Life forms and architectural models of some species of Campanula subgenus (Campanula L., Campanulaceae)
")
Author: Akbarova A., Salimov R.
Journal: Бюллетень науки и практики @bulletennauki
Section: Биологические науки
Article in issue: 6 т.10, 2024.
Free access
Campanula L. is the largest genus of Campanulaceae family and comprises about 400 (as well as 110 species in Caucasus and 46 species in Azerbaijan flora) herbaceous perennial, sometimes annual and biennial monocarp species distributed in the temperate zone of the northern hemisphere. Most of Campanula species are cosmopolitan. The species of this genus have long been used in the national economy, primarily as decorative plants, but also as pollinator, melliferous, food and medicinal plants. The genus Campanula L. yet does not have a modern taxonomic classification, although many species are of high horticultural value, and many are rare and endangered. The only monograph was published by A. de Candolle (1830). The final systematic composition of the genus was given in Flora of the Caucasus by M. E. Oganesian and V. V. Shvanova (2008). Representatives of this genus grows in various environmental conditions: forests, meadows, rocks and rarely in deserts and semi - deserts. Most of them are numerous and varied in high mountains, in the subalpine and alpine belts.
Species, genus, species systematics, taxonomy
Short address: https://sciup.org/14130178
IDR: 14130178 | UDC: 582.992 | DOI: 10.33619/2414-2948/103/05
Жизненные формы и архитектурные модели некоторых видов подрода Campanula (Campanula L., Campanulaceae)
Campanula L. - самый крупный род семейства Campanulaceae включает около 400 видов (в том числе 110 во флоре Кавказа и 46 во флоре Азербайджана) травянистых многолетних, иногда однолетних и двулетних одноплодных видов, распространенных в умеренной зоне северного полушария. Большинство видов колокольчиков космополитны. Виды этого рода издавна используются в народном хозяйстве, прежде всего как декоративные растения, а также как пыльценосные, медоносные, пищевые и лекарственные растения. Род Campanula L. пока не имеет современной таксономической классификации, хотя многие виды имеют высокую садоводческую ценность, а многие из них редки и находятся под угрозой исчезновения. Единственную монографию опубликовал А. де Кандоль (1830). Окончательный систематический состав рода приведен в «Конспекте Флоры Кавказа» М. Е. Оганесяна и В. В. Шванова (2008). Представители этого рода произрастают в различных природных условиях: лесах, лугах, скалах и редко в пустынях и полупустынях. Большинство из них многочисленны и разнообразны в высокогорьях, в субальпийском и альпийском поясах.
Text of the scientific article Life forms and architectural models of some species of Campanula subgenus (Campanula L., Campanulaceae)
Бюллетень науки и практики / Bulletin of Science and Practice
The concept of life form was first introduced to science in the 80s of the 19th centuries by the famous Danish botanist E. Warming. He first focused on the adaptation of the vegetative sphere of the plant to the environmental conditions. A plant's life form is its habitus, which is related to the current and past characteristics of the environment and its growth rhythm [6, 7].
Architectural model — is a new term has appeared during the past several decades to characterize the structure of the plant body. Studies about Architectural models firstly began with the examination of tropical trees. Observations of structure and its growth on tropical trees indicate that each species has its own structure of growth (model) and it can be seen on a young plant. The major characteristics include the following features:
-
1) trunk monopodial or sympodial;
-
2) trunk growing rhythmically or continuously;
-
3) branches orthotropic or plagiotropic; sympodial determined or indetermined;
-
4) flowering lateral or terminal.
Each model was named after the name of appropriate scientist. For example, model of Holttum — determinate trunk, terminal inflorescence, no branches except those within inflorescence; model of Corner — monopodial trunk, lateral branches within inflorescence; model of Tomlinson — sympodial branching sequence with each orthotropic sympodial unit born on the proximal portion of a previous unit, sympodial units indeterminate or determinate etc.
The different models are distinguished by the presence of certain features such as: monopodial or sympodial trunk, continuous or rhythmical growth, orthotropic or plagiotropic branches, and terminal or lateral flowering, etc. It is known that the structure of herbaceous plants differs very much from trees and shrubs. The existing stem and branches of woody plants demonstrate the entire body of the plant including the dead tissue in the stem, whereas herbaceous plants shed most above-ground parts of their body annually. Herbaceous plants were studied by Jeannoda-Robinson (1977) who showed that some of the same architectural models found in woody plants can be found within herbaceous plants. She studied 180 species from 48 different families and found that their structure is basically similar.
