al., 2022). The upper layers yield significantly less archaeological materials (~1300 spec.) than the lower layers. According to the technical and typological characteristics of the artifacts and their stratigraphic positions, four main cultural and chronological complexes were identified: the Initial Upper Paleolithic (layers 7.2–6.1), the Middle Upper Paleolithic (layers 5.1–4.1), the Late Upper Paleolithic (layers 3–2), and the Late Bronze Age (layer 1) (Anoikin et al., 2019).
The chronological attribution of the site was made on the basis of a series of 20 OSL- and 5 AMS-dates. According to the Bayesian age model, layers 7.2– 6.1 were most likely accumulated ~52–37 ka BP, layers 5.1–4.1 in the range of ~25–21 ka BP, and layers 3 and 2 ~18–14 ka BP (Ulyanov et al., in press).
Research methods
Paleoclimatic fluctuations in the area of the site at various stages were studied using multiple natural science methods. In addition to the taphonomic and paleoecological assessment of the faunal species’ composition using the classic paleontology techniques, we carried out a ZooMS analysis of a series of unidentifiable mammalian bone fragments. Sample preparation for the ZooMS analysis followed the protocol proposed by M. Buckley (Buckley et al., 2009) and updated by C. Brown (Brown et al., 2020; Shnaider et al., 2022). The findings were interpreted using the database of reference taxa (Welker et al., 2016). Lithochemical studies of the deposits uncovered in the stratigraphic sequence of the site were based on the results of X-ray fluorescence analysis (ARL-9900-XP spectrometer) converted to litho-chemical indices CIA, CIW, and ICV (Nesbitt, Young, 1982; Cox, Lowe, Cullers, 1995).
The study of faunal remains also involved a wide range of modern mineralogical methods (Silaev et al., 2017, 2022): thermal and chemical analyses; determination of Corg content by coulometric titration; X-ray fluorescence analysis (XRD-1800 Shimadzu); optical microscopy (OLYMPUS BX51 complex); X-ray diffractometry (XRD-6000); analytical scanning electron microscopy (JSM-6400 Jeol; Tescan Vega); determination of nanoporosity by nitrogen adsorption/ desorption kinetics (Nova 1200e, Quantachrome Instruments); analysis of the elemental composition of collagen (EA 1110 (CHNS–O)) and the composition of amino acids in collagen (GC-17A Shimadzu with a flame ionization detector); inductively coupled plasma mass spectrometry (NexION 300S Perkin Elmer); infrared spectroscopy (FT-2 Infralum); mass spectrometric analysis of the isotopic composition of C and O in bioapatite and C and N in bone collagen (Delta V. Avantage with the analytical complex of Thermo Fisher Scientific).
Litho-chemical analysis of sediments
For litho-chemical analysis, in trench 1, 19 samples were collected from the sediment column with a step of ~0.1 m (Fig. 2). Changes in the values of CIA- and ICV-indices and the depth-depending proportion of CaO are shown in Fig. 3, 1 .
A gradual increase in CaO concentrations takes place in the middle part of the selected interval (horizon 4.1), reaches its maximum values at a depth of ~2.5 m (boundary between layers 4.1 and 3.3), and then gradually decreases. This indicates the accumulation of readily soluble and mobile forms of calcium (mainly carbonates), which is typical of dry and cold climatic conditions. In the lowermost part of trench 1 (layers 5.1–4.2), CaO concentrations vary insignificantly, which suggests wetter and warmer environments during sedimentation.
