Макрозообентос эстуария малой лососевой реки острова Сахалин на примере р. Горная

Автор: Корнеев Е. С., Лабай В. С., Живоглядова Л. А., Березова О. Н., Шарлай О. Б., Абрамова Е. В., Ахмадеева Е. С.
Журнал: Вестник Мурманского государственного технического университета @vestnik-mstu
Рубрика: Биологические науки
Статья в выпуске: 2 т.27, 2024 года.
Бесплатный доступ
Эстуарии как связующее звено между пресноводными и морскими экосистемами в лососевых реках играют особую роль. От условий в эстуарных зонах рек зависит успех адаптации молоди лососевых рыб при переходе к морскому периоду жизни. Работа продолжает начатый ранее цикл публикаций по рекам о-ва Сахалин. На примере одного из малых водотоков юго-восточной части острова рассматриваются гидрологические параметры и особенности структурной организации сообществ макрозообентоса эстуария, не типичного по своим характеристикам. В отсутствии штормовых заплесков проникновения морских вод в эстуарий практически не происходит. Воды с соленостью более 12 psu отмечаются только в устье у дна во время прилива, в отлив эстуарий полностью заполняется водой с соленостью 0,1 psu и ниже. Исследования гидрологического режима и донной фауны были проведены в июле 2022 г. Пробы зообентоса отбирали по продольному профилю эстуария на пяти разрезах (суммарно 75 проб). Работы показали, что большую часть эстуария заселяют виды, характерные для нижней ритрали водотоков острова. Высокая изменчивость среды обусловливает формирование в этом эстуарии молодых и нестабильных сообществ макрозообентоса, представленных быстроразмножающимися мелкоразмерными видами.
Донные сообщества, молодь лососей, кормовая база, гидрологический режим, эстуарий, bottom communities, juvenile salmon, food base, hydrological regime, estuary
Короткий адрес: https://sciup.org/142240265
IDR: 142240265 | УДК: 574.5 (571.64) | DOI: 10.21443/1560-9278-2024-27-2-126-141
Текст научной статьи Макрозообентос эстуария малой лососевой реки острова Сахалин на примере р. Горная
Корнеев Е. С. и др. Макрозообентос эстуария малой лососевой реки острова Сахалин на примере р. Горная. Вестник МГТУ. 2024. Т. 27, № 2. С. 126–141. DOI:
е-mail: , ORCID:
Korneev, E. S. et al. 2024. Macrozoobenthos in the estuary of the small salmon river of Sakhalin Island on example of the Gornaya River. Vestnik of MSTU, 27(2), pp. 126–141. (In Russ.) DOI:
В современной терминологии эстуарий рассматривается как водный объект в устьевой области реки, характеризующийся процессами смешения речных и морских водных масс ( Михайлов и др., 2009; Колпаков, 2017 ). Интерес исследователей к этим водным объектам обусловлен их особой функциональной ролью в системе морских и пресных водоемов, а также значением в жизнедеятельности многих морских и проходных видов ( Столяров, 2011 ; Колпаков, 2017 ). В эстуариях лососевых рек условия нагула и гидрологический режим определяют выживаемость молоди лососевых рыб на одном из важнейших этапов жизненного цикла – переходе из пресноводных систем в морские ( Волобуев и др., 2011; Изергин и др., 2018 ).
Настоящая работа продолжает цикл публикаций, посвященных особенностям организации донной фауны эстуариев лососевых рек о-ва Сахалин. Ранее на примере водотоков с хорошо выраженными эстуарными зонами (р. Мануй, бассейн Охотского моря и р. Сусуя, бассейн залива Анива) были показаны пространственная гетерогенность и изменение структуры сообществ макрозообентоса в градиенте солености и других факторов среды ( Лабай и др., 2022 ). В отличие от указанных водотоков многие малые реки о-ва Сахалин, сохраняющие в нижнем течении характер предгорного водотока, не имеют развитого эстуария. Затока соленых вод в такие водные объекты практически не происходит, структурирующая роль солености в их эстуариях слабо выражена или отсутствует. Работа посвящена описанию гидрологического режима и особенностей организации донных сообществ подобных эстуариев на примере одного из водотоков восточной части острова – реки Горная.
