Лесная селекция в России: достижения, проблемы, приоритеты (обзор)
")
Автор: Тараканов В.В., Паленова М.М., Паркина О.В., Роговцев Р.В., Третьякова Р.А.
Журнал: Лесохозяйственная информация @forestry-information
Рубрика: Лесная селекция и генетика
Статья в выпуске: 1, 2021 года.
Бесплатный доступ
Рассматриваются современное состояние и достижения лесной селекции в России. Дан детальный обзор реализованных в стране селекционных программ и исследований. Сформулированы основные проблемы, препятствующие повышению эффективность лесной селекции, а также приоритетные направления исследований по лесной селекции в России на ближайшую перспективу. Предложены конкретные шаги для наращивания потенциала и исследований в этой области.
Лесная генетика и селекция, единый генетико-селекционный комплекс (егск), генетическое разнообразие, популяционная структура видов, плантационные леса
Короткий адрес: https://sciup.org/143175747
IDR: 143175747 | УДК: 630.165.6: | DOI: 10.24419/LHI.2304-3083.2021.1.09
Forest tree breeding in Russia: achievements, challenges, priorities (overview)
The current state and achievements of forest selective breeding in Russia are considered in the article. A detailed overview of the tree breeding and research programs implemented in the country is provided. The main problems that reduce the efficiency of forest selection are formulated, as well as priority research directions on forest tree breeding in Russia for the near future. Concrete steps are proposed for capacity building and research in this area.
Текст научной статьи Лесная селекция в России: достижения, проблемы, приоритеты (обзор)
В формировании и реализации научно обоснованной стратегии развития лесной селекции и лесного семеноводства важнейшую роль играет наличие надежной и своевременно предоставляемой информации о состоянии лесных генетических ресурсов и уровне организации лесной селекции и семеноводства в стране. В России на управленческом уровне ощущается дефицит актуальной информации в области лесной генетики и селекции, а также отсутствие всеобъемлющей аналитической оценки современного состояния и тенденций развития лесной селекции.
Работа по селекционному улучшению деревьев в России ведётся на объектах единого генетико-селекционного комплекса (ЕГСК), которые созданы в масштабе всей страны с учётом лесосеменного районирования. Эти работы осуществляют организации, подведомственные Рослесхозу, институты Российской академии наук и некоторые вузы. Кроме того, селекционные работы с древесными и кустарниковыми видами ведут сотрудники ботанических садов и дендрариев, а также владельцы частных коллекций древесных растений. Сдерживающее влияние на развитие лесного селекционного семеноводства оказывает проблема «ранней диагностики», обусловленная длительностью онтогенеза и поздним вступлением в семеношение лесообразующих пород бореальных лесов, а также резкое снижение уровня финансирования селекционных программ после распада СССР.
В апреле 2019 г. Российская Федерация приступила к реализации программы развития генетических технологий1. Основной целью Федеральной научно-технической программы развития генетических технологий на 2019– 2027 годы является «комплексное решение задач ускоренного развития генетических технологий, в том числе технологий генетического редактирования, и создание научно-технологических заделов для медицины, сельского хозяйства и промышленности, а также совершенствование мер предупреждения чрезвычайных ситуаций биологического характера и контроля в этой области». Программой предусмотрены следующие направления: биобезопасность и обеспечение технологической независимости; генетические технологии для развития сельского хозяйства; генетические технологии для медицины; генетические технологии для промышленной микробиологии. К сожалению, на данный момент развитие генетических технологий в области лесной генетики и селекции, за которые отвечают Минприроды России/Рослесхоз, не включены в Программу.
Тем не менее заинтересованность Российской Федерации в развитии современных научных генетических технологий требует оценки состояния лесной генетики и селекции, выделения актуальных целей и задач, обоснования перспективных направлений развития новейших технологий в гармонии с развитием классических методов лесной селекции.
Цель исследования – обзор достижений лесной селекции в Российской Федерации за последние 20–30 лет, а также определение конкретных шагов для наращивания потенциала и исследований в этой области.
Методы лесной селекции
В России наибольшее развитие получили такие методы лесной селекции, как отбор географических происхождений и популяций (популяционная селекция); массовый отбор по фенотипу плюсовых деревьев в сочетании с их индивидуальным отбором по потомству; клоновая селекция; межвидовая гибридизация; селекция на гетерозис и др. Своеобразие лесов России заключается в обилии естественных популяций лесообразующих видов и их сравнительно невысокой продуктивности, особенно в северных районах. В связи с этим проблема селекционного улучшения деревьев более актуальна для интенсивно эксплуатируемых и продуктивных южно-таёжных и лесостепных районов страны.
Отбор географических происхождений – один из наиболее ранних способов изучения наследственных различий между насаждениями (популяциями), произрастающими в разных частях ареала, и выявления перспективных клима-типов. Он активно реализуется в России с конца XIX в., но результаты этого метода еще не изучены в связи с относительно небольшим возрастом большинства географических культур.
Популяционная селекция базируется на отборе и изучении генетического разнообразия лучших естественных (плюсовых) насаждений и гарантирует устойчивый, надежно прогнозируемый селекционный эффект при сохранении высокого уровня генетической изменчивости насаждений, свойственного природным популяциям. Схема популяционной селекции в Российской Федерации предусматривает: 1) отбор плюсовых насаждений и закладку на их основе «популяционных» постоянных лесосеменных участков (ПЛСУ); 2) испытание плюсовых насаждений по потомствам и отбор среди них «элитных», разграничение заложенных ПЛСУ на «плюсовые» и «элитные»; 3) дальнейшую селекцию на базе «элитных» насаждений и ПЛСУ. Данный подход имеет преимущество в плане сохранения высокого уровня полиморфизма и задействования эволюционно сложившихся механизмов генетического гомеостаза [1, 2]. Принципиально важно, что успешность отбора лучших популяций по их потомствам не зависит от соотношения аддитивных и неаддитивных компонентов генетической дисперсии. Выделенные по результатам испытаний «элитные» популяции и созданные на их основе ПЛСУ могут сразу использоваться как семенные заказники для производства сортовых семян, что экономит одно поколение отбора.
В России в рамках популяционной селекции должны проводиться испытания экотипов (из сухих, свежих, влажных условий), плюсовых и обычных высокопродуктивных насаждений, блоков лесосеменных плантаций (ЛСП) и ПЛСУ [3]. Однако данных о таких испытаниях пока недостаточно [4]. Своеобразной разновидностью этого метода можно считать адаптивную селекцию. Она направлена на выведение сортов-популяций, обладающих высоким адаптивным потенциалом, т.е. устойчивостью к неблагоприятным климатическим условиям, различным биотическим и абиотическим стрессам [5]. В этом случае отбор осуществляется с учётом взаимодействий «генотип – среда» по адаптивно важным признакам. Для России с ее огромной территорией, разнообразием природных зон, существенной экологической гетерогенностью местообитаний данное направление также является перспективным.
Программы по селекционному семеноводству в России базируются преимущественно на массовом отборе плюсовых деревьев в естественных насаждениях с последующим генетическим изучением отобранных по фенотипу плюсовых деревьев в испытательных культурах и предусматривают следующую последовательность действий: селекционная инвентаризация насаждений с выделением плюсовых насаждений и плюсовых деревьев; создание архивов клонов и прививочных маточных плантаций плюсовых деревьев; создание лесосеменных плантаций первого порядка семенным и вегетативным потомством плюсовых деревьев; создание испытательных культур семенным потомством плюсовых деревьев, получаемым от их свободного опыления; отбор элитных деревьев по итогам испытания плюсовых деревьев в испытательных культурах; создание лесосеменных плантаций второго порядка вегетативным потомством элитных деревьев. Для каждого из обозначенных этапов (объектов) разработаны соответствующие нормативные документы [3]. В дальнейшем запланированы контролируемые скрещивания элитных деревьев.
Выделенные in situ плюсовые деревья и плюсовые насаждения, генетические резерваты и созданные ex situ лесосеменные объекты, а также географические культуры образуют так называемый единый генетико-селекционный комплекс. Объекты, предназначенные для сбора семян (главным образом лесосеменные плантации), а также семенные заказники на базе плюсовых насаждений вместе с постоянными лесосеменными участками образуют постоянную лесосеменную базу (ПЛСБ).
Индивидуальный отбор сельскохозяйственных видов растений базируется на испытаниях отдельных растений по потомствам и анализе родословных, отличаясь модификациями метода у самоопылителей и перекрёстноопыляемых растений. В то время как у перекрёстноопыляемых долгоживущих древесных растений однократный индивидуальный отбор (генетическую оценку) осуществляют тестированием плюсовых деревьев по их полусибовым потомствам в специально создаваемых испытательных культурах. Для этой цели на первых этапах селекции используют сеянцы, выращенные из семян от свободного опыления плюсовых деревьев. В связи с естественным варьированием фона опылителей предусматривается испытание потомств разных генераций, получаемых от урожаев семян в разные годы [6, 7]. Индивидуальный отбор успешен при наличии хотя бы у некоторых плюсовых деревьев высокой общей комбинационной способности (ОКС). При этом у древесных растений имеется возможность длительного сохранения и повторного использования генотипов лучших (элитных) деревьев, выявляемых по результатам тестирования в испытательных культурах, которые локализуются в виде привитых саженцев на лесосеменных плантациях второго порядка (ЛСП-2). Это происходит благодаря длительности онтогенеза и возможности неограниченно долгой репродукции генотипов древесных растений методом прививки. Применение на ЛСП-2 сохранённых генотипов элитных деревьев в большей мере гарантирует воспроизведение в их потомстве планируемого генетического сдвига (рассчитываемого по превосходству над контролем в испытательных культурах), чем при использовании на ЛСП-2 лучших полусибо-вых потомков элитных деревьев (семейная селекция). Такой отбор может быть эффективным даже при отсутствии корреляций между родителями и потомками (при низких значениях аддитивной генетической дисперсии и коэффициента наследуемости в узком смысле).
Семейная селекция отличается отбором для дальнейшей репродукции лучших семей. При этом для репродукции используют не все особи элитных семей, а только лучшие деревья.
Семейная селекция в сочетании с внутрисемейным отбором, как и массовый отбор, эффективна при высокой аддитивной генетической дисперсии признаков, что ограничивает её использование. Преимущество данного метода заключается в скорости получения улучшенных семян, так как испытательные культуры преобразуют в лесосеменные плантации, как правило, изреживанием, а также в большей долговечности и устойчивости деревьев за счёт их семенного происхождения.
В связи с изменчивостью лесообразующих видов по самофертильности/самостерильности у них возможен отбор самофертильных генотипов и создание на их основе инбредных линий [8]. В этом случае могут применяться как классические методы индивидуальной селекции, используемые для самоопылителей, так и селекция на гетерозис.
В связи с длительностью онтогенеза древесных растений и процесса их селекции понятие «сорт» у лесообразующих видов имеет свои особенности [9]. В настоящее время государством зарегистрировано лишь около двух десятков сортов основных лесообразующих видов. Преимущественно это клоны лучших деревьев и межвидовых гибридов лиственных пород. Учитывая большое количество выделенных лесными селекционерами форм и генотипов, отличающихся по комплексу морфофизиологических признаков, потенциал для официальной регистрации сортов лесных древесных растений в России является существенным. Но этот процесс сдерживается сложностью процедуры регистрации, стереотипами мышления лесных селекционеров и экономическими причинами.
Метод межвидовой гибридизации в России, как и во всём мире, оказался особенно перспективным для тополей, у которых возможно вегетативное или микроклональное размножение лучших гибридов [10–12]. Наряду с искусственной гибридизацией различных видов тополя, развивается направление по изучению и введению в культуру ценных естественных гибридов тополей чёрного и лавролистного [13].
Среди хвойных растений межвидовая гибридизация оказалась перспективной у лиственниц
[14, 15]. У хвойных для получения гибридных семян создают двухвидовые плантации из предварительно подобранных клонов/генотипов, обеспечивающих максимальный гетерозисный эффект. Из семян, собранных на таких плантациях, получают посадочный материал для промышленных плантаций2. Перед их созданием осуществляют отбраковку сеянцев. Выявление естественных межвидовых гибридов перспективно также у кедра и других хвойных и лиственных пород [16–18].
Селекция на внутри- и межпопуляционный гетерозис у древесных растений пока находится в стадии экспериментального изучения. Однако судя по успехам в селекции типичного «перекрёстника» – кукурузы, отбор на гетерозис теоретически может стать наиболее эффективным методом селекции лесообразующих видов на продуктивность. Одной из причин медленного развития этого направления является необходимость получения инбредных линий, что у древесных растений проблематично. При случайном подборе пар частота гетерозиса у сосны не превышает 2 %. При скрещиваниях географически не сильно удаленных экотипов более эффективными оказались внутрипопу-ляционные скрещивания [8]. Для повышения частоты гетерозиса необходимо иметь аналоги инбредных линий, в связи с чем предложены различные подходы [8, 19]. Для долгоживущих видов древесных растений крайне актуально развитие методов ранней диагностики гетерозисных растений [16]. Кроме того, для целенаправленного получения гетерозиса перспективным может оказаться метод В.А. Струнникова, основанный на формировании компенсационных комплексов генов (ККГ) при отборе по жизнеспособности на фоне полулетальных мутаций [20]. Модификация этого подхода, предусматривающая, с учётом длительного онтогенеза древесных растений, выявление ККГ у гетерозигот от скрещивания лучших деревьев из местных популяций с генетически депрессивными формами, заслуживает внимания [21]. Таким образом, развитие метода селекции на гетерозис у сосны имеет большие перспективы. Однако для его реализации необходимы дальнейшие исследования.
Еще в ХХ в. в российской лесной селекции начали применять так называемые «нетрадиционные методы»: экспериментальный мутагенез, создание митотических, мейотических и зиготических полиплоидов, исследование культуры клеток и клеточных тканей. Позднее в селекцию лесных древесных растений стали проникать и разработанные на основе развития генетических технологий соматогенез, трансгенез и другие методы генетической инженерии.
В последние годы селекционеры России возлагают определённые надежды на новые технологии, основанные на достижениях в области геномики, эпигенетики и микроразмножения, которые могут ускорить процесс селекции и раскрыть огромный потенциал видов лесных деревьев. Первые работы по геномике в России были сосредоточены на хвойных деревьях, которые являются лесообразующими видами бореальных экосистем Евразии – лиственнице сибирской (Larix sibirica Ledeb.), сосне кедровой сибирской (Pinus sibirica Du Tour) и сосне обыкновенной (Pinus sylvestris L.). Полное секвенирование, сборка и аннотация генома хвойных позволят понять эволюцию этого очень непростого и древнего генома, имеющего гигантский размер, многократно превышающий геном человека. Геномные исследования позволят получить важную информацию и разработать высокоинформативные молекулярно-генетические маркеры, такие как микросателлиты и однонуклеотидные полиморфизмы (SNPs), которые можно будет эффективно использовать для генетического анализа селектируемых признаков и быстрого выявления ценных генотипов, изучения и мониторинга генетической изменчивости хвойных деревьев, их адаптации к изменению климата и для создания селекционных и природоохранных программ. В отечественной селекции этот перспективный метод пока используется преимущественно для генетической паспортизации деревьев и объектов ЕГСК.
Необходимо отметить, что идентификация генотипов деревьев представляет определенную проблему, поскольку на их внешний облик (фенотип) значительное влияние оказывают и условия окружающей среды. Например, точная идентификация сортов и видов Populus по морфологическим или фенологическим характеристикам весьма проблематична – учет 64 признаков по системе идентификации, рекомендованной Международной комиссией IUPOV (International Union for the Protection of New Varieties of Plants), чрезвычайно трудоемок и субъективен. Он возможен лишь при выявлении фенов – генетически обусловленных альтернативных вариаций морфо-анатомических качественных признаков, практически не зависимых в своём проявлении от экологических условий. Такого рода фенетиче-ские исследования пока проведены лишь на сосне обыкновенной и частично на тополях [22–24]. В связи с этим использование молекулярных маркеров для идентификации генотипов лесных деревьев (составления генетических паспортов) является актуальной задачей. Во Всероссийском научно-исследовательском институте лесной генетики, селекции и биотехнологии (ВНИИЛ-ГИСбиотех, ранее – Центральный НИИ лесной генетики и селекции, ЦНИИЛГиС) провели оценку 28 селекционно ценных видов, сортов и гибридов тополя с помощью 12 SSR-маркеров. Были выявлены наиболее полиморфные маркеры, которые могут использоваться для генетической паспортизации и идентификации взрослых деревьев и посадочного материала тополя.