The CIA and ICV indices are well correlated with the distribution of CaO along the profile. For example, layers 5.1–4.2 show relatively warm environments, with a slight shift in conditions and intense accumulation of clay minerals; these change sharply in the upper part of horizon 4.1, indicating the coldest conditions. Thus, the upper part of the selected
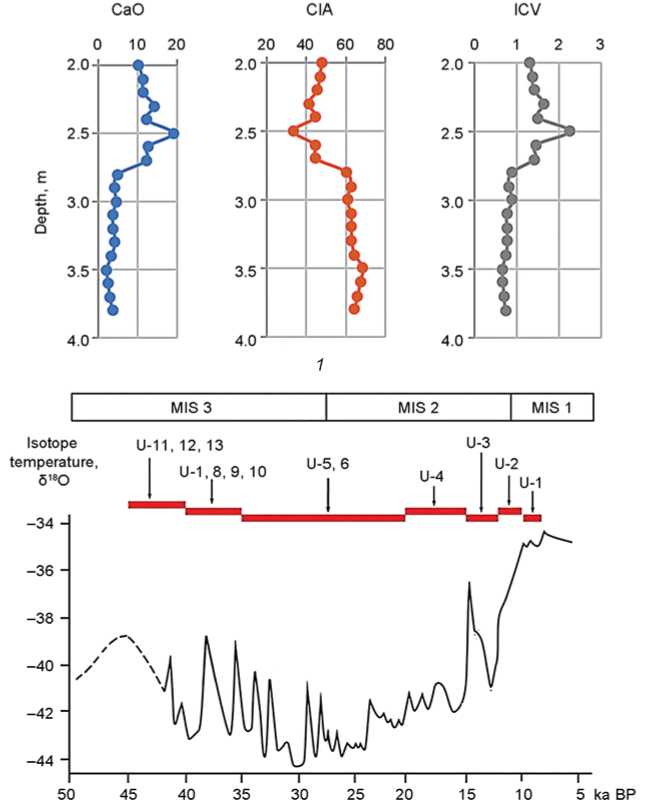
Fig. 3. Geochemical indicators of sedimentation conditions in Ushbulak layers 5.1–3.1 ( 1 ), correlation of the age of fossil bone detritus from Ushbulak (U) with the isotope temperature of living environment of the animals on the modern diagram of Late Pleistocene climatic periodization (after (Silaev et al., 2022)) ( 2 ).
portion of the geological profile (depths 2.7–2.0 m, layers 4.1–3.1) is markedly different in its geochemical composition, which suggests environmental changes during this period of sedimentation. The study of litho-geochemical indicators revealed an intense accumulation of carbonates and a simultaneous decrease in the proportion of clay minerals during this period, corresponding to drier and cooler climatic conditions. In layers 5.1–4.2, the geochemical composition is more monotonous, with an increased content of clay minerals and lower content of CaO, which is typical of a warmer climate.
Analysis of faunal remains
More than 500 bone fragments were recovered from the Pleistocene layers in excavation trenches 1 and 2. The majority of bone fragments do not exceed 50 mm in size. Identifiable remains (21 spec.) are mainly teeth or fragments thereof. The lower layer yielded bones of Bison priscus (primordial bison), Equus ferus (wild horse), Equus hemionus (kulan), Ovis ammon (argali), and Capra sibirica (mountain goat) (see Table ). The remains of argali and horses are the most numerous. Horizon 5.1 contains solitary
Composition of identifiable fauna at Ushbulak, spec.