Also Russian scientist T. V. Shulkina studied the life forms and architectural models of the species included in the subsections indicated in the flora of the USSR [2, 5, 10].
Material and methods
As material we used plant samples collected during expeditions organized to different regions of Azerbaijan in 2018-2022, as well as herbarium materials stored in the herbarium fund of the Institute of Botany. During the field trips were also studied the habitat and life forms of species and comparisons were made with literature data.
Results
Among the numerous systems proposed for the life forms of plants, one of the most widely used to date is the classification of the Danish botanist K. Raunkier (1918) [4].
This classification is based on the adaptation of the plants to the unfavorable times of the year (cold and dry) and the condition of the regeneration shoots in the plant depending on the level of the substrate and the snow cover. He introduced 5 main life forms: phanerophytes, chamephytes, hemicryptophytes, cryptophytes, and therophytes.
In phanerophytes, the shoots spend the winter or dry season in the open, they are high above the ground (trees, shrubs, tree-like lianas, epiphytes). They are usually covered with scales that protect the growth cone and young leaf bases from moisture loss. In chamephytes, the shoots are 20-30 cm from the ground. located at height (shrubs, semi-shrubs, creeping plants). In cold and temperate climates, these shoots overwinter under snow. In hemicryptophytes, shoots are located at ground level, immersed in a covering formed from leaves and dead plant debris (herbaceous plants).
Cryptophytes include either geophytes (rooted, bulbous, tuberous plants) whose regeneration shoots are located somewhat deep in the soil, or hydrophytes whose regeneration shoots spend the winter in water. In therophytes, by the end of the season, all parts of the plant are destroyed and there are no overwintering shoots (annual plants).
Most species of Campanula L. subgenus belong to hemycryptophytes, only C. bayerniana belong to chamephytes .
The life forms of bluebells are extremely diverse, which corresponds to the high endurance of the genus and the significant diversity of their habitats (subalpine and alpine mountain belts, forests, meadows, rocks, partly steppes and deserts) [1].
According to the information of G.I. Khalipova (2000) and based on I. G. Serebryakov's (1964) classification of life forms, species of this subgenus are divided into the following classes and subclasses [7, 8]:
-
a. Herbaceous polycarps
Class: Herbaceous polycarps with assimilating corms of the non-succulent type
I subclass: having a spindle-shaped root ( C. glomerata, C. alliariifolia )
II subclass: those with brush-shaped roots and short rhizomes ( C. bononiensis, C. latifolia )
VII subclass: those with additional roots ( C. rapunculoides )
-
b. Monocarp grasses
Class: Monocarpous herbs with assimilating corms of the non-succulent type
Subclass: Biennials ( C. sibirica )
The large number of life forms of Campanula species is related to the diversity of their habitats: (forest, meadow, steppe, alpine and subalpine zones in the mountains, etc.)
According to the research of A. B. Bezdelevoi and T. A. Bezdelevoi (2006), the species of this subgenus in the adult generative state are divided into the following life forms:
-
1. A biennial summer-green herbaceous monocarp with a semi-rosette erect stem, growing monopodially ( C. sibirica ).
-
2. Perennial summer-green, herbaceous, sympodially branching polycarp with semi-rosetted erect stem with long rhizome-spindle-brush-like root ( C. bononiensis, C. latifolia ).
-
3. Perennial summer green, herbaceous short-rooted, sympodial branching polycarp with a semi-rosetted erect stem with spindle-brush-shaped root ( C. bononiensis, C. latifolia ).
-
4. Perennial summer green, herbaceous, sympodially branching polycarp with a semi-rosetted erect stem with spindle-brush-shaped root and spike-shaped appendages ( C. alliariifolia ) [9].
-
5. Perennial summer green, herbaceous, spindle-brush-stolon-forming sympodially branching polycarp with semi-rosette erect stem ( C. glomerata ) [3, 9].
Campanula L. genus is characterized by various architectural models, which indicates the genus is polytypic for architecture. Species of this genus grow in various environmental conditions: in forests, in meadows, in the steppe and rarely in semi — desert conditions, but they occur mostly in subalpine and alpine zones. The large set of life — forms explain why representatives of the genus inhabit many different environmental conditions [5].
-
C. sibirica L. grow in dry subalpine meadow, in old fields, or in margins of forest. This species develops a rosette of leaves during the first year and flowering stem from the epicotyl axis with elongated internodes during the second year. Panicle — like inflorescence is characteristic for
C. sibirica. C. sibirica belongs to the model of Holltum. As an exception, after fruiting some axillary meristems stay alive and produce new shoot the next year.