Ядерные микросателлитные SSR-маркеры были успешно применены для идентификации по сохранившимся аутентичным гербарным образцам утерянных генотипов тополя невского
( Populus x newensis Bogd.) и тополя ленинградского ( Populus x leningradensis Bogd.), отличающихся высокой продуктивностью и морозостойкостью на Северо-Западе России [25]. В другой работе аналогичным способом с помощью 8 ISSR-маркеров были идентифицированы быстрорастущие и устойчивые к ложному трутовику Phellinus tremulae генотипы осины, сохранившиеся в коллекции in vitro [26]. Наиболее перспективный из них (‘Кострома’) рекомендован для плантационного выращивания на Северо-Западе России.
С помощью микросателлитных маркеров были генотипированы селекционно ценные образцы берёз (пушистая, повислая, их гибриды, карельская) в насаждениях и в коллекции in vitro . Это позволило определить наиболее полиморфные локусы, которые рекомендуются для генетической паспортизации березы. Молекулярно-генетический анализ двух клонов карельской березы ( Betula pendula var. Carelica ), высаженных в питомник после длительного хранения в культуре in vitro , показал их различия по 4-м микро-сателлитным локусам, что говорит об их разном происхождении. Практическим выводом исследования является то, что данные локусы можно применять для идентификации генотипов этого вида [27]. SSR-маркеры оказались эффективны для оценки таксономического статуса берез ( B. pendula , B. pubescens и их гибриды) в сырых типах леса и болотах Центральной России [28]. Было показано, что гибридами являются 11 % деревьев, в то время как определение видов березы по ключам из определителей флоры часто приводит к ошибочным результатам. Важность идентификации по молекулярным маркерам была показана и на иве ( Salix sp.), которая является ценным источником биомассы. Гибридное происхождение многих культивируемых сортов ивы затрудняет их дифференциацию по морфологическим признакам. Генотипирование 5 видов ивы в коллекции in vitro по микросателлитным локусам позволило выявить генетический полиморфизм и составить генетические паспорта.
Наряду с новейшими технологиями развивается теория В.А. Драгавцева и соавт. [29], согласно которой спектры генов, влияющих на селектируемый признак, меняются под воздействием лимитирующих экологических факторов. Поэтому важнейшее значение при селекции древесных растений приобретает учёт нормы реакции и взаимодействий «генотип – среда». На клоновых плантациях сосны в Алтайском крае оценено разнообразие генотипов по норме реакции годичных приростов древесины на динамику метеофакторов и выделены клоны, устойчивые к некоторым лимитирующим факторам [30].
Целевые породы и направления лесной селекции
Деятельность в области селекции лесных древесных растений в России ограничивается несколькими экономически важными видами деревьев. Наибольшие успехи российской лесной селекции достигнуты по быстрорастущим легкоклонируемым видам – особенно по тополю и иве. Кроме того, в отечественные программы по селекции включены хвойные лесообразующие виды ( Pinus sylvestris , P. sibirica , P. koraiensis , Larix sibirica , L. gmelinii L. sukaczewii , Picea abies , P. obovata , P x fennica , Abies sibirica и др.) и твёрдолиственные ( Quercus robur , Q. mongolica , Acer sp . , Fraxinus sp . ) как наиболее ценные в экологическом и ресурсном отношении, а также мягколиственные ( Betula sp . , Populus tremula , Alnus glutinosa и др.) и некоторые ценные интродуцен-ты ( Psevdotsuga menziesii , Pinus contorta , Betula sp . , Tilia sp . и др.).
В Российской Федерации лесные селекционеры осуществляют отбор основных лесообразующих видов преимущественно на продуктивность (интенсивность роста) и качество стволовой древесины и в меньшей степени по другим признакам. Выбор селектируемых признаков может быть ориентирован на создание культур плантационного типа (быстрорастущих, высокопродуктивных, устойчивых к неблагоприятным условиям среды), мелиоративных, рекреационных и других видов насаждений. В России ведутся также работы по выведению, испытанию и отбору сортов для создания полезащитных лесных полос, насаждений на нарушенных техногенных ландшафтах, насаждений для агролесоводства3. В последние годы в России развивается селекция декоративных видов деревьев для озеленения и ландшафтного дизайна [18].
Для примера перечислим целевые признаки, по которым осуществляется селекция некоторых наиболее важных лесообразующих видов в Российской Федерации:
V Сосна обыкновенная: интенсивность роста и качество стволовой древесины, устойчивость к болезням (корневой губке, фузариозу и пр.), высокая смолопро-дуктивность, засухоустойчивость.
V Лиственницы сибирская, Сукачева, европейская: интенсивность роста и качество стволовой древесины, устойчивость к болезням и вредителям, семенная продуктивность.
V Ели обыкновенная (европейская), сибирская, финская ( Picea х fennica ): продуктивность и качество стволовой древесины, устойчивость к болезням (корневая губка и пр.).
V Сосна кедровая сибирская (кедр сибирский): высокая семенная продуктивность, ускоренный рост, смолопродуктивность.
V Дуб черешчатый: быстрота роста, устойчивость к засухе, устойчивость к болезням (трахеомикозу, мучнистой росе и т.п.) и энтомовредителям (зеленой дубовой листовертке, желудевому долгоносику и т.п.), форма ствола.
V Тополи: способность к вегетативному размножению зимними стеблевыми черенками, продуктивность и качество стволовой древесины, содержание и качество целлюлозы, декоративность, пылепогло-щаемость, устойчивость, долговечность, корнеотпрысковость; для затопляемых мест – выживаемость в анаэробных условиях и пр.
V Карельская береза: высокая декоративность древесины, быстрота роста, прямизна деловой части ствола, большой диаметр и протяженность делового отрезка древесины (с текстурой), устойчивость к гнили.
Общепринято, что максимальный селекционный эффект можно получить лишь при отборе деревьев по одному хозяйственному признаку, но в действительности помимо основного селектируемого признака, как правило, учитывается устойчивость к болезням и вредителям, интенсивность семеношения.
Реализация программ по лесной селекции
В области развития лесной селекции и сохранения генофондов лесообразующих видов Россия наследует результаты реализации крупнейших государственных программ, разработанных при руководящем участии ВНИИЛГИСбиотех и Всесоюзного НИИ лесоводства и механизации лесного хозяйства (ВНИИЛМ). Они отражены в правительственных постановлениях и нормативных документах по 1) развитию лесного семеноводства; 2) изучению и созданию новых географических культур; 3) разработке лесосеменного районирования. В конце 1980-х гг. во многих субъектах страны были разработаны региональные программы по лесному семеноводству, учитывающие почти 20-летний опыт реализации «Генеральной схемы лесного семеноводства» [2].
Огромным преимуществом России является обилие естественных лесов. С одной стороны, это создаёт хорошие предпосылки для селекции, так как естественные высокополиморфные популяции – «кладезь генов» для отбора. Но, с другой стороны, возникает проблема сохранения естественной генетической гетерогенности популяций при селекции, поскольку любой отбор может привести к обеднению генофонда и, как следствие, снижению устойчивости насаждений. Генетический потенциал популяций древесных растений, аккумулированный в течение их длительной естественной истории, является «мобилизационным генетическим резервом» российских лесов, который должен рассматриваться в качестве стратегически важного государственного ресурса. Поэтому в России, в отличие от западноевропейских стран, «селекционно улучшенные», но генетически обеднённые леса должны занимать ограниченные площади [1, 31].
При разработке в 1970-х гг. первых крупномасштабных государственных (всесоюзных и региональных) программ по плюсовой селекции и сохранению генофондов лесообразующих видов, которые в России (СССР) традиционно называются «программами по лесному семеноводству», задача сохранения естественной генетической гетерогенности популяций не осознавалась в должной мере. По примеру Скандинавских стран предполагалось создавать культуры преимущественно селекционно улучшенным посадочным материалом, а гетерогенность популяций поддерживать большим числом отселектированных генотипов (клонов) на лесосеменных плантациях (ЛСП) [6]. Очевидно, что такой подход неизбежно привел бы к трансформации естественных генофондов и разрушению популяционной структуры видов, что снизило бы устойчивость лесных экосистем.
По мере развития лесной генетики и селекции в рассматриваемых крупномасштабных государственных программах обнаружились другие просчёты. Первый и важнейший из них, частично исправленный в современный период [3], относится к ориентации на приспевающий и спелый возраст плюсовых деревьев и на длительный период их испытания по потомствам, составляющий не менее половины возраста рубки. Эта тенденция исходила из первоначальных планов использования улучшенного посадочного материала для массовой закладки лесных культур, которые по скорости онтогенеза близки к естественным насаждениям. В настоящее время сортовой посадочный материал используется для создания быстрорастущих промышленных плантаций, закладываемых на высоком агрофоне при относительно низкой густоте. Такие плантации дают «урожай» древесины и других ценных ресурсов в значительно более молодом возрасте, чем возраст спелости естественных насаждений. Кроме того, со временем накопились данные об относительно ранней стабилизации рангов и о возможности более ранней диагностики лучших деревьев и потомств [4, 32, 33]. В совокупности это позволило сократить возраст испытаний и, соответственно, ускорить процесс селекции.
Другой серьёзный недостаток государственных программ 1970–1980-х гг. заключался в громоздкости и длительности испытаний плюсовых деревьев по потомствам, обусловленных ориентацией на поздний возраст окончательной оценки при относительно высокой густоте насаждений, а также на приоритете использования потомств плюсовых деревьев от свободного опыления в естественных древостоях. Было доказано, что особенности потомств плюсовых деревьев (ПД) вследствие варьирования фона опылителей могут существенно отличаться от генерации к генерации [6, 7]. В связи с этим был сделан вывод о необходимости испытаний ПД потомствами не менее 3-х различных генераций, полученных от свободного опыления. Таким образом, с учётом статистических требований минимальных объёмов выборки и числа повторностей на каждое отобранное плюсовое дерево планировалось задействовать около 0,5 га испытательных культур [2]. Например, в 2008 г. на 36 тыс. шт. плюсовых деревьев, выделенных в России, площадь испытательных культур должна была достичь 18 тыс. га. В действительности в стране к этому году было заложено менее 1 тыс. га испытательных культур. Столь грандиозный объём запланированных испытаний не был реализован ни в одном регионе России. Элементарные расчёты показывают, что эта проблема устраняется при получении потомств из семян от переопыления клонов плюсовых деревьев на плантациях, проведении поэтапных тестирований с постепенным уменьшением числа испытываемых ПД при снижении возраста окончательной оценки и густоты опытных насаждений [4, 32]. Если же испытательные культуры встраивать в план закладки соответствующих промышленных плантаций, занимая ими часть площадей, то проблема специального поиска необходимых площадей снимается в принципе. Также в этом случае густота культур и другие требования будут идеально соответствовать условиям промышленных плантаций, что снизит негативное влияние взаимодействий «генотип – среда» при промышленном использовании сортов.
Некоторые авторы заостряют внимание на невысокой эффективности массового отбора плюсовых деревьев по фенотипу [8, 18, 34]. Это может быть обусловлено различными факторами: отбором плюсовых деревьев в спелых и приспевающих насаждениях, неоднократно пройденных приисковыми рубками; неадекватным подбором «контроля» в испытательных культурах, взаимодействием «генотип – среда», маскирующим межсемейные различия при испытаниях на экологически гетерогенных участках; испытанием потомств ПД от свободного опыления; низким качеством отбора; невысокой долей аддитивных генетических эффектов [8, 35]. Методики отбора плюсовых деревьев и испытательных культур, как и любые другие, со временем совершенствуются [32]. Но с учетом всего комплекса проблем массовая закладка лесосеменных плантаций 1-го порядка (ЛСП-1) от не проверенных в испытательных культурах и архивах клонов плюсовых деревьев представляется необоснованной.
Несмотря на указанные неизбежные просчёты, обусловленные уровнем развития генетики и селекции на тот период, советские программы по лесному семеноводству, разработанные лесоводами, заложили прочный фундамент для дальнейшего развития лесной селекции в России и странах СНГ. Важно, что первые программы строились с учётом специально разработанного лесосеменного районирования [36]. Это позволило минимизировать риск разрушения исторически сложившейся естественной популяционной структуры видов. Именно благодаря этим программам в настоящее время в России насчитывается 32 061 плюсовое дерево 63-х видов, 5 828,5 га лесосеменных плантаций, в том числе аттестованных – 4 464,7 га (из них 119,0 га ЛСП повышенной генетической ценности, в том числе аттестованных 72,2 га), 14 122,2 га плюсовых насаждений
31-го вида древесных растений, 17 149,2 га постоянных лесосеменных участков 56-ти видов древесных растений, 573,6 га архивов клонов и 210,2 га архивно-маточных плантаций, 785,1 га испытательных культур и 35,1 га популяционно-экологических (типологических) культур4.
Проблемы сохранения и создания объектов лесного семеноводства
В обстановке дефицита бюджета и непрерывной реорганизации лесной отрасли в постсоветский период в лесном хозяйстве страны на государственном уровне осуществлялась преимущественно политика сохранения объектов лесного селекционного семеноводства. В Указаниях по лесному семеноводству [3] были обобщены нормативные документы и приведены некоторые новые положения, учитывающие накопленный положительный опыт. К ним относятся возможность закладки постоянных лесосеменных участков посадкой отборных сеянцев (саженцев), в том числе полусибами плюсовых деревьев, а также выделение новой категории ЛСП – лесосеменных плантаций повышенной генетической ценности (ЛСПпгц). Последние представляют собой лесосеменные плантации, созданные привитыми саженцами лучших плюсовых деревьев, положительно зарекомендовавших себя по результатам предварительной оценки их потомств в испытательных культурах II класса возраста, а также в архивах клонов. Кроме того, очень важным является положение о возможности создания различных по представительству клонов ЛСПпгц и ЛСП-2: 1) типичных многоклоновых – с числом клонов не менее 50; 2) с ограниченным числом клонов; 3) двух- и одноклоновых с высокими показателями специфической и/или общей комбинационной способности (СКС и ОКС соответственно). Это положение сглаживает противоречие между требованием сохранения высокого уровня генетической гетерогенности на ЛСП для обеспечения высокой устойчивости селекционно улучшенных насаждений, с одной стороны, и необходимостью интенсификации отбора для повышения его эффективности – с другой.
В европейской части России в последние десятилетия организованы 131 га ЛСПпгц [37], в азиатской части планируется создать ЛСПпгц в Новосибирской обл. на площади 10 га [38]. Недостаточные объемы созданных ЛСПпгц обусловлены небольшим количеством испытательных культур и резким снижением финансирования отрасли в последние десятилетия.
Состояние большинства объектов лесного селекционного семеноводства страны постепенно ухудшается. Лесная селекция и связанные с ней процессы сохранения и пополнения ценного генофонда лесообразующих видов – очень динамичные области деятельности. Несмотря на длительность онтогенеза древесных пород, плюсовые насаждения и деревья, отобранные преимущественно в спелых и приспевающих древостоях, стареют и отмирают, насаждения на плантационных объектах ЕГСК быстро смыкаются и теряют ценность из-за отсутствия лесо-водственных уходов. Затормозить этот процесс противопожарными мероприятиями и сбором шишек, т.е. исключительно охраной от пожаров и эксплуатацией, на которые в настоящий период выделяются бюджетные средства, невозможно. Это наглядно демонстрируют инвентаризации объектов ЕГСК. Так, после инвентаризации, осуществлённой силами ФГУ «Рослесозащита» в 2007 г., было списано много объектов, особенно в Сибири. Например, в 2010 г. в Новосибирской обл. были исключены из госреестра 27 % плюсовых деревьев и 25 % ЛСП.
Что касается отобранных плюсовых деревьев, то в настоящее время, по нашим оценкам, средний возраст ПД хвойных пород превышает 120 лет, многие из них поражены грибными болезнями [38]. Так, в Сибири в лидирующих по селекционному семеноводству Новосибирской обл. и Алтайском крае фонд плюсовых деревьев по всем видам составляет всего около 600 шт. на регион. По некоторым данным, интенсивность отбора плюсовых деревьев в России в 15–26 раз ниже, чем в соседних Финляндии и Швеции [37]. Тем не менее есть и положительные примеры. Например, в Республике Карелия отобрано 1 695 плюсовых деревьев, преимущественно сосны и ели, что с учетом общей площади лесов, безусловно, ниже, чем в Финляндии, но существенно выше, чем в среднем по России [39, 40]. Интенсивность отбора плюсовых деревьев влияет не только на эффективность селекции по прямому признаку, но и, косвенным образом, на возможность повышения семенной продуктивности и сроков эксплуатации семеноводческих объектов.