|
Species, family |
Layer |
Subtotal |
||||||
|
1 |
2 |
3 |
4 |
5 |
6 |
7 |
||
|
Identified to the species level (visually) |
||||||||
|
Equus ferus |
– |
1 |
– |
– |
1 |
4 |
1 |
7 |
|
Equus hemionus |
– |
– |
– |
– |
– |
– |
1 |
1 |
|
Ovis ammon |
– |
– |
1 |
– |
– |
5 |
3 |
9 |
|
Capra sibirica |
– |
– |
– |
– |
1 |
1 |
– |
2 |
|
Bison priscus |
– |
– |
– |
– |
– |
1 |
1 |
2 |
|
Identified to the subfamily / genus level (ZooMS) |
||||||||
|
Ursus sp. |
– |
– |
– |
1 |
– |
– |
– |
1 |
|
Equus sp. |
– |
1 |
– |
5 |
– |
20 |
1 |
27 |
|
Coelodonta sp . |
– |
– |
– |
– |
– |
1 |
– |
1 |
|
Camelidae |
– |
– |
– |
– |
– |
2 |
1 |
3 |
|
Ovis sp. |
1 |
1 |
2 |
2 |
– |
1 |
– |
7 |
|
Ovis/Capra |
2 |
2 |
– |
3 |
– |
10 |
2 |
19 |
|
Bison priscus/Bos primigenius |
1 |
– |
– |
– |
1 |
14 |
9 |
25 |
|
Identified to the family level (ZooMS) |
||||||||
|
Bovidae |
– |
– |
– |
– |
– |
2 |
– |
2 |
|
Bovidae/Cervidae |
1 |
– |
– |
– |
– |
– |
1 |
2 |
|
Saiga/ Cervidae |
– |
– |
– |
– |
– |
1 |
1 |
2 |
|
Ovis/Saiga/ Cervidae |
– |
1 |
– |
– |
– |
– |
– |
1 |
|
Ovis/Capra/Saiga/ Cervidae |
– |
3 |
– |
1 |
– |
– |
– |
4 |
|
Total |
5 |
9 |
3 |
12 |
3 |
62 |
21 |
115 |
teeth of Equus ferus and Capra sibirica ; horizon 3.1 contains Ovis ammon .
ZooMS-analysis
The ZooMS analysis was carried out for the selected unidentifiable bone fragments over 20 mm in size, found in situ . In total, 122 samples were taken from layers 7–1.
94 samples were identified during the study (see Table). All the samples show a high degree of taxonomic identification. For 27 samples from layers 7.2–4.1, six to nine peptides with m/z were identified: 1105.6; 1182.6; 1198.6; 1427.7; 1550.8; 1649.8; 2145.1; 2883.4; and 2899.5, which suggest their attribution to Eguus sp. Among the identifiable bones from the same layers, remains of Equus ferus and Equus hemionus were recorded; hence, the bones under analysis belonged precisely to these equine species. The 25 samples collected from layers 7.2– 5.2 represented large animals of the Bovidae family; these showed from 6 to 11 peptides with m/z: 1105.6; 1192.6; 1208.7; 1427.7; 1580.8; 1648.8; 2131.1; 2792.3; 2853.4; 2869.4; 3017.5; and 3033.5. Taking into account the paleo-landscape reconstructions, in the Pleistocene this territory might have been inhabited by Bison priscus (steppe bison) and Bos primigenius (primitive aurochs) (Kozhamkulova, 1969: 79). The paleontological collection also includes teeth of Bison priscus from horizons 6.8 and 7.1; most likely, the remains of Bovidae belonged to this species. Seven samples revealed peptides with m/z: 1105.6; 1196.6; 1427.7; 1580.8; 1648.8; 2131.1; 2883.4; 2899.4; 3017.5; and 3033.5, which suggests genus Ovis. 19 samples, containing from 6