-
C. latifolia L., C. bononiensis L., C. rapunculoides L., C. odontosepala Boiss. These species grow in mixed, broad — leaved or dark coniferous forests; C. latifolia also grows among tall grasses and shrubs in the subalpine zone and in dry meadows. All 4 species are shade tolerant. They have a rosette of leaves in the juvenile stage and erect leafy elongated stem from the primary (epicotyl) axis. Inflorescences are panicles and racemes. Renewal buds are developed at the base of the flowering stem. These plants have dormant buds covered by scales situated at the base of the stem and on the hypocotyl. All species have one model — that of Tomlinson.
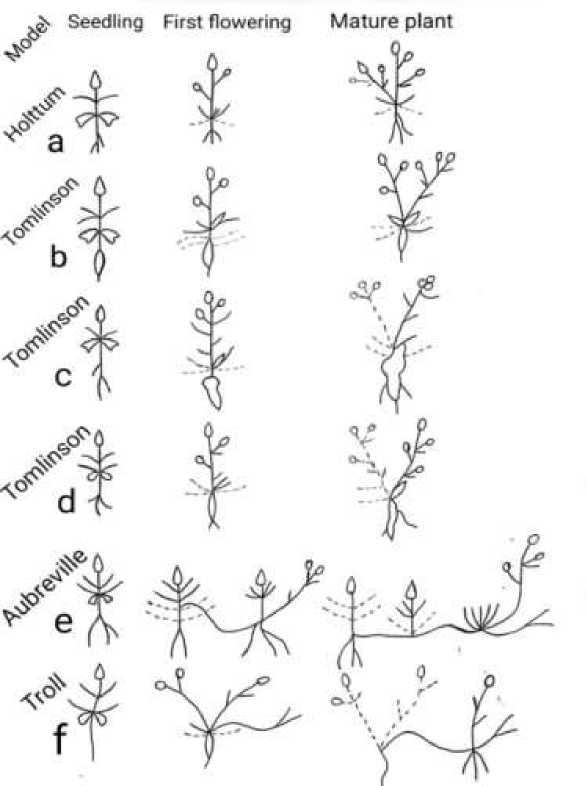
Figure. Architectural models of some species of Campanula subgenus a. Section: Campanula ( C. sibirica ) b. Section: Campanula ( C. latifolia , C. odontosepala , C. rapunculoides , C. bononiensis ) c. Section: Involucratae ( C. glomerata ) d. Section: Cordifoliae ( C. alliariifolia ) e. Section: Cordifoliae ( C. sarmatica , C. collina ) f. Section: Symphyandriformes ( C. kolenatiana , C. bayerniana )
-
C. glomerata L. — grows mostly at the edge of open broad — leaved forests, in meadows and rarely among shrubs. This species is not shade tolerant. C. glomerata develops a rosette of leaves in the juvenile stage and elongated stems, usually unbranched by anthesis. Later growth develops from the axils of lower leaves. Plants have a tap root and a short horizontal rhizome; a sympodial system of branching. This is the model of Tomlinson.
-
C. alliariifolia Willd. — grow mostly on limestone or dolomite rocks, cliffs, rocky soil in mid mountain and subalpine belts. During the juvenile stage, this specie develops a rosette of leaves.
(ос) CD
The epicotyl axis is erect, leafy 10-120 cm tall with a raceme. After first flowering and fruiting, the reproductive stems die to base. C. alliariifolia do not have any above ground parts in winter, only buds, covered by scales. Next year elongated stems are developed at the proximal portion of previous units. Model of Tomlinson.
-
C. collina Sims., C. sarmatica Ker Gawl. C. collina grows in meadows in subalpine (sometimes alpine) zones; C. sarmatica grows in shallow soil, in cliffs, cracks and talus, mostly in subalpine and alpine zones. The epicotyl axis of these species never comes to flower. The primary rosette gives rise to daughter rosettes and elongated reproductive stems develop from axillary meristem in these daughter rosettes. C. collina develops primary and axillary rosettes and also elongated plagiotropic leafless stems (rhizomes) just on the ground surface, which in their turn terminate in rosettes, but C. sarmatica do not produce long plagiotropic rhizomes. C. collina have monopodial growth from early stages of the ontogenesis, becoming sympodial when the main axis dies. C. sarmatica have a model of Tomlinson, C. collina — fragment of the model of Aubreville.