Доля семян улучшенной селекционной категории, заготавливаемых на ЛСП, пока не превышает 2–3 % общего объёма заготовок семян древесных растений в России. Несмотря на то что новые ЛСП (после 1990-х гг.) в стране практически не создавались, урожайность плантаций, имеющих невысокий возраст, в принципе постепенно увеличивается и за счёт этого вклад улучшенных семян в общий урожай может повыситься. Однако в результате того, что семенные деревья на ЛСП произрастают по I–III классам бонитета и доступность урожая семян для сбора имеющимися средствами возможна до высоты не более 7–8 м, к 30–40-летнему возрасту от половины и более биологического урожая хвойных пород оказывается недоступной для сбора. Быстрый рост деревьев на семенных плантациях усугубляется их загущенностью и отсутствием изреживаний, что приводит к перемещению урожая в верхнюю, наименее доступную для сбора часть кроны. По этой причине фактический средний многолетний урожай семян за период эксплуатации ЛСП, составляющий около 25 лет, сокращается в 2–3 раза по сравнению с планируемым. Например, среднемноголетний урожай сосновых семян на плодоносящих ЛСП Озёрского лесничества Алтайского края не превышает 2 ц при их площади около 100 га. Это соответствует урожайности около 2 кг/га вместо планируемых 5 кг/га [2].
Изучение динамики роста крон показало, что у таких хвойных пород, как сосна, кедр и лиственница, оптимальная густота древостоя, позволяющая обходиться без изреживаний и частично замедляющая прирост в высоту, составляет не более 70 шт./га [38]. При редком размещении многие деревья формируют низко-опущенную крону, перспективную для позднего интенсивного обезвершинивания, применение которого может продлить срок эксплуатации ЛСП. Этот метод, используемый на некоторых ПЛСУ сосны обыкновенной в аридных условиях горной лесостепи Забайкалья, нуждается в апробации применительно к ЛСП других пород в различных лесорастительных условиях. Достигнуть увеличения семенной продуктивности и, частично, сроков эксплуатации лесосеменных плантаций при редком размещении деревьев можно за счёт дополнительного отбора деревьев на семенную продуктивность. В связи с отсутствием значимой генетической корреляции между этим признаком и интенсивностью роста дополнительный отбор на семенную продуктивность, по крайней мере у такой породы, как сосна, не приведёт к снижению скорости роста [2, 7]. Но интенсивность отбора в этом случае должна быть существенно повышена, о чём мы упоминали выше. Расчёты, проведенные с учётом маскирующих эффектов взаимодействий «генотип – среда» и дополнительного отбора на семеношение, показывают [38], что интенсивность отбора должна составлять не менее 1 тыс. плюсовых деревьев на лесосеменной район в его «старом» понимании [36].
Ещё одной важной проблемой лесного селекционного семеноводства, на которой заостряют внимание многие специалисты, является точность маркировки деревьев на лесосеменных плантациях. В настоящее время при наличии средств эта проблема может быть легко решена с помощью методов ДНК-диагностики. Показано, что у сосны достаточно хорошие результаты даёт также использование методов фенетики. Однако «научные» объекты – архивы клонов и испытательные культуры – для снижения вероятности ошибок в маркировке родословных лучше создавать при научных учреждениях. Задача паспортизации деревьев на ЛСП и в архивах клонов свидетельствует о целесообразности разграничения функций между научными организациями-кураторами по лесной селекции и лесохозяйственными предприятиями (лесничествами, арендаторами) по принципу: «наука выводит сорта, а лесничества и арендаторы используют их для производства сортовых семян и культур» [38]. Постсоветский период восстановления лесного семеноводства начался с инвентаризации объектов ЕГСК (2007–2008 гг.), которая была инициирована руководителем ФБУ «Рослесозащита» М.Е. Кобельковым. Генетические отделы региональных центров защиты леса начали осуществлять работы по генетической паспортизации объектов лесного семеноводства с применением ДНК-маркеров, идентификации патогенов на ЛСП и другим направлениям.
В настоящее время в головном учреждении по лесной селекции ВНИИЛГИСбиотех значительно сократилась численность сотрудников, опорных пунктов и опытных объектов в сравнении с советским периодом и, в соответствии с современными тенденциями, институт ориентирован преимущественно на развитие молекулярной генетики и методов культуры клеток и тканей. В то же время в его распоряжении находится уникальная экспериментальная база, включающая свыше 900 объектов площадью около 10 тыс. га, которые служат полигоном для научных исследований, развития классических и разработки новых методов селекции.
К другим подведомственным Рослесхозу учреждениям, имеющим селекционные объекты, относятся Северный НИИ лесного хозяйства (СевНИИЛХ, г. Архангельск) и система центров защиты леса ФБУ «Рослесозащита».
При сопоставлении площади различных объектов ЕГСК, существующих в Российской Федерации в настоящее время, можно отметить, что за исключением некоторых регионов темпы закладки испытательных культур плюсовых деревьев относительно низкие. Например, по данным А.П. Царева [37], на этот вид объектов в 2013 г. приходилось всего 821 га. Если исходить из минимальных требований к испытаниям плюсовых деревьев в расчёте выращивания 40–100 деревьев на одной делянке и 3-кратной повторности испытаний, то на 1 плюсовое дерево необходимо создать около 0,09–0,05 га испытательных культур. Расчеты показывают, что из 35 тыс. отобранных в стране плюсовых деревьев испытаны по потомствам около 9–16 тыс., т.е. менее половины. Таким образом, самым несовершенным звеном в программах по лесному семеноводству является устаревшая методика создания испытательных культур. Очевидно также, что в связи с длительной окупаемостью программы по лесному семеноводству в России могут осуществляться только при плановой государственной поддержке, которая многократно снизилась после распада СССР.
Достижения последних десятилетий в области лесной селекции
Определённый объем работ по реализации и развитию программ плюсовой селекции, а также оценке генетической структуры видов был проведен в последние десятилетия в разных регионах России силами сотрудников отраслевых и академических институтов, а также вузов. Однако если исходить из определения селекции как науки о выведении сортов, то достижения лесных селекционеров России пока довольно скромны [9]. На 2016 г. в Государственном реестре селекционных достижений было указано всего 13 сортов лесных древесных растений. Большей частью они относятся к быстрорастущим породам (тополь, ива), всего 4 – к хвойным лесообразующим породам, включая кедр сибирский (автор д-р б. н. Е.В. Титов). К настоящему времени сотрудниками ведущего института по лесной генетике и селекции ВНИИЛГИСбиотех получены патенты на новые селекционные достижения, среди них: 3 сорта тополя (автор д-р б. н. А.П. Царёв), а также 2 сорта сосны – патент № 9187 на сорт Сосна ‘Острогожская’, отличающийся засухоустойчивостью (авторы к. б. н. Н.Ф. Кузнецова и О.С. Машкина), и патент № 9188 на сорт Сосна ‘Красавица’ с высокой смолопродуктивностью и устойчивостью к корневым патогенам (автор д-р с.-х. н. А.А. Высоцкий).
К 2019 г. в Центральном Черноземье отсе-лектировано и размножено 12 лучших гибридов тополя, 3 из них получили статус сорта. Это гибриды А.П. Царева ‘Болид’, ‘Ведуга’ и ‘Степная Лада’, на которые Госсортокомиссией Российской Федерации выданы патенты ВНИИЛГИСбиотех и авторские свидетельства селекционеру. Еще один сорт Р.П. Царевой и В.А. Царева – ‘Бриз’ – принят Госсорткомиссией на испытание. Проводится дальнейшая работа по патентованию и других кандидатов в сорта [10]. Выделенные гибриды в 18–30-летнем возрасте превышают по росту и продуктивности средние показатели семей от 30 до 170 %. Опыты по их сортоиспытанию заложены на ряде опытно-производственных участков в Воронежской, Тамбовской, Липецкой, Волгоградской областях. Очевидно, что это направление лесной селекции требует дополнительного финансирования.
В последние десятилетия сотрудниками ВНИ-ИЛГИСбиотех разработаны рекомендации и технологии по созданию плантационных культур карельской берёзы, сохранению и размножению in vitro ценного генофонда лиственных древесных растений. В монографиях сотрудников института обобщен опыт создания ЛСП и географических культур важнейших лесообразующих пород [7, 41, 42], а также разработаны генетико-селекционные технологии, экономический эффект от внедрения которых составляет от 20 до 130 %.
Сотрудниками института СевНИИЛХ изучается потенциальная возможность очень ранней идентификации быстрорастущих деревьев сосны по маркерному признаку – числу семядольных хвоинок.
В европейской части России исследования в области лесного семеноводства осуществляют также Воронежский лесотехнический университет им. Г.Ф. Морозова (ВГЛТУ), Институт леса КарНЦ РАН, Петрозаводский государственный университет, Московский государственный лесотехнический университет (МГЛТУ), Санкт-Петербургский лесотехнический университет (СПб-ГЛТУ), Институт биологии Коми НЦ УрО РАН, Поволжский государственный технологический университет, Южно-Уральский ботанический сад-институт Уфимского федерального исследовательского центра РАН, Пермский государственный национальный исследовательский университет и другие научные и образовательные учреждения.
На Урале и в азиатской части России исследования по лесной селекции осуществляют Институт леса и его Западно-Сибирское отделение, вошедшие в ФИЦ КНЦ СО РАН, Институт мониторинга климатических и экологических систем (ИМКЭС СО РАН), Институт лесных технологий СибГУ им. академика М.Ф. Решетнева (бывший Сибирский государственный технологический университет, СибГТУ), Ботанический сад УрО РАН, Институт экологии растений и животных (ИЭРиЖ УрО РАН), Федеральный научный центр биоразнообразия наземной биоты Восточной Азии (ФНЦ Биоразнообразия ДВО РАН) и др.
Наиболее важные результаты, полученные российскими учреждениями в последние десятилетия:
V выявлена связь морфологических признаков со смолопродуктивной способностью сосны обыкновенной ( Pinus sylvestris L.) и разработаны критерии отбора высокосмолопродуктивных деревьев при рубках ухода в условиях гидроморфных и автоморфных типов леса [43];
V в Горном Алтае на уникальных клоновых плантациях, где сконцентрирован «плюсовый» генофонд кедра сибирского ( Pinus sibirica du Tour), изучена связь габитуаль-ных признаков со смолопродуктивностью потомств плюсовых деревьев, отобранных на скорость роста, семеношение и смо-лопродуктивность; уточнены критерии отбора лучших деревьев по смолопродук-тивности в условиях ЛСП [44];
V проведен сравнительный селекционно-генетический анализ клонов сосны обыкновенной на лесосеменных плантациях Дюртюлинского лесничества Министерства лесного хозяйства Республики Башкортостан; установлены существенные различия клонов по интенсивности цветения, энергии прорастания, всхожести семян и их зараженности [45];
-
V в Нижегородской обл. получены многомерные сравнительные оценки 36 плюсовых деревьев сосны обыкновенной по комплексу показателей содержания запасных веществ (жиров и углеводов) в тканях побегов для определения степени их статистической и генотипической близости, что планируется использовать при подборе пар для гибридизации [46];
-
V в Пермской обл. проведены многолетние исследования по оценке влияния густоты родительских древостоев на высоту ствола полусибовых потомств у ели финской и сосны обыкновенной в испытательных культурах I–II класса возраста разной густоты; на основании исследований огромного количества испытательных культур сделан вывод о наличии у потомков «генетической памяти» об условиях локальной загущенности материнских древостоев; разработаны критерии отбора плюсовых деревьев для создания оптимально загущенных лесосырьевых плантаций; в качестве показателя условий локальной загущенности древостоя на раннем этапе онтогенеза деревьев предложено использовать сбег ствола; разработана методика поэтапной генетической оценки деревьев, сочетающая ускоренное тестирование большого числа деревьев в начале опыта с более длительной проверкой немногих оставшихся деревьев, прошедших отбор на первом этапе [32, 47]. По итогам исследований М.В. Рогозиным разработана Региональная программа по селекции сосны обыкновенной и ели финской [4];
-
V в условиях Новосибирской обл. проведена селекция кедра сибирского на урожайность орехов; выделены высокоурожайные клоны; показано, что плантационные культуры кедра, созданные прививкой от лучших клонов, позволяют получать урожай орехов, в 2–2,2 раза превышающий
урожай лучших природных насаждений кедра [48];
-
V на северо-западе Российской Федерации на основе изучения вегетативного и семенного потомства плюсовых деревьев ели европейской и сосны обыкновенной на лесосеменных плантациях и в испытательных культурах разработана и апробирована методика комплексной оценки генетических свойств плюсовых деревьев; выявлены генотипы, рекомендуемые для использования при создании лесосеменных плантаций повышенной генетической ценности с перспективой создания лесосеменных плантаций второго порядка [49, 50];
-
V на основании анализа литературы и собственных исследований испытательных культур сосны обыкновенной и ели финской в Кировской обл. и Удмуртской Республике сделан вывод о низкой эффективности массового отбора на быстроту роста; указаны возможные причины неоднозначности получаемых прогнозов эффективности массового отбора по фенотипу; намечены пути его дальнейшего развития [34];
-
V продолжены исследования по оптимизации методики испытательных культур и возможности ранней диагностики лучших генотипов [2, 4, 32, 38, 49]. Большинство исследователей сходятся во мнении о необходимости этапности испытаний и сокращения размера семей, возможности применения ранней диагностики по скорости роста. В связи с ориентацией селекционно улучшенного материала на закладку плантаций с коротким оборотом рубки и отрицательной селекцией при лесозаготовках предложено проводить отбор плюсовых деревьев в III классе возраста до проведения проходных рубок;
-
V в Красноярском крае изучается возможность ранней диагностики кедра сибирского на основе связи хозяйственно ценных признаков деревьев, семей и клонов с их характеристиками на стадии
всходов и сеянцев; ведётся непрерывный мониторинг хода роста деревьев основных лесообразующих видов на клоновых и семейственных плантациях, в географических посадках [51];
V в Кемеровской обл. в зоне гибридизации тополей чёрного и лавролистного выявляют и вводят в культуру для дальнейшего изучения и размножения ценные по успешности размножения зимними стеблевыми черенками, скорости роста, качеству ствола и декоративности деревья этих видов, а также их естественных гибридов [13, 24];
V продолжаются исследования по генетической паспортизации и проверке достоверности маркировки деревьев на ЛСП [52, 53]. Для снижения стоимости работ их предложено осуществлять в 2 этапа, используя вначале методы лесной фене-тики, а затем аллозимные или ДНК-маркеры, что снижает затраты генетической паспортизации ЛСП сосны обыкновенной почти на порядок [23, 54, 55].
V при изучении ЛСП-1 хвойных пород в Республике Марий Эл выявлен высокий уровень генетического полиморфизма по различным целевым признакам и ДНК-маркерам, что свидетельствует о перспективности дальнейшего отбора и высоком генетическом потенциале изученных объектов [52, 56, 57]; для утилизации урожая улучшенных семян при недоступности сбора шишек имеющимися средствами предложено использовать метод подпологовых питомников [58].
Как показывает анализ программ по лесной селекции, выполненных в последние годы, наиболее плодотворными и перспективными являются комплексные работы в тех регионах России, в которых на объектах лесного семеноводства вместе работают сотрудники подведомственных организаций Рослесхоза и представители академической или вузовской науки (Новосибирская, Пермская, Кировская, Томская, Архангельская области, Республика Башкортостан, Красноярский край, Республика Карелия, Республика Марий Эл). В этих случаях достигается максимально возможное и результативное научное сопровождение программ по лесной селекции и лесному семеноводству. При этом используются как ранее заложенные в рамках государственных программ объекты «плюсовой селекции», так и объекты, созданные учеными по оригинальным программам с применением нетрадиционных методов.