to 10 peptides with the m/z values of 1105.5; 1180.6; 1196.6; 1427.7; 1580.8; 1648.8; 2131.1; 2792.1; 2883.4; and 2899.4, indicate Ovis/Capra. The bones of Ovis/Capra were recovered from layers 7.1–2.1. According to regional reconstructions, Ovis ammon and Capra sibirica dominated in the local fauna during the Upper Pleistocene (Kozhamkulova, 1981). Four bone samples (three from horizons 2.2 and 2.1, one from horizon 4.2) were identified as Ovis/Capra/ Saiga/Cervidae by 6–8 peptides with m/z: 1105.6; 1180.6; 1196.6; 1427.7; 1648.7; 2131; 2883.4; and 2899.4. One specimen from layer 2 was identified as Ovis/Saiga/Cervidae; it reveals a set of peptides with m/z: 1105.6; 1180.6; 1196.6; 1427.7; 1648.8; 2131.1; 2883.1; and 3017.7. Two samples from layers 7.1 and 1 contained peptides with m/z: 1105.6; 1427.7; 1648.8; 2131.1; 2792.4; 2883.4; 2899.4; 3017.5; and 3033.5, and were identified as Cervidae/Bovidae. Three samples from layers 6.5 and 7.1 contained peptides with m/z: 1105.6; 1221.7; 1443.7; 1550.8; 1634.8; 2131.1; and 2,883.4 which showed the Camelidae. One sample from layer 4.2 contained peptides with m/z: 1105.7; 1217.7; 1233.7; 1453.8; 2163.2; and 2957.8, corresponding to Ursus sp. During the Late Pleistocene, the bears in the region could have been represented by Ursus arctus (brown bear) or U. Savini (cave bear). One sample from horizon 6.4 contained peptides with m/z: 1105.5; 1182.6; 1198.6; 1427.7; 1649.7; 2145; and 2869, and referred to rhinoceros. According to regional reconstructions, it may have been Coelodonta antiquitatis (woolly rhinoceros). Two samples from horizons 6.5 and 6.3 contained peptides with m/z: 1105.6; 1427.7; 1580.7; 1648.8; 2131.1; and 2883.4, which are characteristic of Bovidae. The morphological analysis has shown that these tubular bones, with a significant thickness of the diaphysis walls, belonged to large representatives of the Bovidae family. Two samples from horizons 6.5 and 7.1 contained peptides with m/z: 1105.6; 1180.6; 1196.6; 1427.7; 1550.8; 1648.8; 2131.1; and 2883.4, which may correspond to Alces sp./Cervus elaphus/ Saiga sp./Capreolus capreolus. According to the data on the paleoecological situation, Cervus elaphus (red deer) and Saiga (saiga) could have inhabited the region during that period (Ibid.).
The results of the ZooMS-analysis provide a significant supplement to the data on the faunal collection of the site (Fig. 4). For layers 7 and 6, a much larger proportion of remains of primitive bison was recorded, in comparison with the overlying
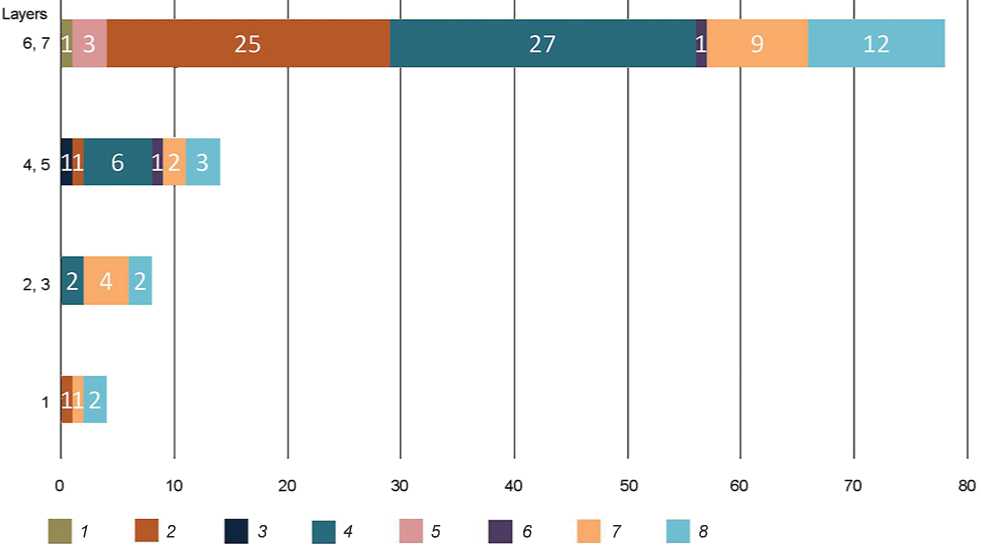
Fig. 4. The composition of the identifiable fauna established through taxonomic and ZooMS analysis in the main lithological units.