-
C. kolenatiana C. A. Mey., C. bayerniana Rupr. These species grow in forest zone (sometimes also in subalpine zone), in rocky soil, limestone cliffs, cracks and basalt rocks. They do not occur in alpine meadows, in steppe and among tall grasses. In the first year these plants have a rosette of leaves; in the second-year plants develop axillary rosettes. Reproductive stems derive from the epicotyl axis and from the axillary meristem of lower rosette leaves. The main axis and axillary stems have an ability to terminate in flowers. The young plants have a model of Troll (monopodial), but mature plants have a model of Troll (sympodial) (Figure).
Discussions
The current studies showed that Campanula species exhibit a diversity of architectural models and life forms and that is possible to select morphological characters of vegetative organs that are stable with the species at different taxonomic levels. The most stable and common vegetative character for the genus is the presence of rosette of leaves in the early stage of ontogenesis.
Table
|
Section |
Species |
Distribution (in Azerbaijan) |
Architectural models |
|
Campanula |
C. sibirica L. |
GC (Quba) GC (east) |
Holltum |
|
Campanula |
C. latifolia L. |
GC (Quba) GC (east) |
Tomlinson |
|
Campanula |
C. bononiensis L. |
throughout Azerbaijan |
Tomlinson |
|
Campanula |
C. rapunculoides L. |
throughout Azerbaijan |
Mc. Clure |
|
Campanula |
C. odontosepala Boiss. |
Lankaran |
Tomlinson |
|
Involucratae (Fomin) Charadze |
C. glomerata L. |
throughout Azerbaijan |
Tomlinson |
|
Cordifoliae (Fomin) Charadze |
C. alliariifolia Willd. |
LC (north) |
Tomlinson |
|
Cordifoliae (Fomin) Charadze |
C. collina Sims. |
GC (Quba) GC (east) GC (west) LC (north) |
Aubreville |
|
Cordifoliae (Fomin) Charadze |
C. sarmatica Ker Gawl. |
GC (Quba) GC (east) GC (west) |
Tomlinson |
|
Symphyandriformes (Fomin) Charadze |
C. kolenatiana C. A. Mey. |
GC (west) |
Troll |
|
Symphyandriformes (Fomin) Charadze |
C. bayerniana Rupr. |
Nakhchivan (Ordubad) |
Troll |
Note: GC — Great Caucasus; LC — Lesser Caucasus.
Some species might change their architectural model during their lifespan, for instance, starting as a monopodial model plant can switch to sympodial ramification loosing dead parts od structure. İt means that plant can have 2 different models: one for young plant and second for mature plant.
Some species of the genus have an epicotyl axis which terminates in an inflorescence after the production of some nodes (sympodial structure), and also some species have a monopodial branching structure, where the epicotyl axis never terminates in a flower.
References Life forms and architectural models of some species of Campanula subgenus (Campanula L., Campanulaceae)
- Федоров А. А. Флора Азербайджана. Т. 8. Баку, 1961. С. 126.
- Шулькина Т. В. Архитектурные модели в семействе Campanulaceae S. Str., их география и возможные пути преобразования // Ботанический журнал. 1988. Т. 73. №1. С. 3-16.
- Аллаярова И. Н., Миронова Л. Н., Мингажева А. М. Жизненные формы растений на примере рода Колокольчик (Campanula L.). Уфа, 2012. 26 с. EDN: UHMQGX
- Ellenberg H., Mueller-Dombois D. A key to Raunkiaer plant life forms with revised subdivisions. 1967.
- Шулькина Т. В. Типы жизненных ферм и их значение для систематики Campanula L. // Ботанический журнал. 1977. Т. 62. №8. C. 1102-1114.
- Серебрякова Т. И. Учение о жизненных формах растений на современном этапе // Итоги науки и техники. Сер. Ботаника. 1972. Т. 1. С. 84-169.
- Серебряков И. Г. Экологическая морфология растений: Жизненные формы покрытосеменных и хвойных. М.: Высш. школа, 1962. 378 с.
- Халипова Г. И. Колокольчики. М.: Астрель, 2005. 141 с.
- Аллаярова, И. Н., & Миронова, Л. Н. (2011). Онтогенез некоторых представителей рода Campanula L. при культивировании в условиях Башкирского Предуралья. Региональные геосистемы, 15(9-1 (104)), 140-145.
- Шулькина Т. В. Жизненные формы и архитектурные модели видов рода Campanula L. (Campanulaceae) и их связь с таксономией. М.; СПб.: Т-во науч. изд. КМК, 2015. 99 с.