Хороший пример взаимодействия науки и производства демонстрируют Новосибирская обл. и Алтайский край, лидирующие среди субъектов Сибирского федерального округа по созданию объектов лесного семеноводства видов хвойных деревьев. На территории Новосибирской обл. расположен большой генетико-селекционный комплекс, в котором представлены все хвойные лесообразующие виды Сибири: сосна обыкновенная, кедр сибирский, лиственница сибирская, ель сибирская и пихта сибирская. По результатам регулярных исследований испытательных культур более 100 плюсовых деревьев и географических культур из 37 происхождений сосны, вступивших во II класс возраста, были выделены «предэлитные» деревья, перспективные для создания ЛСП повышенной генетической ценности, и климатипы-кандидаты в «сорта-популяции» [2, 38, 59]. Также идентифицированы клоны кедра сибирского, отличающиеся регулярными высокими урожаями семян, для оформления их в качестве сортов [48]. Этот регион первым в Сибири приступил к разработке и реализации новой Региональной программы селекции, рассчитанной на ближайшие 20 лет.
Значительный объём прикладных и фундаментальных исследований осуществляется на генетико-селекционных объектах сосны в Алтайском крае [2]. Здесь при тесном взаимодействии новосибирских ученых с лесоводами Алтая разработаны технологии создания улучшенных ПЛСУ на генетической основе плюсовых насаждений и «плантационно-обсеменительных культур», апробирована технология массового доопыления молодых насаждений ЛСП. Доказано влияние генетических особенностей деревьев на химический состав почв, выявлены генетические различия и оценена наследуемость различных морфометрических, фенетических, химических и физиологических признаков, оценена устойчивость потомств плюсовых деревьев к техногенным воздействиям и возбудителям фузариоза, а также созданы и изучаются географические посадки ценного интродуцента сосны скрученной (Pinus contorta).
В Республике Алтай сконцентрирован уникальный генофонд плюсовых деревьев кедра сибирского, отобранных на смолопродуктивность, интенсивность роста, а также на урожайность семян.
К сожалению, с 2013 г. по настоящее время насаждения этого ценного вида на территории питомника подвержены новому прогрессирующему заболеванию – дотистромозу, выявленному с применением современных методов ДНК-диагностики [44], опасность которого до сих пор не осознана ни Министерством природных ресурсов и экологии Республики Алтай, ни руководством Центра защиты леса Алтайского края. Очевидно, что программы по селекции и сохранению ценных генофондов лесообразующих видов могут реализовываться только при полной государственной поддержке и согласованной работе всех лесных ведомств.
Один из самых крупных объектов лесного семеноводства хвойных пород и карельской берёзы создан в Республике Карелия, где лесными селекционерами отобрано около 2 тыс. плюсовых деревьев, преимущественно сосны обыкновенной [39, 50, 60, 61].
В регионе отбор лучших плюсовых деревьев рекомендуют осуществлять по комплексной оценке признаков одноименных клонов и семей. Селекционеры полагают, что приемлемый возраст отбора в испытательных культурах сосны обыкновенной, лидирующих по росту семей, составляет 7 лет, т.е. возможна довольно ранняя диагностика. Значительное внимание уделяется также селекции ценного интродуцента – сосны скрученной.
Изучением этого интродуцента занимаются и в Республике Коми [62]. Установлено, что северные экотипы сосны скрученной перспективны для выращивания на юге республики. Они обгоняют в росте местную сосну обыкновенную, но несколько уступают ей по качеству ствола. Ведущим лесным селекционером этого региона А.Л. Федорковым на основе анализа собственных данных и зарубежной литературы сделан вывод о приемлемой эффективности фенотипического (массового) отбора плюсовых деревьев: превышение потомств плюсовых деревьев над контролем в испытательных культурах в пределах 10–25 % по высоте и объёму ствола [35]. Он считает возможным использовать «низкозатратный» метод селекции, заключающийся в непрерывном массовом отборе по схеме «отбор в естественных популяциях – размножение – отбор в плантационных насаждениях – размножение – отбор в плантационных насаждениях …», эффективность которого повысится в условиях одновозрастных плантаций.
Значительный вклад в развитие лесной селекции хвойных и лиственных пород и интродукции ценных экзотов внесён лесными селекционерами Республики Башкортостан [14, 63]. Примечательно, что все работы по лесной селекции и семеноводству осуществлялись в этом регионе параллельно с изучением популяционной структуры видов. Кроме этого, здесь разработана оригинальная для России технология многоступенчатого вегетативного размножения трудноукореняемых древесных растений [64]. Лесовосстановление на территории ареалов редко встречающихся генетически обеднённых популяций главной породы В.П. Путенихин и Г.Г. Фарукшина предлагают осуществлять смешанным потомством соседних высокоадаптивных ценопопуляций той же породы.
Еще одним впечатляющим примером реализованной в Российской Федерации селекционной программы по кедру сибирскому и другим хвойным является многолетняя совместная работа отдела лаборатории дендроэкологии ИМКЭС СО РАН (Томск), осуществляющего исследования на научном стационаре «Кедр»5, и ООО Сибирская академия деревьев и кустарников6 (ООО «САДиК»).
Коллективами под руководством д-ра б.н. С.Н. Го-рошкевича выполняются селекционные работы преимущественно по хвойным породам (прежде всего по 5-хвойным соснам) в двух направлениях: селекция сортов для промышленных плантаций и селекция декоративных сортов. Из материала, собранного во время многолетних экспедиционных работ, созданы архивы из 700–750 клонов, организованных в несколько логических совокупностей: широтный профиль, долготный профиль, несколько высотных профилей, несколько экологических (лесоболотных) профилей и др.
Наряду с естественными популяциями 5-хвойных сосен изучаются их культивары (сорта) из многих мировых коллекций. Главное внимание при генетико-селекционных исследованиях кедровых сосен сосредоточено на анализе естественного формового разнообразия, гибридизации, в том числе между разными видами, и генетическом анализе различных декоративных признаков, включая «ведьмины мётлы» [65, 66]. Выведенные сорта размножаются преимущественно методом прививки. Томскими лесными селекционерами впервые выведены уникальные декоративные и скороплодные сорта кедра, разработана технология их вегетативного размножения, создан селекционный питомник, где применяют уникальный для лесной селекции метод выращивания семенного потомства культиваров от свободного опыления с последующим отбором.
Здесь испытано большое число экотипов кедрового стланика, выделен сорт-популяция ‘Кунашир’ (происхождение – Южные Курилы); созданы декоративные, низкорослые, скороплодные и другие привойные сорта кедра сибирского, сорта-гибриды хвойных, например кедра сибирского и кедрового стланика (гибриды прямостоячих видов со стелющимися). В содружестве с Российским обществом селекции и интродукции хвойных7 создана уникальная база данных по российским сортам и культиварам хвойных8, в которой представлены около 350 сортов хвойных, выведенных на основе различных видов сосны (Pinus sibirica, P. pumula, P. koraiensis, P. sylvestris), ели (Picea abies, P. оbovata), пихты (Abies sibirica, A. holophylla), лиственницы (Larix sibirica, L. gmelinii, L. decidua) и др.
Существенный вклад в изучение популяционной структуры и организацию лесного семеноводства сосны обыкновенной на популяционной основе в Кировской обл. и Республике Коми, а также в развитие методов фенетической паспортизации деревьев на клоновых плантациях этого вида внесён лесным фенетиком и селекционером А.И. Видякиным [22, 67, 68]. Этот исследователь на примере сосны обыкновенной развил и внедрил в практику лесной генетики и селекции популяционные подходы к оценке и рациональному использованию генетической гетерогенности в масштабе ареала вида, разрабатываемые в нашей стране Н.В. Глотовым и Л.Ф. Семериковым [69]. Впервые были чётко обозначены отдельные популяции сосны обыкновенной, площадь которых составила от нескольких десятков до 100–200 тыс. га. Надёжность дифференцировки популяций, выделяемых с применением фенетических и аллометрических маркеров, подтверждена с использованием ISSR-маркеров [68].
Полученные результаты согласуются с выводом Л.Ф. Семерикова о том, что площадь популяции дуба черешчатого приблизительно соответствует площади лесного урочища или лесного массива в ландшафтном понимании этого термина [69]. Близкие по размерам популяции хвойных пород на Урале были выделены по комплексу качественных признаков С.А. Мамаевым [70] и В.П. Путенихиным9 [14]. К сожалению, данные Л.Ф. Семерикова и Л.И. Видякина о размерах популяций были получены значительно позже разработки программы по географическим культурам, в связи с чем они не могли быть приняты во внимание разработчиками программы.
Итоги реализации государственной программы по закладке и изучению географических культур
Не останавливаясь на истории исследований географической изменчивости, которая достаточно подробно рассмотрена в специальной литературе, перейдём к результатам реализации самой масштабной программы 1972 г. [71]. Инициатор этой программы и один из основателей лесной селекции в СССР, зав. лабораторией лесного семеноводства ВНИИЛМ Е.П. Проказин полагал, что «изучение географической изменчивости наследственных свойств лесных пород ... даёт возможность, во-первых, вскрыть эволюционно-географические закономерности формирования биогеоценозов, во-вторых, выявить локализацию популяций (климатипов), использование семян которых позволяет сохранить или повысить продуктивность лесных насаждений, и, в-третьих, разработать лесосеменное районирование, обеспечивающее эффективное использование географической изменчивости лесных пород в селекционно-семеноводческих целях» [71, с. 1].
В рамках нашей статьи важны две последних задачи. Рассмотрим вначале результаты, полученные по первой из них – отбору лучших популяций (климатических экотипов, или климатипов).
Программа предусматривала поэтапную организацию всего селекционного процесса с отбором вначале лучших популяций, а затем с отбором внутри них лучших деревьев. В этой связи планировалось: «развернуть более широкое исследование климатипов, показавших лучшие результаты в прежних географических культурах, рассматривая их в качестве сортовых, и одновременно заложить опыты по испытанию потомства лучших деревьев климатипа» [71, с. 3–4]. К сожалению, принцип этапности отбора был лишь отчасти реализован на практике. Фактически отбор лучших климатипов и плюсовая селекция начали развиваться в нашей стране параллельно и почти независимо друг от друга [8].
Тем не менее была запланирована программа с испытанием 290–355 географических популяций 6 лесных пород в 106–130 пунктах СССР (таблица). О масштабах её реализации свидетельствуют публикации, в которых обобщены данные по сосне обыкновенной [42, 72, 73]: по этому виду из запланированных к испытаниям 126 климатипов в 37 пунктах на площади 829 га были изучены 113 климатипов в 33 пунктах на площади 545 га; каждый из климатипов испытан в среднем в 11–12 пунктах. На территорию Российской Федерации приходится 23 из 33 пунктов испытаний (69,7 %).
Результаты исследований географических опытов за последние десятилетия освещены в многочисленных публикациях, которые мы не имеем возможности процитировать в связи с ограничениями по объёму статьи.
Планируемые объёмы закладки географических культур по программе 1972 г. [71]
|
Порода |
Пункты заготовки семян (размещения климатипов) |
Пункты испытания климатипов |
|||
|
Число |
Среднее расстояние между пунктами, град. |
Число |
Среднее расстояние |
||
|
пунктов |
По широте |
По долготе |
пунктов |
между пунктами |
|
|
Сосна |
100–125 |
1,5–7 |
4–12 |
30–35 |
Создаются по «в 2–3 раза более редкой координатной сетке», чем пункты заготовки [71] |
|
Ель |
50–60 |
20–25 |
|||
|
Лиственница |
50–60 |
20–25 |
|||
|
Дуб |
50–60 |
20–25 |
|||
|
Кедр |
20–25 |
8–10 |
|||
|
Пихта |
20–25 |
8–10 |
|||
|
Итого |
290–355 |
- |
- |
106–130 |
- |
Наиболее представительные по числу клима-типов и площади географические культуры созданы в Воронежской, Московской, Ленинградской, Мурманской, Вологодской, Псковской, Архангельской, Липецкой, Кировской, Новосибирской и Омской областях, республиках Карелии, Коми и Бурятии, Красноярском и Забайкальском краях.
Один из кураторов программы 1972 г. А.М. Шутяев отмечает, что после изучения географических культур, представленных средними по показателям роста насаждениями, и выделения сортов-популяций предполагалось детальное изучение их полиморфизма. Для этого была разработана соответствующая программа [74], которая пока не воплощена. В сложившихся экономических условиях предложено осуществлять её на региональном уровне [75]. На наш взгляд, в этой связи перспективна организация «региональных» географических культур на основе плюсовых насаждений лучших климатипов. Учитывая, что селекционная инвентаризация во всех регионах будет продолжена, испытание плюсовых насаждений может быть совмещено с испытаниями плюсовых деревьев, что повысит эффективность отбора [2, 8].
Что касается выделения кандидатов в сорта-популяции, то в настоящее время возраст потомств испытываемых географических популяций, заложенных по программам 1972 г. и ранее, превысил 40 лет. Во многих регионах выделены лучшие сорта-популяции (климатипы) сосны, ели, дуба, лиственницы и др. пород, получены и изучаются культуры следующих генераций. Приведём несколько примеров.
По результатам изучения географических культур сосны, ели и лиственницы, созданных в различных лесхозах (лесничествах) Воронежской, Липецкой и Пензенской областей под руководством М.М. Вересина и А.М. Шутяева, были выявлены климатипы, перспективные для выращивания в лесостепи центральной части России. Например, при испытании сосны обыкновенной в Воронежской обл. оказались близки к местному или превосходили его по высоте и запасу древесины 3 климатипа из 60-ти испытуемых (5 % выборки) – Рязанский, Тамбовский и Удмуртский.
В 27-летнем возрасте их запас древесины составлял 247–295 м3/га против 242 м3/га в контроле, а высота ствола – 11,6–12,5 м против 11,5 м (максимальное превышение над контролем по высоте – 8,7 %) [42]. Родительские насаждения отстоят от пункта испытаний на 200–1000 км. Ель и лиственница являются интродуцентами для условий Воронежской обл. Установлено, что потомства некоторых популяций перечисленных выше видов произрастают в лесостепи центральной части России по высшим классам бонитета и могут быть успешно интродуцированы.
Самый крупный в Сибири эксперимент по посадке 83 климатипов сосны обыкновенной был проведен А.И. Ирошниковым и соавт. в Богучанском лесхозе Красноярского края. Его отличал ряд методических особенностей. Во-первых, густота посадки составила 8,9 тыс. шт./га, что существенно выше рекомендуемой 5,3 тыс. шт./га (при схеме посадки 2,5 х 0,75 м) [71]. Во-вторых, испытания осуществлялись в 2-х типах условий – на бедной дерново-подзолистой песчаной почве и более плодородной темно-серой суглинистой. Средние высота и запас стволовой древесины отличаются в этих условиях почти в 2 раза [76]. На дерново-подзолистой почве доля климатипов, потомство которых растёт лучше контрольного Богучанского, оказалась очень высокой и составила 33 %. При этом среднее превышение по высоте ствола в группе лидирующих климатипов над контрольным климатипом достигло 24 %. Однако при оценке доли климатипов, которые одновременно лидируют и на бедной, и на богатой почве, этот показатель снижается до 9,6 % (8 из 83 климатипов).
В засушливой горной лесостепи западного Забайкалья географические культуры сосны обыкновенной создавали из 59 климатипов при сниженной густоте посадки 4,0 тыс. шт./га [77]. В 20-летнем возрасте культур отмечено существенное сужение размаха изменчивости по средней продуктивности потомств в сравнении с исходными насаждениями, косвенно оцениваемое по высоте и составившее 3 (с III по V) и 6 (с Ia по V) классов бонитета соответственно. К лучшим отнесено 9 из 59 испытываемых кли-матипов (15,3 %). По сравнению с контрольным
Заудинским климатипом (Республика Бурятия) высотой 5,4 м (100 %), высота лучшего Сузунско-го (Новосибирская обл.) и худшего Кандалакшского составила 6,3 м (116 % контроля) и 3,3 м (61 % контроля) соответственно.
Ценным объектом для изучения популяционно-географической изменчивости различных видов лиственницы в Московской обл. являются географические культуры Бронницкого участкового лесничества. В посадках 1954–1955 гг. на площади 41,7 га были высажены 6 видов лиственницы из 41 географического пункта СССР (рисунок). В дальнейшем популяционно-видовой состав был расширен до 53 климатипов и 12 видов, в число которых входят: лиственницы европейская (Larix decidua Mill.), польская (L. polonica Racib.), широкочешуйчатая (L. eurolepic Henry), Сукачёва (L. Sukaczewii Dylis), сибирская (L. sibirika Ledeb.), Чекановского (L. Czekanovskii Szaf), даурская (L. dahurica Turcz), амурская (L. amurensis Kolesn.), ольгинская (L. olgensis Henry), Кемп-фера (L. kaempferi (Lamb.) Carriere), курильская (L. kurilensis Maur.) и американская (L. laricina (Duroi) K. Koch). В 2015 г. географические посадки лиственницы достигли 60-летнего возраста, что позволило сделать объективные выводы о том, какие виды и климатипы в наилучшей степени отвечают местным лесорастительным условиям и имеют высокую продуктивность [78].