1 – Coelodonta sp.; 2 – Bison priscus/Bos primigenius ; 3 – Ursus sp.; 4 – Equus sp.; 5 – Camelus sp.; 6 – Capra sibirica ; 7 – Ovis sp.;
8 – Ovis/Capra .
deposits; these remains, together with horse bones, occur in almost all horizons of these layers. The materials analyzed contain an insignificant share of bones of small ungulates ( Ovis/Capra ). Remains of previously unreported large ungulate mammals (camel and woolly rhinoceros) have been recorded. Layer 4 also contains numerous remains of horse and, probably, argali. A bone from a brown or cave bear of the species that had not been previously noted at the site was found. The faunal composition of the Upper Pleistocene layers 3 and 2 is dominated by the remains of sheep (argali?), and almost lacks bones of larger ungulates. Layer 1, attributed to the Holocene, demonstrates a similar situation, according to the ZooMS data, but its faunal complex might have also included domesticated animals.
Mineralogical and geochemical studies
A sample of 13 bones and teeth of ungulate mammals from different archaeological horizons of layers 7–1 were subjected to mineralogical and geochemical analyses. Chronologically, these horizons are fairly evenly distributed in the range between early MIS 3 and MIS 1.
The microstructure of the remains, the composition of the illuviated mineral impurities, epigenetic minerals, the composition of microelements, the crystalo-chemical properties of bioapatite, the thermal properties, and the elemental and amino acid composition of the bone collagen were analyzed. The patterns of changes in the properties of fossil bones in the process of their fossilization were revealed; the isotopic compositions of C and O in bioapatite, and C and N in bone collagen, were analyzed; paleoclimatic and paleoecological reconstructions were proposed (Silaev et al., 2022).
The bone remains (fragments of tubular bones, cervical vertebrae, and teeth of the upper jaw of argali, kulans, and wild horses) under study revealed 52 microelements, including 12 essential elements, 18 physiologically active elements, and 22 antibiont elements. The total concentration of the microelements varies in the range of 419– 2711 g/t, growing up with the age of the fossils. The proportion between the group concentrations of essential elements and antibionts in the bones under study varies from the youngest to the oldest in the range of 6.54–0.08. This pattern is confirmed by the ratio of the content of essential zinc to the content of physiogenically active copper, which varies from
33.33 to 0.35. Thus, the proportion between essential microelements and antibionts is a reliable criterion for assessing the degree of fossilization and, accordingly, the relative age of fossil bones. In addition, it further confirms the consistent deposition of paleontological and archaeological materials at the site. Another reliable indicator is the total concentration of lanthanides accumulating in the bones as a result of their interaction with the enclosing soils. It varies from 4 g/t in the youngest bone to 171–188 g/t in the oldest ones. The degree of crystallinity of bone bioapatite, determined by X-ray diffraction patterns and IR-spectroscopy data, also shows the correct chronological sequence. As the organics become older, the values of X-ray crystallinity indices increase by 55–100 %, i.e. the degree of crystallinity of bioapatite increases 1.5–2 times (Ibid.).
The elemental composition of collagen in the studied bones (wt%) is: C = 60–65; N = 20–25. The atomic ratios of C/N are 3.39–3.91, which suggests satisfactory preservation of the chemical composition of bone organic matter. The revealed isotopic composition of carbon and nitrogen in bone collagen indicates that the analyzed samples generally correspond to herbivorous animals. The most ancient animals lived in a relatively cold and dry climate in meadow-steppe and forest-steppe landscapes; their diet was based on plants of the C3 type. The younger populations were steppe animals with a mixed C3– CAM diet; they lived in a relatively cool climate. The even younger mammals lived in the warm climatic conditions of the transition from steppe to savannah environments. Finally, the population of the Early Holocene existed in steppes and savannahs with a warm arid climate; it had a transitional diet from CAM to C4 (see Fig. 3, 2 ).
Discussion and findings
Information on the current environmental conditions in the area of the site will add to a better understanding of paleogeographic reconstructions. According to the Köppen classification (Beck et al., 2018), the climate here is identified as cold steppe (Bsk). The climate of the Shilikta Valley is desert-steppe dry, while in the surrounding mountains, including the plateaulike tops of the Saur ridge, the climate corresponds to the alpine tundra-meadow zone. In the valley, summers are dry and hot, winters are cold and severe. The average annual precipitation is 281 mm; the average annual air temperature is -4 °C. The main mammal species recorded in the Pleistocene layers of the site are argali, Siberian ibex, brown bear, and others; these species still inhabit the region nowadays. Domesticated horses, bulls, and camels are also present.
The data of the litho-chemical and mineralogical-geochemical analyses correlate well with each other; on the basis of these data and the established chronology and stratigraphy of the site, the following paleoecological reconstructions can be proposed (see Fig. 2).
The earliest stage (~52–37 ka BP) falls in the Middle Zyrian (Middle Valdai within Moershufd-Hengelo) Interglacial, with a moderately warm and relatively humid climate. Unfortunately, the available data do not provide sufficient information to identify any smaller fluctuations of climate within this long period. At the same time, these data show a general trend of some increase in average annual temperatures, and possibly an increase in aridity. It is generally believed that the composition of animal communities in northeastern Kazakhstan at that time was similar to the classic mammoth faunal complex (Kozhamkulova, 1981; Kozhamkulova, Pak, 1988). The complex includes woolly rhinoceros and bison/ aurochs; fossils of these animals were recovered from the lowermost layers of the site. Bone remains of kulan/wild horse and camel, which species are characteristic of the Late Pleistocene fauna of the northern parts of Central Asia, have also been recorded. According to the Pleistocene fauna of the region, the latter was most likely Knobloch’s camel ( Camelus knoblochi ), which was widespread in Kazakhstan, southern Transbaikalia, and northwestern China at that time (Klementiev et al., 2022). Taking into account the climatic settings, the data from the mineralogical and geochemical analysis, and the species composition of the fauna, it can be assumed that forest-steppe, meadow-steppe, and semi-desert landscapes existed in the valley during that period.
The second stage began with a break in sedimentation and the probable destruction of some deposits in the period preceding the formation of horizon 5.1. This stage is represented by layers 5.1– 4.1 and corresponds to the range of ~25–21 ka BP. This period is characterized by a transition from relatively warm and possibly humid environments to the coldest settings identified at the site. According to climatic reconstructions for the northern regions of Kazakhstan, it was a period of gradual aridization (Kozhamkulova, 1981; Kozhamkulova, Pak, 1988). Climatically, this was a transition from the terminal stages of the Lipovsko-Novoselovo (Bryansk/ Denekamp) interstadial to the early stages of the Last Glacial Maximum. The deposits corresponding to the cooling peak were probably not preserved in the site’s sequence. The faunal composition remained virtually unchanged; only the remains of camel are missing, which may be due to the change in hunting strategies during this period, since local taphocenosis was most likely formed by the anthropogenic activities (absence of gnawing marks on the bones and predator remains s). During that period, the site was surrounded by the predominantly steppe landscapes. The occurrence of bear bones in the faunal collection should not be considered the evidence of forest taphocenoses, since the brown bear inhabited the Eastern European and Kazakh steppes as late as in the 16th–18th centuries (Kozhamkulova, Pak, 1988). Perhaps, during the LGM, the climate in the Shilikta Valley corresponded to the Dwc type (subarctic continental climate). Notably, horizon 4.1, which is associated with the coldest conditions, contains almost no archaeological material.
The third stage, identified at the site after a certain sedimentation hiatus (layer 3, ~18–16 ka BP), corresponds to the transition from the LGM to the late glacial interstadial, with its relatively cool and dry climate, when temperature and humidity gradually increased. The available data do not provide enough information on the faunal composition, but it had hardly undergone any radical changes. Most likely, it corresponded to steppe and forest-steppe environments.