Согласно полученным данным, в условиях Московской обл. по высоте лидируют климатипы лиственницы европейской, польской и японской, худшие показатели у лиственницы даурской. Лидером по запасу стволовой древесины является лиственница европейская, формирующая насаждения со средним запасом 1 094 м3/га. Последнее место занимает лиственница даурская – 204 м3/га. Самая низкая сохранность деревьев в пункте испытаний зафиксирована у дальневосточных провениенций, а самая высокая – у лиственниц европейской и Сукачёва [79–81].
В географических культурах сосны обыкновенной средней густоты (5,3 тыс. шт./га), созданных в 1976 г. в лесостепи Западной Сибири, сосна из Зеленодольского лесхоза Татарстана в возрасте 30 лет статистически значимо превосходила местный Сузунский климатип: на 24 % по запасу древесины (340 м3/га) и на 4,6 % по высоте ствола (16 м) [59]. Расстояние от пункта испытаний до Зеленодольска превышает 2 тыс. км.
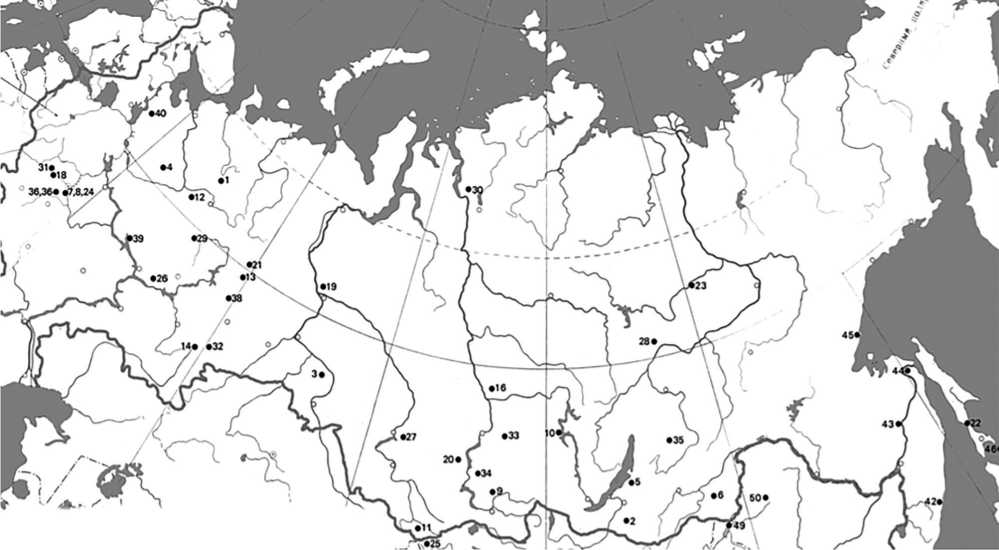
Географическое происхождение экотипов лиственницы в географических культурах Бронницкого лесничества Московской обл. [78]
Эти примеры можно продолжить. Основной вывод из них сводится к тому, что практически в каждом пункте испытаний выделяются происхождения, превосходящие местный климатип, которые могут быть рекомендованы в качестве сорта-популяции. Многие исследователи отмечают перераспределение рангов по интенсивности роста на начальном этапе онтогенеза, но в статье С.Р. Кузьмина и Н.А. Кузьминой [76] подчёркивается усиление этой тенденции в возрасте 30–45 лет. В связи с часто наблюдаемой загущенностью посадки культур возникает вопрос о влиянии данной методической особенности на перераспределение рангов в возрасте, когда наступает усиленная дифференциация древостоев. Отметим, что, за редким исключением, рекомендация по рубкам ухода на части делянок [71] почти нигде не была выполнена. Какое влияние это могло оказать на ход роста климатипов?
Климатипы с высоким уровнем адаптации к условиям пункта испытаний отличаются большей сохранностью, чем менее адаптивные. Вследствие этого при отсутствии рубок ухода со временем из-за повышенной полноты они оказываются в худших условиях по сравнению с менее адаптивными климатипами. Как писал ещё полвека назад П.С. Погребняк (1968): «...максимальная продуктивность вначале принадлежит самой густой посадке, с увеличением возраста она переходит к всё более редким посадкам…»10. Это особенно может проявляться на бедных и более сухих песчаных почвах, что, вероятно, и имеет место в рассмотренном выше красноярском опыте. Детальное изучение влияния начальной густоты на ход роста культур привело к открытию «закона развития одноярусных древостоев» [47, с. 32]: «каждый древостой … достигает предельных состояний развития по сомкнутости крон, запасам древесины, … после чего снижает их тем сильнее, чем выше была начальная густота». Отсутствие своевременных рубок ухода в географических культурах могло существенно повлиять не только на отбор перспективных климатипов, но и на выводы о границах лесосеменных районов.
В отношении последнего напомним, что на основе обобщения комплекса данных о районировании территории и изменчивости исследуемых видов, включая имеющиеся на то время данные по географическим культурам, было разработано и введено в действие Лесосеменное районирование основных лесообразующих пород в СССР11 [36]. В 2015 г., по-видимому на основе обобщения результатов исследований географических культур (публикации в открытой научной печати нами не обнаружены), было разработано и утверждено новое лесосеменное районирова-ние12. Оно отличается от первого варианта значительным укрупнением лесосеменных районов. Насколько оно обосновано?
Если исходить из определения лесосеменного района, которое приведено в капитальной монографии 1982 г. [36, с. 4], инициаторы этих исследований исходили из необходимости учитывать при выделении лесосеменных районов особенности пространственной популяционной структуры видов. О целесообразности ведения лесного хозяйства на популяционной основе писали такие выдающиеся исследователи-лесоводы, как Л.Ф. Правдин, С.А. Мамаев, Л.Ф. Семериков, А.И. Видякин и др. Учитывая адаптивное значение не только генетической гетерогенности отдельных популяций, но и популяционной структуры вида в целом, идеалом должна быть ситуация, когда лесосеменной район тождествен популяции лесообразующего вида. Не случайно разработчик методики выделения и изучения сортов-популяций А.М. Шутяев [74, с. 4–5] даёт определение популяции в понимании этого термина популяционным генетиком-эволюционистом Н.В. Тимофеевым-Ресовским. На момент разработки лесосеменного районирования 1982 г. популяционно-генетическая структура видов-лесообразователей не была изучена должным образом, эта задача не решена в практически приемлемом виде и по сей день. Однако авторы монументального труда, исходя из определяющего влияния особенностей экологических условий и степени изоляции на генетические особенности популяций, разработали приблизительную схему лесосеменных районов с учетом комплекса естественных районирований территории и потенциальных современных изоляционных барьеров в ареале вида в предположении дальнейшего уточнения схемы по мере накопления необходимых данных. В этой связи большая надежда возлагалась и на данные по географическим культурам, заложенным по программе 1972 г.
Однако, не располагая данными по размерам популяций и не имея примерной схемы лесосеменных районов, которая появилась на 10 лет позже, а также учитывая огромные ареалы ви-дов-эдификаторов на территории страны, в основу координатной сетки пунктов испытаний был положен слишком крупный «шаг», намного превышающий размер популяции. Судя по данным С.А. Мамаева, Л.Ф. Семерикова, В.П. Путенихина и А.И. Видякина, размер популяции тождественен урочищу или лесному массиву. В лесостепной зоне это приблизительно соответствует площади одного лесничества в 100–200 тыс. га.
Последнее, на первый взгляд, противоречит данным по изучению географических культур. Многие исследователи приходят к выводу о возможности переброски семян на большие расстояния. Например, один из основателей отечественного лесного семеноводства М.М. Ве-ресин, отталкиваясь от имеющихся данных по дифференциации климатипов в географических культурах, полагал, что для равнинных территорий переброска семян с севера на юг возможна на 300–600 км, а с востока на запад – на 500–850 км. Ещё более масштабную переброску рекомендуют и некоторые современные исследователи географических культур. Например, при дефиците семян для целей лесовосстановления нижнеангарских сосняков допускается использовать семена из южной части Карелии, т.е. на расстоянии свыше 4 тыс. км [82]. Вероятно, именно такие «смелые» заключения и были приняты во внимание при укрупнении лесосеменных районов в новом варианте районирования. В данном случае авторы путают задачу выделения сортов-популяций, которые могут быть районированы на больших территориях при условии недопущения их скрещивания с местными популяциями, с задачей разработки лесосеменного районирования, преследующего не только унитарные цели, но и сохранение естественно-исторически сложившейся популяционной структуры видов. Необходимо напомнить, что фенотипически сходные особи могут иметь различные генотипы, тем более по экологически лабильным полигенно обусловленным количественным признакам. В этой связи стоит упомянуть и о морфологически идентичных видах-двойниках, потерявших способность к скрещиванию. Реализация такого рода рекомендаций приведёт к генетическому загрязнению и дестабилизации сложившейся пространственной популяционной структуры видов. Критические отзывы специалистов на новый проект лесосеменного районирования уже появились в открытой печати [83]. С учётом современных знаний становится очевидным, что результаты географических испытаний по крупной координатной сетке пунктов заготовки семян, превышающей масштаб карты популяционной структуры, могут лишь косвенно приниматься во внимание при уточнении схемы лесосеменного районирования [36]. Такие результаты с точки зрения обсуждаемой задачи выявляют лишь географические тренды в наследственной изменчивости признаков.
Вместе с тем в тех случаях, когда оценивают норму реакции климатипов по данным их испытаний на различных экофонах (в различных пунктах), дифференциация климатипов по особенностям их реакции (взаимодействиям «генотип–среда») становится более очевидной. Вероятно, оценка взаимодействий «климатип – среда» возможна и по данным одного пункта испытаний при изучении климатического отклика деревьев по ширине и структуре годичных колец. Напомним также, что выводы о дифференциации климатипов по данным географических культур могут существенно измениться после проведения рубок ухода, устраняющих маскирующие эффекты различий в степени загущенности древостоев. Несомненно, что для уточнения популяционной структуры лесообразующих видов большое значение может иметь применение адекватных фенетических и ДНК-маркеров [22, 23, 68].
Завершая краткий анализ проблемы, выразим уверенность, что после её детального обсуждения российским научным сообществом лесосеменное районирование лесообразующих пород будет усовершенствовано и завершится публикацией столь же основательной для современного этапа развития науки коллективной монографией, какой была и остаётся «Лесосеменное районирование основных лесообразующих пород в СССР» [36].
Достижения и проблемы применения современных биотехнологий в лесной селекции
Выше отмечалась несомненная перспективность молекулярной генетики и клонального микроразмножения для решения таких задач, как генетическая паспортизация деревьев и популяций, идентификация патогенов, размножение ценных генотипов лиственных пород. Актуальность разработки и применения в Российской Федерации новейших технологий в селекции деревьев отражена в одном из направлений лесной биотехнологии Госпрограммы «БИО2020» (2012) – «Создание биотехнологических форм деревьев с заданными признаками» (утверждена Правительством РФ 24.04.2012 № 1853п-П8). Не будет преувеличением сказать, что в настоящее время большая часть бюджетных средств и средств грантодателей в области биологии расходуется именно на эти направления. Существенные и стабильные вложения в эту сферу деятельности в последние годы позволили достичь определенных результатов.
Кратко опишем перспективы использования методов биотехнологии в лесной селекции и лесном семеноводстве.
Методы клонального микроразмножения растений (включая соматический эмбриогенез) предназначены для ускорения репродукции ценных генотипов, в том числе для производства высококачественного посадочного материала при создании лесосырьевых плантаций. Работы по культуре in vitro лиственных и хвойных пород родов Populus , Betula , Salex , Fraxinus , Pinus и Larix ведутся в России достаточно давно. Особенно актуальны они для плохо черенкующихся хозяйственно ценных пород и гибридов, в том числе лиственных – тополей белого и сереющего, осины. Принимая во внимание результаты многолетних исследований российских ученых по культуре in vitro осины, гибридных тополей и ив во ВНИИЛГИСбиотех, различных видов березы в Институте леса КарНЦ РАН, триплоидной осины и различных клонов березы в СПбНИИЛХ, ясеня и ив в ИБХ РАН, тополя серебристого в Центральном сибирском ботаническом саду СО РАН, можно прогнозировать успешное использование этих пород для плантационного лесовыращивания [84]. Определённые успехи в этом направлении достигнуты также в ИЛ СО РАН по лиственнице [85].
Одна из первых в России коллекций in vitro лиственных древесных растений была сформирована во ВНИИЛГИСбиотех в 1991 г. Эта коллекция содержит продуктивные диплоидные и триплоидные гибриды тополя белого (Populus alba L.) и тополя сереющего (P. canescens Sm.), продуктивные и гнилеустойчивые формы осины (P. tremula L.), формы березы повислой (Betula pendila L.) и березы пушистой (B. pubescens Ehrh.), генотипы березы карельской (B. pendula Roth vаr. саrеliса Мerkl.) с узорчатой текстурой древесины, декоративные формы березы далекарлийской (B. pendula «dalekarlica» (L. f.)), а также быстрорастущие формы различных видов ивы (Salix spp.) [86]. Хранение коллекции осуществляется двумя способами: 1) длительными (до полугода) субкультивациями на безгормональной среде, 2) депонированием при пониженных температурах, что удлиняет интервал между пересадками до года. Исследование влияния условий долгосрочного хранения in vitro (не менее 27 лет) на последующее поведение растений B. pubescens и B. pendula var. сarelica показало, что оба режима обеспечивают высокие показатели мультипликации и ризогенеза, а также отсутствие признаков сомаклональной изменчивости у этих видов [87].
Полевые испытания после длительного (до 25 лет) хранения in vitro показали как высокую приживаемость и сохранность растений, так и их идентичность исходным генотипам. Например, генотипы тополя белого после 20-летнего хранения в коллекции in vitro сохраняли высокую жизнеспособность и регенерационную активность, а полевые испытания в течение 17 лет после 2–3 лет культивирования in vitro продемонстрировали высокую приживаемость, сохранность характерных признаков и, что очень важно, отсутствие сомаклональной изменчивости [88]. Растения двух генотипов карельской березы, высаженных в питомник после хранения в культуре in vitro в течение 1, 5 или 11 лет, также не показали генетических изменений [27]. Таким образом, данная коллекция ценных генотипов может использоваться не только для сохранения ex situ , но и для создания лесных плантаций.
Коллекция асептических культур in vitro ценных генотипов осины, березы, ивы также была создана в филиале Института биоорганической химии РАН (ФИБХ, Пущино). Эта коллекция использовалась при широкомасштабном получении посадочного материала (более 1 млн растений) для закладки промышленных плантаций в различных субъектах Российской Федерации. По результатам исследований был получен ряд патентов на способы хранения и криоконсервации растительного материала in vitro .
Ещё одно перспективное направление исследований с применением современных методов культуры клеток и тканей – создание биотехнологических тест-систем in vitro для отбора на устойчивость. Исследователи из ВНИИЛГИСбио-тех предложили проводить отбор устойчивых генотипов на средах с добавлением NaCl, что позволяет моделировать солевой и осмотический стресс. Для лиственных пород (береза, тополь, осина) оценку проводили на стеблевых сегментах. В результате удалось выделить генотипы, устойчивые к летальному засолению. Для индикации стрессоустойчивости сосны обыкновенной применяли каллусную культуру. Разработанная биотест-система на основе каллусогенеза может быть использована в комплексной оценке и отборе устойчивых к стрессам генотипов взрослых деревьев. В Институте леса СО РАН каллусной культурой сосны обыкновенной (Pinus sylvestris L.) воспользовались для изучения влияния различных веществ и условий культивирования на лигнификацию.
В процессе культивирования in vitro растительной ткани (особенно каллусной) возникает сомаклональная изменчивость, которая нежелательна при клональном микроразмножении и сохранении in vitro ценных генотипов. Нестабильность геномов является серьёзной помехой при сохранении и репродукции ценных генотипов, а также в специальных исследованиях по секвенированию геномов. Однако это явление может быть использовано для получения новых растений и клеточной селекции. Например, путем регенерации через каллус был получен клон березы карельской с комплексом измененных признаков (отсутствие ризогенеза, кустообразный габитус), сохраняющихся на протяжении 18 лет культивирования in vitro [87].