The period of sedimentation of layer 2 (~15– 14 ka BP) is marked by one of the warmest climatic conditions recorded at the site in the Pleistocene. Climatically, this was the Belling-Allered/Nyapan/ Sopkey interstadial. The identified species do not give an idea of the entire faunal diversity; however, the paleontological collection contains remains of inhabitants of open steppe landscapes (horse), as well as mountain and foothill belts (Siberian mountain goat). Landscape was close to modern, possibly with a greater proportion of forest communities.
The last, Early Holocene, stage was characterized by a moderately warm and humid climate, with a transition from steppe and forest-steppe to savannatype landscapes. The fauna includes steppe and foreststeppe species, as well as the mountain belt species.
In general, the data obtained through various methods correlate well both with each other and with the reconstructions of the Late Pleistocene environments proposed for other regions in the north of Central Asia.
Conclusions
The research done has provided the first paleogeographic reconstructions for the southern part of East Kazakhstan. These show a gradual cooling and general aridization of the climate in this area during the period corresponding to MIS 3, especially at its terminal stages; more favorable conditions were determined during the last late-glacial interstadial and the transition to the Holocene. The faunal collection of the Shilikta Valley generally corresponds to the mammoth fauna of steppe and forest-steppe landscapes, and shows a stable composition of the main species during the second half of the Late Pleistocene. This indicates a high degree of sustainability and adaptability of various species of ungulates to a wide range of climate fluctuations and abrupt environmental changes. At the initial stages of the Upper Paleolithic (layers 7 and 6), hunted mainly large ungulates, such as bison/aurochs, horse, kulan, and camel; bones of these animals predominate in the sediments. The medium-sized ungulates (argali, mountain goats) were hunted too, but these remains are less abundant. Subsequently, medium-sized ungulates became the main hunting purpose, while the proportion of large prey decreased markedly. The site was intensely used in the early stages. This is reflected in the quantity and composition of stone industry, as well as in the amount and diversity of faunal materials. At this stage, the climatic and landscape conditions were optimal for ancient human habitation, including available sources of stone raw materials and stable animal hunting.
Acknowledgements
References Late Pleistocene environments of East Kazakhstan (based on Ushbulak site materials)
- Anoikin A.A., Pavlenok G.D., Kharevich V.M., Taimagambetov Z.K., Shalagina A.V., Gladyshev S.A., Ulyanov V.A., Duvanbekov R.S., Shunkov M.V. 2019 Ushbulak—A New Stratified Upper Paleolithic Site in Northeastern Kazakhstan. Archaeology, Ethnology and Anthropology of Eurasia, vol. 47 (4): 16–29.
- Beck H.E., Zimmermann N.E., McVicar T.R., Vergopolan N., Berg A., Wood E.F. 2018 Data Descriptor: Present and future Köppen-Geiger climate classifi cation maps at 1-km resolution. Scientifi c Data, vol. 5. Art. No. 180214. doi: 10.1038/sdata.2018.214
- Brown S., Hebestreit S., Wang N., Boivin N., Douka K., Richter K.K. 2020 Zooarchaeology by Mass Spectrometry (ZooMS) for bone material. Acid insoluble protocol. doi: 10.17504/protocols.io.bf43jqyn
- Buckley M., Collins M., Thomas-Oaies J., Wilson J.C. 2009 Species identifi cation by analysis of bone collagen using matrix-assisted laser desorption/ionisation time-of-fl ight mass spectrometry. Rapid Communications in Mass Spectrometry, No. 23: 3843–3854.
- Cox R., Lowe D.R., Cullers R.L. 1995 The influence of sediment recycling and basement composition on evolution of mudrock chemistry in the Southwestern United States. Geochimica et Cosmochimica Acta, vol. 59: 2919–2940. doi.org/10.1016/0016-7037(95)00185-9
- Kharevich V., Kharevich A., Pavlenok G., Bocharova E., Taimagambetov Zh., Anoikin A. 2022 Ten millennia without the Levallois technique: Primary knapping methods in Initial Upper Paleolithic industries at the Ushbulak site, eastern Kazakhstan. Archaeological and Anthropological Sciences, vol. 14: 207. doi.org/10.1007/s12520-022-01672-6
- Klementiev A.M., Khatsenovich A.M., Tserendagva Y., Rybin E.P., Bazargur D., Marchenko D.V., Gunchinsuren B., Derevianko A.P., Olsen J.W. 2022 First documented Camelus knoblochi Nehring (1901) and fossil Camelus ferus Przewalski (1878) from late Pleistocene archaeological contexts in Mongolia. Frontiers in Earth Science, vol. 10. Art. No. 861163. doi: 10.3389/feart.2022.861163
- Kozhamkulova B.S. 1969 Antropogenovaya iskopayemaya teriofauna Kazakhstana. Alma-Ata: Nauka.
- Kozhamkulova B.S. 1981 Pozdnekaynozoyskiye kopitniye Kazakhstana. Alma-Ata: Nauka.
- Kozhamkulova B.S., Pak T.K. 1988 Pozdnepleistotsenovaya teriofauna stoyanki Aktas (Kokchetavskaya oblast) i yeyo sopostavleniye s odnovozrastnoy faunoy Vostochnogo Kazakhstana. In Mezhregionalnoye sopostavleniye mezozoy-kainozoiskikh faun i fl or Kazakhstana. Alma-Ata: Nauka, pp. 121–154. (Materialy po istorii fauny I fl ory Kazakhstana; vol. 10).
- Nesbitt H.W., Young G.M. 1982 Early Proterozoic climates and plate motions inferred from major element chemistry of lutites. Nature, vol. 299: 715–717.
- Shnaider S.V., Abdykanova A., Baranova S.V., Sryvkina Y.V., Ershova O.V., Alisher kyzy S., Chernonosov A.A. 2022 Primeneniye ZooMS-analiza k materialam stoyanki Aygyrzhal-2 (Narynskaya dolina, Kyrgyzstan). In Problemy arkheologii, etnografii, antropologii Sibiri i sopredelnykh territoriy, vol. XXVIII. Novosibirsk: Izd. IAET SO RAN, pp. 400–405.
- Silaev V.I., Anoikin A.A., Pavlenok G.D., Khazov A.F., Filippov V.N., Kiselyova D.V., Shanina S.N., Makeev B.A., Urazova K.K. 2022 Kostniye ostatki mlekopitayushchikh s paleoliticheskoy stoyanki Ushbulak (Severo-Vostochniy Kazakhstan): Arkheologicheskiy kontekst, mineralogo-geokhimicheskiye svoystva i paleoekologicheskiye rekonstruktsii. Vestnik geonauk, No. 4 (328): 3–37.
- Silaev V.I., Ponomarev D.V., Kiseleva D.V., Smoleva I.V., Simakova Yu.S., Martirosyan O.V., Vasil҆ev Е.А., Khazov A.F., Tropnikov E.M. 2017 Mineralogical-geochemical characteristics of the bone detritus of Pleistocene mammals as source of paleontological information. Paleontological Journal, vol. 13 (13): 1395–1421.
- Ulyanov V.A., Kurbanov R.N., Semikolennykh D.V., Pavlenok G.D., Kharevich V.M., Anoikin A.A. (In press)
- Etapy razvitiya relyefa v rayone paleoliticheskoy stoyanki Ushbulak (Vostochniy Kazakhstan). Geomorfologiya I paleogeografi ya.
- Welker F., Hajdinjak M., Talamo S., Jaouen K., Dannemann M., David F., Julien M., Meyer M., Kelso J., Barnes I., Brace S., Kamminga P., Fischer R., Kessler B.M., Stewart J.R., Pääbo S., Collins M.J., Hublin J. 2016 Palaeoproteomic evidence identifies archaic hominins associated with the Châtelperronian at the Grotte Du Renne. Proceedings of the National Academy of Sciences of the United States of America, iss. 113 (40): 11162–11167. doi:10.1073/pnas.1605834113