В последние годы в России расширяется применение маркер-опосредованной селекции ( marker assisted selection , MAS ) для отбора генотипов с ценными признаками. MAS относится к непрямой селекции и заключается в статистическом связывании молекулярных маркеров с хозяйственно важными признаками. В силу того что на маркеры не оказывают влияние условия среды или особенности онтогенеза, данный инструмент предоставляет широкие возможности, особенно при ранней диагностике (например, качество древесины на стадии сеянцев). Первым этапом в такой работе является анализ локусов количественных признаков (QTL, Quantitive trait loci), который проводят путем их картирования в гибридной популяции.
В Санкт-Петербургском государственном лесотехническом университете им. С.М. Кирова в 2016 г. провели скрещивание двух генотипов осины с различным фенотипом и полученную популяцию высадили в трех регионах с разными климатическими условиями: Ленинградская обл., Ханты-Мансийский автономный округ — Югра (Сургут) и Воронежская обл. С помощью технологии RADseq в родительских деревьях и Ленинградской популяции идентифицировали однонуклеотидные замены (SNP), на основе которых составили генетические карты высокой плотности [88]. Полученные данные планируется использовать для идентификации QTL, связанных с такими свойствами, как быстрый рост, устойчивость к сердцевинной гнили, качество древесины и др., а также проведения маркер-опосредованной селекции осины. По результатам QTL-анализа Ленинградской популяции был выявлен локус, сопряженный с высотой однолетних саженцев. Генотипирование Воронежской и Сургутской популяций показало, что этот локус сохраняет своё влияние на показатели роста осин в разных климатических условиях [89]. Таким образом удалось выявить стабильный, не зависящий от климатических условий локус, связанный с ростом осины в молодом возрасте.
Несмотря на всю привлекательность метода для практической селекции, существует ряд ограничений, которые будут препятствовать его применению, по крайней мере в среднесрочной перспективе:
-
V отбор на основе маркеров все еще достаточно дорог для какой-либо крупной группы особей;
-
V связь между маркерами и хозяйственно ценными признаками должна устанавливаться для каждой семьи отдельно;
-
V в различных экологических условиях один и тот же признак может быть сцеплен с различными маркерами.
Последнее повторяет феномен лабильности связей качественных генетически обусловленных признаков с количественными показателями, по которым ведётся отбор, и свидетельствует о большей эффективности прямого отбора по сравнению с любым косвенным [2].
Технологией будущего десятилетия называют трансгенез (геномодификация) древесных растений. Однако в законодательстве Российской Федерации и многих стран применять геномодифицированные растения можно лишь в случае доказательства безопасности их воздействия на окружающую среду. Для лесных деревьев подобная задача не может быть решена на основе краткосрочных опытов, так как деревья растут десятки (и даже сотни) лет, поэтому данные исследования в настоящей статье не рассматриваются. Отметим только, что они проводятся в России на некоторых лиственных породах: продемонстрирована успешность применения трансгенеза для быстрого получения растений с новыми свойствами, в том числе по содержанию лигнина в древесине, эффективности усвоения азота, устойчивости к гербицидам и другим признакам.
Еще одним перспективным направлением биотехнологии растений является использование клеточных культур для синтеза биологически активных веществ. Среди лесных древесных пород большой интерес представляют растения разных видов тиса ( Taxus spp.), которые содержат дитерпеноиды таксанового ряда (таксоиды), обладающие противоопухолевой активностью. В Институте физиологии растений РАН (Москва) ведутся работы со суспензионными культурами Taxus baccata , T. media , T. wallichiana и T. canadensis [90].
В последние десятилетия бурно развивается новое научное направление на стыке генетики и биоинформатики – геномика. В 2014 г. в Сибирском федеральном университете под руководством проф. К.В. Крутовского стартовал большой проект по секвенированию геномов основных лесообразующих пород бореальных лесов Евразии – лиственницы сибирской, сосны кедровой сибирской и сосны обыкновенной, а также их наиболее опасных патогенов, которые вызывают катастрофическое усыхание российских бореальных лесов. Проект «Геномные исследования основных бореальных лесообразующих хвойных видов и их наиболее опасных патогенов в Российской Федерации» финансируется Правительством РФ13. Лиственница сибирская и сосна кедровая сибирская как важнейшие виды бореальных лесов Сибири имеют огромное экономическое, экологическое и эстетическое значение. Данные об их геноме и генах, контролирующих важные адаптивные и селекционные признаки, на момент начала исследований практически отсутствовали.
Первые результаты работ по проекту для лиственницы сибирской и сосны сибирской были опубликованы, когда удалось получить черновые сборки транскриптомов, а также хлоропластных, митохондриальных и ядерных геномов и появилась возможность разработки молекулярно-генетических маркеров, в том числе микросателлит-ных локусов [91, 92]. Работы по секвенированию проводились не только для ядерного, но и для митохондриального и хлоропластного геномов вышеперечисленных видов. Российским исследователям удалось добиться качественной сборки митогенома сибирской лиственницы, который в настоящее время является самым крупным [93]. Впервые была получена полная последовательность хлоропластного генома лиственницы сибирской, разработан ряд высокополиморфных молекулярно-генетических маркеров.
Работы по проекту полногеномного секвенирования хвойных видов России в Сибирском федеральном университете позволили разработать видоспецифичные микросателлитные праймеры для лиственницы сибирской. Из полученной сборки генома лиственницы было протестировано 59 пар праймеров для локусов с динуклеотид-ными мотивами, которые обладают наибольшей изменчивостью. Из них, в итоге, были отобраны 11 локусов с высоким уровнем полиморфизма, которые могут использоваться как для идентификации происхождения древесины и растительного материала, так и в популяционно-генетических исследованиях. Позднее в геномной de novo сборке лиственницы сибирской был проведен дополнительный поиск более удобных для генотипирования микросателлитных локусов с три-, тетра-, пента- и гексануклеотидными тандемными повторами, в результате чего были разработаны и проверены 60 пар праймеров [94]. По итогам тестирования было отобрано 14 наиболее перспективных полиморфных локусов, которые могут успешно применяться для изучения и идентификации не только лиственницы сибирской, но и двух других видов лиственниц – L. gmelinii (Rupr.) Rupr. и L. cajanderi Mayr. До недавнего времени микросателлитные маркеры также отсутствовали для сосны сибирской, но на основе черновых сборок генома этого вида было отобрано 18 видоспецифичных полиморфных локусов, которые могут использоваться в качестве молекулярно-генетических маркеров [92]. Данные маркеры можно эффективно применять для идентификации географического происхождения древесины и растительного материала в борьбе с нелегальными рубками и незаконным оборотом лесного посадочного материала. Исследования, проведённые в рамках данного проекта, стали основой разработки высокоинформативных молекулярно-генетических маркеров для селекции, а также изучения механизмов патогенности ряда грибов и отбора древесных растений на устойчивость [95].
В настоящее время в России для хвойных пород не реализована теоретически возможная перспектива ускорения селекции с применением метода «marker-assisted selection (MAS)» и «gene-assisted selection (GAS)» (маркер- и ген-ассоци-ированного отбора), с помощью которых предполагалось выявить Qtl-гены, ассоциированные с целевыми количественными признаками, и перспективные генотипы. Во-первых, для этого необходимо получить потомков второго поколения гибридов ( F 2) от контролируемых скрещиваний, что для хвойных пород требует много времени и значительных средств. Во-вторых, обнаружение маркеров (особенно в большом количестве), обычно слабокоррелированных с количественными признаками, не так много добавляет к традиционным методам количественной генетики и генетического/гибридологического анализа.
Например, если предположить, что выявлено 8 генов (участков ДНК), ассоциированных с устойчивостью к фитопатогенам, то возникает задача комбинирования нужных локусов этих генов в одном генотипе. Только в очень редкой ситуации, когда они случайным образом окажутся локализованными в нужном сочетании хотя бы в 2-х генотипах, их выявление и скрещивание сразу приведут к получению нужной линии (сорта). Для выявления таких генотипов понадобится огромное число тестов. Чаще будет возникать ситуация, когда необходима комбинация скрещиваний с последующим скринингом и отбором нужных генотипов в нескольких поколениях. Однако выявленные в затратных экспериментах гены и аллели могут оказаться не самыми важными, и их сочетание в одном генотипе может дать в итоге лишь небольшой эффект от потенциально возможного. Судя по всему, эти задачи достаточно сложные и пока никем не реализованы, во всяком случае в России и странах СНГ не появилось ни одного сорта древесных растений, полученных с использованием технологии MAS.
Кроме рассмотренных, в лесной селекции существуют проблемы «переопределения генетической формулы признаков» и «эмержентно-сти» количественных признаков по отношению к генам [29, 96, 97]. Даже если удастся собрать в одном генотипе все ассоциированные с нужными признаками аллели нужных генов, это не будет гарантировать надежного генетического сдвига по селектируемому признаку в варьирующей среде обитания по следующим причинам. Во-первых, при изменении экологических условий созданное сочетание аллелей перестанет обеспечивать заданный генетический сдвиг (взаимодействие «генотип – среда»). Во-вторых, в силу эмержентности на уровне целостных генотипов должны существовать наследственные факторы «надмолекулярного» уровня, не сводимые к совокупности тех или иных локусов тех или иных генов. Как справедливо отмечает В.А. Дра-гавцев, не существует, например, «генов высоты ствола». Белковые продукты генов оказывают лишь косвенное воздействие на формирование этого сложного признака, который реализуется на уровне организма как целого в его непростом взаимодействии с окружающей средой. Только при выявлении небольшого числа генов (вряд ли больше 3–4), существенно коррелированных с изменчивостью селектируемых признаков в известном градиенте лимитирующих факторов, характерном для возделывания будущих сортов, методы молекулярной генетики смогут помочь практикующим лесным селекционерам существенно ускорить процесс выведения сортов. Именно на это, с нашей точки зрения, должны быть нацелены усилия молекулярных генетиков для эффективного приложения их результатов в сфере лесной селекции.
В совокупности рассмотренные проблемы подчеркивают важность, во-первых, понимания организма и генотипа как целого [98, 99], во-вторых, оценки наследуемости количественных признаков на так называемом «феноменологическом» уровне – на основе корреляций между родственниками той или иной степени родства [100] и, в-третьих, необходимость всесторонней оценки особенностей нормы реакции генотипов в контролируемых условиях, без которой вряд ли удастся научиться управлять селекционным процессом [29].
В последней связи нельзя не упомянуть о развитии в России методов идентификации генотипов по фенотипам без смены поколений и о Теории эколого-генетической организации количественных признаков (ТЭГОКП) [96, 97, 101–103]. Эта теория разработана в целом для растений, но ряд работ её идеолога В.А. Драгавцева посвящен непосредственно древесным растениям. Идентификация ценных генотипов, устойчивых к лимитирующим экологическим факторам, у древесных пород может осуществляться, во-первых, сопоставлением динамики годичных приростов древесины с динамикой метеорологических факторов. Во-вторых, к древесным растениям может применяться метод «фоновых признаков». Отметим, что на первых этапах разработки этого метода к экологически лабильным фоновым признакам предъявлялось почти не выполнимое требование отсутствия генетической дисперсии, что сдерживало практическое применение метода.
Но в доработанном варианте метода допускается, что фоновый признак может иметь и собственную генетическую дисперсию.
Что касается ТЭГОКП, то в её основе лежит постулат о переопределении «генетической формулы» количественных признаков при смене лимитирующих экологических факторов, что и приводит к эффекту взаимодействия «генотип – среда». В этой связи важнейшим инструментом отбора растений является оценка его нормы реакции на главные лимитирующие факторы. Автор данной теории считает, что главным инструментом селекционеров должен стать фитотрон, позволяющий имитировать любые сочетания лимитирующих факторов для оценки нормы реакции [97]. «Просеивая» большое число растений на различных сочетаниях провокационных фонов, можно быстро и надежно выделить редкие генотипы с нужными параметрами фенотипических свойств в предполагаемом районе возделывания. Применительно к древесным растениям с длительным онтогенезом это позволит отбирать генотипы, способные на ранних стадиях развития «включать» соответствующие среде эпигены («каналы развития» по Уоддингтону14), способствующие максимальной адаптации к данному сочетанию условий. Судя по имеющимся данным, вполне возможно, что фитотрон может выступать в качестве инструмента предварительной «эпигенетической настройки» сорта на определенные условия возделывания. Как отметили Л.И. Милютин с соавт. [102], «Теория эколого-генетической организации количественных признаков растений… позволила прогнозировать трансгрессии.., гетерозис, составлять прогнозы величин взаимодействия «генотип – среда», подбирать оптимальные пары родителей для скрещивания и делать другие прогнозы, …это привело … к созданию новой технологии селекции растений на продуктивность и урожайность».
К сожалению, практическое применение этой теории сдерживается отсутствием средств на создание селекционных фитотронов для древесных растений. В свете теории ТЭГОКП очевидна необходимость модификации методики испытательных культур лесообразующих пород, которые целесообразно создавать непосредственно в месте возделывания будущего сорта на различных агрофонах, отличающихся по уровням стимуляции плодородия почв (слабый, средний, сильный). У перспективных для выделения в качестве сорта-популяции климатипов, испытываемых одновременно в различных пунктах (типах лесорастительных условий), должна быть оценена норма реакции на комплекс почвенно-климатических факторов. На всех селекционных объектах ценная информация о норме реакции может быть получена при ретроспективном анализе динамики приростов по высоте и диаметру в связи с динамикой метеоусловий.
Подводя итоги, отметим, что, несмотря на достигнутые успехи, в практике лесного селекционного семеноводства клонирование наиболее ценных хвойных пород до сих пор преимущественно осуществляется методом прививки. Этот технически простой и недорогой метод вполне приемлем для целей сохранения генофонда и создания ЛСП, но не решает проблемы массового выращивания клонированных деревьев на промышленных лесосырьевых плантациях, в связи со снижением качества древесины у привитых деревьев. Некоторым исключением могут оказаться лишь клоны межвидовых гибридов лиственницы [15].
Не отрицая необходимости дальнейшего развития и несомненной перспективности методов молекулярной генетики и клонального микроразмножения в лесной селекции, нельзя не отметить, что развитие традиционных/классиче-ских методов на данный момент демонстрирует их более высокую практическую эффективность. Даже в условиях очень низкого финансирования этого направления в последние десятилетия были созданы ЛСПпгц и выведены первые сорта лесных деревьев, а также созданы ЛСП-1, дающие урожаи семян на генетической основе местных популяций (поскольку соблюдение лесосеменного районирования при создании лесосеменных объектов четко контролировалось) [37]. Из этого следует экономическая целесообразность существенного увеличения финансирования этой эффективной наукоемкой отрасли, включая вложения в развитие новых методов, основанных на углубленной оценке нормы реакции генотипов, в том числе в селекционных фитотронах.
Основные проблемы и приоритетные направления развития лесной селекции в России
Для повышения эффективности лесной селекции в России необходимо решить ряд проблем: 1) концептуальную; 2) экономическую; 3) организационную; 4) научную. Первая (концептуальная) проблема заключается в убежденности части лесоводов и лесных чиновников в том, что задачу сохранения генетической гетерогенности и устойчивости «селекционно улучшенных» лесов можно решить такими полумерами, как увеличение числа клонов на ЛСП, а также уверенность в том, что преобразование естественных лесов России в облагороженные селекцией хозяйственно трансформированные леса (как, например, в Финляндии) является высочайшим благом для нашей страны. В действительности, высочайшим благом для России, навсегда утерянным для технологически развитых стран, временно обогнавшим нас в области лесной биотехнологии, является именно обилие естественных лесов, основу которых составляют высокополиморфные «дикие» популяции основных лесообразующих видов.
Сказанное вовсе не означает, что сохранение генетической гетерогенности и устойчивости естественных лесов и развитие лесной селекции несовместимы. Лесная селекция – очень перспективная, эффективная наукоёмкая отрасль, без развития которой не решить задачу интенсификации лесного хозяйства России. Это возможно только при условии, что она будет нацелена на выведение сортов, предназначенных для создания быстрорастущих промышленных плантаций с ускоренным (сокращенным по времени)
производственным циклом их эксплуатации. Практическая возможность и целесообразность создания таких плантаций давно продемонстрирована не только за рубежом, но и в странах СНГ, включая Россию. При этом важно, чтобы лесные плантации создавали на относительно небольшой части лучших по лесорастительным условиям площадей с применением стимулирующих агротехнических приёмов. Создание подобных сортовых насаждений должно осуществляться таким образом, чтобы не приводить к деградации естественно-исторически сложившейся популяционной структуры лесообразующих видов и снижению их генетического потенциала (генетической эрозии). Для этого быстрорастущие промышленные плантации необходимо создавать на участках, тем или иным образом изолированных от естественных популяций того же вида, и воспроизводить исключительно искусственным путём, как это делается в сельском хозяйстве. Принятие перечисленных выше идей:
-
1) устраняет не имеющую приемлемого решения проблему сохранения высокого уровня генетической изменчивости при отборе;
-
2) в значительной мере решает проблему ранней диагностики, так как отбору подлежат деревья, дающие селекционный эффект в возможно более раннем возрасте;
-
3) существенно ослабляет эксплуатационную нагрузку на естественные леса, выполняющие экологические (биосферные) функции.
Вторая (экономическая) проблема – резко снизившийся уровень финансирования лесного семеноводства – тесно связана с первой. Это обусловлено весьма отдаленной окупаемостью затрат. Только резкое сокращение сроков выведения сортов и нацеленность на плантации с ускоренным производственным циклом лесо-выращивания способны переломить негативную ситуацию низкого финансирования лесного семеноводства.
Третья (организационная) проблема многоплановая. Укажем лишь, во-первых, на губительность непрерывных реорганизаций системы лесного хозяйства. Она особенно неблагоприятна для наукоёмких отраслей, к которым относится лесная селекция, тем более в ситуации, когда различные звенья научно-производственной цепочки должны быть надёжно связаны друг с другом. Реорганизация в этом случае приведет к потере объектов ЕГСК, квалифицированных кадров, снижению эффективности взаимодействия науки и производства и утраты преемственности, без которых лесная селекция просто невозможна. Во-вторых, с учетом современных реалий для повышения эффективности селекции целесообразно разделить функции научных организаций и лесных предприятий по принципу: «наука выводит сорта, а лесные предприятия используют их для производства селекционно улучшенных семян и посадочного материала». Для реализации этого принципа научные организации должны иметь на правах бессрочного пользования для научных целей соответствующие лесные площади и объекты (испытательные культуры и архивы клонов). В любом случае необходимо восстановить систему научного кураторства объектов ЕГСК, поскольку много нерешенных задач есть и в чисто семеноводческой сфере.
Четвертая проблема связана с необходимостью существенной модернизации устаревших научных программ и методов лесной селекции с учётом накопленного опыта и последних достижений лесной генетики, селекции, семеноводства и интродукции древесных растений. В этой связи приоритетными являются:
-
1) разработка национальной и региональных программ по сохранению и рациональному использованию лесных генетических ресурсов. Прообразом национальной программы может служить программа, разработанная в цикле научных совещаний по этой теме15 [31];
-
2) разработка и внедрение в практику новых методик по отбору и генетической оценке плюсовых деревьев, нацеленных на главную задачу – выведение быстрорастущих сортов, предназначенных для создания промышленных
плантаций с ускоренной ротацией на высоком агрофоне;
-
3) переход селекционеров от малоинформативного этапа изучения полусибовых потомств, получаемых от свободного опыления плюсовых деревьев в естественных древостоях, к этапу анализа их потомств, получаемых от контролируемых скрещиваний клонов на плантациях (поликросс, топкросс, диаллельные и др. скрещивания);
-
3) коренная переработка лесосеменного районирования на основе критического анализа всей совокупности имеющихся данных о популяционно-генетической структуре лесообразующих видов и результатов географических опытов, что возможно при условии привлечения к этой работе ведущих специалистов и открытого обсуждения полученных результатов научным сообществом, в том числе и прежде всего анализа последствий отсутствия своевременных рубок ухода на объектах ЕГСК и особенностей нормы реакции климатипов;
-
4) стимуляция исследований по разработке методов клонального микроразмножения и сохранения стабильности геномов ценных генотипов хвойных лесообразующих пород с целью ускоренного выведения и репродукции сортов-клонов для их использования на промышленных плантациях; при селекции на быстроту роста и качество древесины крайне важно сохранить качество древесины клонируемых деревьев;
-
5) организация молекулярно-генетических исследований по выявлению и локализации ограниченного числа наиболее важных генов («олигогенов»), сильно коррелированных с количественными целевыми признаками в выровненных почвенно-климатических условиях промышленных плантаций конкретных регионов;
-
6) разработка отечественных моделей и производство селекционных фитотронов для оценки нормы реакции и ускорения отбора на ранних стадиях онтогенеза ценных генотипов, проявляющих требуемые свойства при специфическом
сочетании лимитирующих экологических факторов промышленных плантаций; создание испытательных культур в месте возделывания будущих сортов на различных агрофонах; оценка нормы реакции генотипов и популяций при анализе динамики приростов по диаметру и высоте в связи с динамикой метеорологических факторов;
-
7) дальнейшее развитие и внедрение в практику лесного хозяйства селекционных методов уровня «природоподобных технологий», предусмотренных Стратегией научно-технологического развития Российской Федерации, которые сохраняют высокий уровень генетической гетерогенности насаждений, свойственный природным популяциям, и перспективны для следующих ситуаций: в среднепродуктивных лесорастительных условиях (ниже I–II классов бонитета), при восстановлении генетического потенциала насаждений, а также при лесоразведении, в том числе в экстремальных экологических условиях. К таким методам относятся: а)
популяционный (групповой) отбор; б) методы «адаптивной селекции», основанные на отборе высокоустойчивых к лимитирующим факторам генотипов и популяций; в) технология «плантационно-обсеменительных» культур, создаваемых редкой посадкой клонов лучших по семеноше-нию и росту деревьев из местных насаждений для последующего естественного обсеменения ими территории.
Проблематика лесной генетики и селекции (сохранения, развития и устойчивого использования лесных генетических ресурсов) пока известна небольшому числу ученых и специалистов лесного хозяйства России. Вместе с тем для достижения более значимых результатов она должна найти отражение в государственной лесной политике Российской Федерации, специальных лесохозяйственных региональных и федеральных программах. Очень важно привлечь общественный интерес к задачам сохранения и устойчивого использования лесных генетических ресурсов.
Список литературы Лесная селекция в России: достижения, проблемы, приоритеты (обзор)
- О генетико-селекционном аспекте сохранения и улучшения лесов России / Л.Ф. Семериков, Ю.Н. Исаков, B.В. Тараканов [и др.] // Лесохозяйственная информация. - 1998. - № 9-10. - С. 3-12 ; 29-40.
- Селекционное семеноводство сосны обыкновенной в Сибири / В.В. Тараканов, В.П. Демиденко, Я.Н. Ишу-тин [и др.]. - Новосибирск : Наука, 2001. - 230 с.
- Указания по лесному семеноводству в Российской Федерации. - М. : ВНИИЦлесресурс, 2000. - 198 с.
- Рогозин, М.В. Программа селекции хвойных в лесосеменном районе / М.В. Рогозин // Сибирский лесной журнал. - 2016. - № 5. - С. 99-106.
- Царёв, А.П. Селекция лесных и декоративных древесных растений: учеб. / А.П. Царёв, С.П. Погиба, Н.В. Лаур. - М. : МГУЛ, 2014. - 552 с.
- Опыт создания лесосеменных плантаций в СССР / Ю.П. Ефимов, Ю.А. Данусявичус, В.И. Долголиков [и др.] // Лесная генетика, селекция и физиология древесных растений : матер. междунар. симп. (25-30 сентября 1989 г., Воронеж). - М., 1989. - С. 85-91.
- Ефимов, Ю.П. Семенные плантации в селекции и семеноводстве сосны обыкновенной / Ю.П. Ефимов // Воронеж : изд-во Исток, 2010. - 252 с.
- Исаков, Ю.Н. Эколого-генетическая изменчивость и селекция сосны обыкновенной : автореф. ... д-ра биол. наук / Ю.Н. Исаков. - СПб, 1999. - 36 с.
- Царёв, А.П. Государственная регистрация лесных селекционных достижений в России / А.П. Царёв // Лесной вестник. - 2016. - № 5. - С. 184-190.
- Царёв, А.П. Селекция и сортоиспытание тополей / А.П. Царёв, Ю.В. Плугатарь, Р.П. Царёва. - Симферополь : ИТ «Ариал», 2019. - 252 с.
- Бакулин, В.Т. Интродукция и селекция тополя в Сибири / В.Т. Бакулин. - Новосибирск : Наука, 1990. - 174 с.
- Цитологические, молекулярно-генетические и лесоводственно-селекционные исследования полиплоидных тополей / А.И. Сиволапов, Д.В. Политов, О.С. Машкина [и др.] // Сибирский лесной журнал. - 2014. - № 4. - C. 50-58.
- Климов, А.В. Фенотипическое разнообразие качественных признаков Populus nigra L., P. laurifolia Ledeb. и P. x jrtyschensis Ch.Y. Yang. в зоне естественной гибридизации / А.В. Климов, Б.В. Прошкин // Вавиловский журнал генетики и селекции. - 2018. - Т. 22. - № 4. - С. 468-475.
- Путенихин, В.П. Популяционная структура и сохранение генофонда хвойных видов на Урале : автореф. дис. ... д-ра биол.наук : 06.03.01 / В.П. Путенихин. - Красноярск, 2000. - 49 с.
- Ирошников, А.И. Лиственницы России. Биоразнообразие и селекция. - Т. 1. Состояние и перспективы / А.И. Ирошников. - М. : ВНИИЛМ, 2004. - 182 с.
- Пути генетического улучшения лесных древесных растений. - М. : Наука, 1985. - 240 с.
- Коропачинский, И.Ю. Естественная гибридизация древесных растений / И.Ю. Коропачинский, Л.И. Милютин. - Новосибирск : Гео, 2006. - 223 с.
- Горошкевич, С.Н. Генофонд кедра сибирского: сохранение, исследование, резервация и использование для селекции сортов // Опыт создания и проблемы развития единого генетико-селекционного комплекса (ЕГСК) в Сибири. - Новосибирск : «Российский центр защиты леса» ЦЗЛ Новосибирской области, 2008. - С. 40-56.
- Молотков, П.И. Стратегия селекции и семеноводства сосны обыкновенной / П.И. Молотков, И.Н. Патлай // Лесная генетика, селекция и физиология древесных растений. (доп. вып.). - М., 1990. - С. 9-16.
- Струнников, В.А. Природа гетерозиса, методы его повышения и закрепления в последующих поколениях без гибридизации / В.А. Струнников, Л.В. Струнникова // Известия АН. Сер. биологическая. - 2000. - № 6. - С. 679-687.
- Тараканов, В.В. Перспективы селекции Pinus sylvestris L. на гетерозис / В.В. Тараканов // Ш съезд ВОГиС (6-12 июня 2004 г.). - Т. 1. - М., 2004. - 283 с.
- Видякин, А.И. Методические основы выделения фенов лесных древесных растений (на примере сосны обыкновенной Pinus sylvestris L.) / А.И. Видякин // Новые научные методики и информационные технологии. -Вып. 65. - Сыктывкар, 2010. - 28 с.
- Тараканов, В.В. Фенетический анализ клоновых и естественных популяций сосны в Алтайском крае / B.В. Тараканов, Л.И. Кальченко. - Новосибирск : изд-во «Гео», 2015. - 107 с.
- Климов, А.В. Использование морфо-анатомических признаков для выявления гибридных растений в зоне естественной гибридизации Populus laurifolia и P. nigra в Сибири, Россия / А.В. Климов, Б.В. Прошкин // Вестник Томского государственного университета. Сер. Биология. - 2019. - № 46. - С. 64-81.
- Опыт восстановления утерянных селекционных достижений Populus leningradensis Bogd. и Populus newensis Bogd. на основе микросателлитного анализа / М.В. Лебедева, Э.А. Левкоев, В.А. Волков [и др.] // Генетика. - 2016. - Т. 10. - С. 1159-1168.
- Fast and cheap identification of elite aspen clones in the North-West of Russia using ISSR markers / A.V. Zhigunov, D.A. Shabunin, O.Y. Butenko, M.V. Lebedeva // Folia Forestalia Polonica. - 2018. - Vol. 60. - № 4. - P. 207-213.
- Использование RAPD- и SSR-маркеров для генетической паспортизации биотипов тополя / С.Г. Ржевский, Т.А. Гродецкая, Т.П. Федулова, В.А. Царев // Труды СПбНИИЛХ. - 2019. - № 3. - С. 51-61.
- Маслов, А.А. Идентификация видов берез в заболоченных лесах Центра Русской равнины по результатам молекулярно-генетического анализа / А.А. Маслов, О.Ю. Баранов, А.А. Сирин // Лесоведение. - 2019. - № 3. - С. 177-187.
- Драгавцев, В.А. Решения технологических задач селекционного повышения урожаев, вытекающие из теории эколого-генетической организации количественных признаков / В.А. Драгавцев // АгроФорум. - 2019. -№ 1. - С. 64-71.
- Тараканов, В.В. Изучение и развитие научного наследия Н.В. Глотова: норма реакции генотипа и взаимодействие генотип-среда в природной популяции древесных растений / В.В. Тараканов, И.В. Тихонова // Проблемы популяционной биологии : матер. XII Всерос. популяционного семинара памяти Н.В. Глотова (1939-2016). -Йошкар-Ола : ИПФ «СТРИНГ», 2017. - С. 10-13.
- Изучение, сохранение и рациональное использование лесных генетических ресурсов России : проект национальной программы // Сохранение лесных генетических ресурсов Сибири : матер. 4-го междунар. со-вещ. (24-29 августа 2015, Барнаул, Россия). - Красноярск : Институт леса им. В.Н. Сукачева СО РАН, 2015. - C.198-208.
- Рогозин, М.В. Лесная селекция : учебн. пособие / М.В. Рогозин. - М. : ИД Академии Естествознания, 2018. - 298 с.
- Бондаренко, А.С. Оптимизация численности растений в опытах по испытанию семенного потомства плюсовых деревьев ели европейской / А.С. Бондаренко, А.В. Жигунов // Лесоведение. - 2016. - № 3. - С. 187-194.
- Видякин, А. И. Плюсовая селекция сосны и ели: итоги и перспективы развития / А.И. Видякин // Лесохо-зяйственная информация. - 2008. - № 3-4. - С. 33-35.
- Федорков, А.Л. Фенотипический отбор в лесной селекции / А.Л. Федорков // Лесоведение. - 2019. - № 6. -С. 580-584.
- Лесосеменное районирование основных лесообразующих пород в СССР. - М. : Лесн. пром-сть, 1982. -
- Царёв, А.П. Перспективные направления селекции и репродукции лесных древесных растений / А.П. Царёв, Н.В. Лаур // Лесной журнал. - 2013. - № 2 (332). - С. 36-44.
- Состояние и перспективы развития генетико-селекционного комплекса хвойных пород в Сибири (на примере Новосибирской области) / В.В. Тараканов, Д.С. Дубовик, Р.В. Роговцев [и др.] // Вестник Поволжского государственного технологического университета. Сер.: Лес. Экология. Природопользование. - 2019. - № 3(43). -С. 5-24.
- Раевский, Б.В. Селекция и семеноводство сосны обыкновенной (Pinus sylvestris L.) и сосны скрученной (Pinus contorta Dougl. ex Loud. var. latifolia Engelm) на северо-западе таежной зоны России : автореф. дис. ... д-ра с.-х. наук / Б.В. Раевский. - Петрозаводск : СПбГЛА им. С.М. Кирова, 2015. - 43 с.
- Лаур, Н.В. Из истории развития лесной селекции в Республике Карелия / Н.В. Лаур, А.П. Царёв // Наука -лесному хозяйства Севера. - Архангельск : СевНИИЛХ, 2019. - С. 114-119.
- Шутяев, А.М. Биоразнообразие дуба черешчатого и его использование в селекции и лесоразведении / А.М. Шутяев. - Воронеж : НИИЛГиС, 2000. - 335 с.
- Шутяев, А.М. Изменчивость хвойных видов в испытательных культурах Центрального черноземья / А.М. Шутяев. - М., 2007. - 296 с.
- Пастухова, Н.О. Лесоводственно-таксационные признаки смолопродуктивности сосняков и проект мероприятий по повышению выхода живицы / Н.О. Пастухова, А.М. Антонов // Международный студенческий научный вестник. - 2014. - № 3. - С. 31.
- Ильичев, Ю.Н. Состояние клоновых объектов кедра сибирского Pinus sibirica Du Tour Республики Алтай: сохранность и перспективы селекции. / Ю.Н. Ильичев, Д.Н. Шуваев // Сибирский лесной журнал. - 2016. -№ 5. - С. 36-47.
- Селекционно-генетическая оценка клоновых плантаций сосны обыкновенной в Дюртюлинском лесничестве Республики Башкортостан / В.Ф. Коновалов, Ю.А. Янбаев, Т.М. Ганиев [и др.] // Плодоводство, семеноводство, интродукция древесных растений. - 2011. - № XIV. - С. 58-61.
- Бессчетнов, В.П. Селекционная оценка плюсовых деревьев сосны обыкновенной методами многомерного анализа / В.П. Бессчетнов, Н.Н. Бессчетнова // Лесной журнал. - 2012. - № 2. - С. 58-64.
- Рогозин, М.В. Развитие древостоев. Модели, законы, гипотезы / М.В. Рогозин, Г.С. Разин. - Пермь : ПГНИУ, 2015. - 277 с.
- Земляной, А.И. Межклоновая изменчивость кедра сибирского по элементам семенной продуктивности: перспективы отбора / А.И. Земляной, Ю.Н. Ильичев, В.В. Тараканов // Хвойные бореальной зоны. - 2010. -Т. 27. - № 1-2. - С. 77-82.
- Бондаренко, А.С. Оптимизация численности растений в опытах по испытанию семенного потомства плюсовых деревьев ели европейской / А.С. Бондаренко, А.В. Жигунов // Лесоведение. - 2016. - № 3. - С. 187-194.
- Раевский, Б.В. Методика селекционно-генетической оценки клонов сосны обыкновенной на лесосемен-ных плантациях / Б.В. Раевский, М.Л. Щурова // Сибирский лесной журнал. - 2016. - № 5. - С. 91-98.
- Изменчивость, отбор семенного потомства экотипов, плюсовых деревьев и формирование плантационных культур кедровых сосен в пригородной зоне Красноярска / Р.Н. Матвеева, О.Ф. Буторова, Н.П. Братилова, А.М. Пастухова, А.В. Водин. - Красноярск : Сибирский гос. технол. ун-т, 2006. - 268 с.
- Разработка методики идентификации клонов плюсовых деревьев ели обыкновенной (Picea abies L.) с использованием ISSR маркеров / О.В. Шейкина, А.А. Прохорова, П.С. Новиков [и др.] // Научный журнал Куб-ГАУ. - 2012. - № 83. - С. 56-69.
- Шишкина, О.К. Некоторые результаты практического применения анализа ДНК для генетической идентификации клонов на ЛСП сосны обыкновенной / О.К. Шишкина, М.А. Завистяева, А.С. Рабцун // Лесной вестник. - 2013. - № 2(94). - С. 33-34.
- Зацепина, К.Г. Генотипирование деревьев на клоновых плантациях хвойных лесообразующих видов в Западной Сибири / К.Г. Зацепина, А.К. Экарт, В.В. Тараканов // Хвойные бореальной зоны. - 2012. - № 1-2. - С. 67-71.
- Поэтапная паспортизация деревьев на объектах генетико-селекционного комплекса сосны обыкновенной / В.В. Тараканов, Л.И. Кальченко, К.Г. Зацепина [и др.] // Сибирский лесной журнал. - 2014. - № 4. -С. 69-74.
- Криворотова, Т.Н. Фенотипическая и генетическая изменчивость клонов плюсовых деревьев сосны обыкновенной в Среднем Поволжье : дис. ... канд. с.-х. наук : 06.03.01 / Т.Н. Криворотова. - Йошкар-Ола : Поволж. гос. технол. ун-т, 2014. - 141 с.
- Прохорова, Е.В. Оценка роста и семеношения клоновых потомств сосны обыкновенной на коллекционно-маточном участке в республике Марий Эл / Е.В. Прохорова, Э.П. Лебедева, Л.С. Мошкина // Вестник ПГТУ. Серия: Лес. Экология. Природопользование. - 2008. - № 3. - С. 43-49.
- Прохорова, Е.В. Анализ потомства плюсовых деревьев сосны обыкновенной и ели европейской в Среднем Поволжье : автореф. дис. ... канд. с.-х. наук / Е.В. Прохорова. - Йошкар-Ола, 1996. - 24 с.
- Роговцев, Р.В. Продуктивность географических культур сосны в условиях Среднеобского бора/ Р.В. Роговцев, В.В. Тараканов, Ю.Н. Ильичев // Лесн. хоз-во. - 2008. - № 2. - С. 36-38.
- Лаур, Н.В. Плюсовые деревья и насаждения Карелии / Н.В. Лаур // Лесной вестник. - 2012. - № 2 (85). -С.14-22.
- Раевский, Б.В. Селекционно-генетическая оценка плюсовых деревьев сосны обыкновенной в Карелии / Б.В. Раевский, К.К. Куклина, М.Л. Щурова // Тр. Карельского НЦ РАН. - 2020. - № 3. - С. 45-59.
- Федорков, А.Л. Состояние экспериментальных культур сосны скрученной в Республике Коми / А.Л. Федорков, Л.Н. Гутий // Вестник Института биологии Коми НЦ Уральского отделения РАН. - 2017. - № 2 (200). - С. 25-31.
- Путенихин, В.П. Лесоводственные особенности некоторых североамериканских сосен при интродукции в Башкирском Предуралье / В.П. Путенихин, М.Ю. Шарипова, И.Е. Дубовик // Естественные и технические науки. - 2019. - № 10(136). - С. 177-180.
- Путенихин, В.П. Методы сохранения генетической гетерогенности при создании искусственных «популяций» лесообразующих видов / В.П. Путенихин, Г.Г. Фарукшина // Хвойные бореальной зоны. - 2007. - Т. 24. -№ 2-3. - С. 272-278.
- Vasilyeva, G.V. Artificial crosses and hybridization frequency in five-needle pines / G.V. Vasilyeva, S.N. Goroshkevich // Dendrobiology. - 2018. - Vol. 80. - Р. 123-130.
- Zhuk, E.A. Growth and reproduction in Pinus sibirica ecotypes from Western Siberia in a common garden experiment / E.A. Zhuk, S.N. Goroshkevich // New Forests. - 2018. - Vol. 49. - № 2. - P. 159-172.
- Видякин, А.И. Постгляциальная миграция и феногеография популяций сосны обыкновенной (Pinus sylvestris L.) на северо-востоке Русской равнины на северо-востоке Русской равнины / А.И. Видякин, С.Н. Санников, И.В. Петрова // Известия РАН. Сер. биологическая. - 2014. - № 3. - С. 304.
- Генетическая изменчивость, структура и дифференциация популяций сосны обыкновенной (Pinus sylvestris L.) на северо-востоке Русской равнины по данным молекулярно-генетического анализа / А.И. Видякин, С.В. Боронникова, Ю.С. Нечаева [и др.] // Генетика. - 2015. - Т. 51. - № 2. - С. 1401-1409.
- Семериков, Л.Ф. Популяционная структура древесных растений / Л.Ф. Семериков. - М. : Наука, 1986. -
- Мамаев, С.А. Формы внутривидовой изменчивости древесных растений на Урале (на примере сем. Pinaciae) / С.А. Мамаев. - М. : Наука, 1972. - 284 с.
- Изучение имеющихся и создание новых географических культур. - Пушкино : Госкомлес СССР, 1972. - 52 с.
- Shutyaev, A.M. Genetics subdivisions of the range of Scots Pine (Pinus sylvestris L.) Based on a Transcontinental Provinance Experiment / А.М. Shutyaev, M. Giertych // Silvae Genetica. - 2000. - Vol. 49. - P. 137-151.
- Shutyaev, A.M. Scots Pine (Pinus sylestris L.) in Eurasia - A Map Album of Provenance Site Interactions / A.M. Shutyaev, M. Giertych. - Kornik, Poland : Institute of Dendrology, 2003. - 266 р.
- Шутяев, А.М. Методика выделения и изучения сортов-популяций древесных видов на экологической основе / А.М. Шутяев. - Воронеж : ЦНИИЛГиС, 1992. - 19 с.
- Родин, А.Р. О проблемах изучения географических культур основных лесообразующих пород / А.Р. Родин, А.Е. Проказин // Лесное хоз-во. - 1996. - № 4. - С. 16-18.
- Кузьмин, С.Р. Отбор перспективных климатипов сосны обыкновенной в географических культурах разных лесорастительных условий / С.Р. Кузьмин, Н.А. Кузьмина // Лесоведение. - 2020. - № 5. - С. 451-465.
- Новикова, Т.Н. Изменчивость показателей развития кроны у потомств сибирских климатипов сосны обыкновенной / Т.Н. Новикова // Лесной журнал. - 2013. - № 4. - С. 22-29.
- Мельник, П.Г. Географическая изменчивость лиственницы в фазе приспевания / П.Г. Мельник, Н.Н. Ка-расев // Лесной вестник. - 2012. - № 1 - С. 60-73.
- Мельник, П.Г. Результат выращивания климатипов лиственницы в географических культурах северо-восточного Подмосковья / П.Г. Мельник, М.Д. Мерзленко, С.Л. Лобова // Вестник Алтайского государственного аграрного университета. - 2016. - № 2 (136). - С. 62-67.
- Мерзленко, М.Д. Результаты выращивания климатипов лиственницы в географических культурах западного Подмосковья / М.Д. Мерзленко, П.Г. Мельник, А.А. Коженкова // Вестник Алтайского государственного аграрного университета. - 2018. - № 1 (159). - С. 72-77.
- Лесоводственный опыт выращивания культур лиственницы в центре Русской равнины [Электронный ресурс]. / М.Д. Мерзленко, П.Г. Мельник, Ю.Б. Глазунов [и др.]. // Лесохоз. информ. - 2019. - № 4. - С. 55-66.
- Кузьмина, Н.А. Анализ лесосеменного районирования сосны обыкновенной в Средней Сибири / Н.А. Кузьмина, С.Р. Кузьмин // Хвойные бореальной зоны. - 2012. - Т. 29. - № 1-2. - С. 111-113.
- Федорков, А.Л. Лесосеменное районирование сосны обыкновенной на севере Европы / А.Л. Федорков // Сибирский лесной журнал. - 2020. - № 2. - С. 63-68.
- Жигунов, А.В. Лесные плантации триплоидной осины, созданные посадочным материалом in vitro / А.В. Жигунов, Д.А. Шабунин, О.Ю. Бутенко // Вестник Поволжского ГТУ. Сер.: Лес. Экология. Природопользование. - 2014. - №. 4 (24). - С. 21-30.
- Content and immunohistochemical localization of hormones during in vitro somatic embryogenesis in long-term proliferating Larix sibirica cultures / I.N. Tretyakova, M.E. Park, A.S. Kazachenko, A.S. Shuklina, G.R. Kudoyarova, G.R. Akhiyarova, A.V. Korobova, S.U. Veselov // Plant cell, tissue and organ culture. - 2019. - Т. 136. - № 3. - P. 511-522.
- Машкина, О.С. Технология долгосрочного хранения в культуре in vitro ценных генотипов березы и выращивание на ее основе посадочного материала / О.С. Машкина, Т.М. Табацкая, Н.И. Внукова // Биотехнология. -2019 - Т. 35. - № 3. - С. 57-67.
- Молекулярно-генетическая и цитогенетическая оценка перспективных гибридов и размноженных in vitro клонов тополя и осины / О.С. Машкина, Т.П. Федулова, Т.М. Табацкая [и др.] // Вестник ВГУ, серия: химия, биология, фармация. - 2016. - № 2. - С. 60-69.
- Development of F1 hybrid population and the high-density linkage map for European aspen (Populus tremula L.) using RADseq technology / A.V. Zhigunov, P.S. Ulianich, M.V. Lebedeva, P.L. Chang, S.V. Nuzhdin, E.K. Potokina // BMC plant biology. - 2017. - Vol. 17. - № 1. - P. 180.
- Анализ генетических локусов, влияющих на хозяйственно-ценные признаки осины (Populus tremula L.) в различных географических условиях / М.В. Лебедева, Г.М. Кукуричкин, Е.К. Потокина, А. Жигунов // Биотехнология в растениеводстве, животноводстве и ветеринарии : сб. тез. XVIII Всерос. конф. молодых учёных, посвящ. памяти академика РАСХН Г.С. Муромцева. - 2018. - С. 148-149.
- Обнаружение таксуюннанина С в суспензионной культуре клеток тиса канадского (Taxus canadensis) / Д.В. Кочкин, Е.Б. Глоба, Е. Демидова, В.В. Гайсинский, Вл.В. Кузнецов, А.М. Носов // Доклады Академии наук. -2019. - Т. 485. - № 3. - С. 374-376.
- Разработка микросателлитных маркеров лиственницы сибирской (Larix sibirica Ledeb.) на основе полногеномного de novo секвенирования / Н.В. Орешкова, Ю.А. Путинцева, В.В. Шаров, Д.А. Кузьмин, К.В. Крутов-ский // Генетика. - 2017. - Т. 53. - № 11. - P. 1278-1284.
- Postgenomic technologies in practical forestry: development of genome-wide markers for timber origin identification and other applications [Электронный ресурс ]/ K.V. Krutovsky, Y.A. Putintseva, N.V. Oreshkova,
- E.I. Bondar, V.V. Sharov, D.A. Kuzmin // Forest Engineering Journal. - V. 9. - 2019. - № 1. - P. 9-16 - Режим доступа: https://doi.org/10.12737/article_5c92016b64af27.15390296.
- Assembling of the Siberian larch mitochondrial genome using long nucleotide sequence reads, the largest currently known mitogenome/ Y.A. Putintseva, E.I. Bondar, V.V. Sharov, E.P. Simonov, N.V. Oreshkova, D.A. Kuzmin, M.G. Sadovsky, K.V. Krutovsky // 5th International Conference on Plant Genetics, Genomics, Bioinformatics, and Biotechnology (PlantGen 2019) (June 24-29, 2019, Novosibirsk, Russia). - 2019. - P. 163.
- Разработка ядерных микросателлитных маркеров с длинными (трех-, четырех-, пяти- и шестинуклеотид-ными) мотивами для трех видов лиственницы на основе полногеномного de novo секвенирования лиственницы сибирской (Larix sibirica Ledeb.) / Н.В. Орешкова, Е.И. Бондар, Ю .А. Путинцева, В.В. Шаров, Д.А. Кузьмин, К.В. Крутовский // Генетика. - 2019. - Т. 55. - № 4. - С. 418-425.
- Постгеномные технологии в практическом лесном хозяйстве: разработка полногеномных маркеров для идентификации происхождения древесины и других задач / К.В. Крутовский, Ю.А. Путинцева, Н.В. Орешкова, Е.И. Бондар, В.В. Шаров, Д.А. Кузмин // Лесотехнический журнал. - 2019. - Т. 9. - № 1 (33). - С. 9-16.
- Драгавцев, В.В. Эколого-генетическая организация полигенных признаков растений и теория селекционных индексов. / В.А. Драгавцев // Молекулярная и прикладная генетика. - 2009. - Т. 9. - С. 7-13.
- Драгавцев, В.А. Новая система регуляции у растений и необходимость создания селекционного фитотрона в РФ / В.А. Драгавцев // Журнал технической физики. - 2018. - Т. 88. - № 9. - С. 1331-1335.
- Левонтин, Р. Генетические основы эволюции / Р. Левонтин. - М. : Мир, 1978. - 352 с.
- Шмальгаузен, И.И. Избранные труды. Организм как целое в индивидуальном и историческом развитии / И.И. Шмальгаузен. - М. : Наука, 1982. - 383 с.
- 100. Гинзбург, Э.Х. Описание наследования количественных признаков / Э.Х. Гинзбург. - Новосибирск : Наука, 1984. - 250 с.
- 101. Драгавцев, В.А. Введение в теорию эколого-генетической организации полигенных признаков растений и теорию селекционных индексов / В.А. Драгавцев, Н.В. Кочерина. - СПб. : Агрофиз. НИИ, 2008. - 86 с.
- 102. Милютин, Л.И. Развитж лесной генетики в России / Л.И. Милютин, Е.Н. Муратова, А.Я. Ларионова // Сибирский лесной журнал. - 2018. - № 1. - С. 3-15.
- 103. Попов, Е.Б. Три кита эконики: Истоки и перспективы нового направления в общей биологии / Е.Б. Попов, В.А. Драгавцев, С.И. Малецкий. - СПб. : Издательско-полиграфическая ассоциация высших учебных заведений, 2020. - 132 с.