Река Горная является базовой для лососевого рыборазводного завода "ЛРЗ Горная" со средним выпуском молоди лососевых 25 млн шт., из которых 20 млн шт. приходится на кету. Сведения о гидрологическом режиме ее эстуария, количественных показателях донных сообществ, формирующих основу кормовой базы рыб, могут быть востребованы при оценке приемной емкости и иметь практическое значение при организации выпуска молоди рыб.
Район исследований
Река Горная берет свое начало с западного склона г. Дружеская (высота 523 м) Западно-Лисянского хребта и впадает в зал. Терпения Охотского моря. Протяженность основного русла реки составляет 33 км, площадь бассейна – 138 км2.
Долина нижнего течения р. Горная широкая – до полутора километров. Ширина реки на этом участке варьирует от 8 до 45 м. Русло реки извилистое, местами раздваивается, имеются галечные косы длиной от 10 до 50 м. Чередование плесов и перекатов хорошо выражено, скорость течения на плесах достигает 0,6 м/сек, на перекатах – 1,0 м/сек, глубина соответственно 0,15–0,6 м и 0,1–0,4 м.
В роли верхней границы эстуария выступает гравийно-галечный перекат, расположенный в 100 м ниже пересечения с автодорогой (рис. 1). В ходе активных процессов переформирования русла морфология дна и берегов эстуария постоянно меняется. На рис. 1 представлены спутниковые снимки 2005 г. ( а ) и 2022 г. ( б ), позволяющие оценить масштаб этих изменений. К 2022 г. в результате спрямления нижнего участка русла протяженность эстуария р. Горная сократилась почти в два раза, в настоящее время длина эстуарной зоны реки составляет 0,3 км.
Материалы и методы
Отбор и обработку проб макробентоса проводили в соответствии с принятыми методиками1. При отборе проб использовали складной бентометр (модификация бентометра Леванидова) с площадью 0,12 м2, фильтрующий конус (длина 0,6 м) изготовлен из мельничного газа с размером ячеи 220 мкм.
Бентосная съемка выполнена на пяти разрезах (рис. 2). Каждый разрез включал пять станций – рипаль у правого и левого берегов, фарватер и по обе стороны от фарватера.
1 Руководство по методам гидробиологического анализа поверхностных вод и донных отложений / под ред. В. А. Абакумова. Л. : Гидрометеоиздат, 1983. 240 с. ; Методические рекомендации по сбору и определению зообентоса при гидробиологических исследованиях водотоков Дальнего Востока России : методическое пособие. М. : ВНИРО, 2003. 95 с.
б
а

Рис. 1. Район работ, спутниковый снимок эстуария р. Горная: а – 2005 г., б – 2022 г.
Fig. 1. Scheme of the study area, satellite image of the Gornaya River estuary: a – 2005, б – 2022
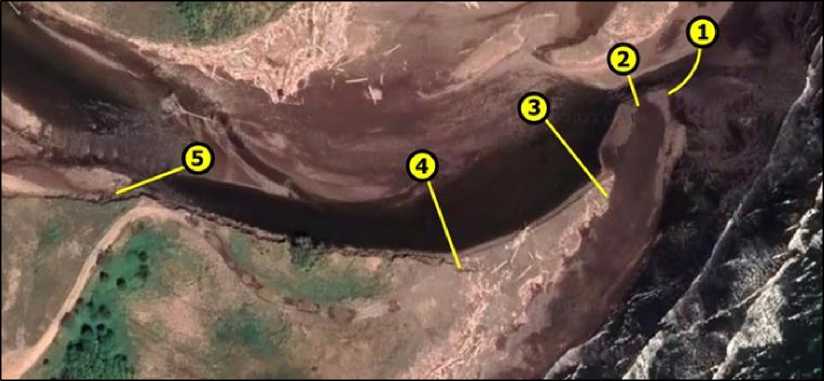
Рис. 2. Карта-схема разрезов на р. Горная
Fig. 2. Map-scheme of sampling transsects of the Gornaya River
Пробы на каждой станции отбирали в трех повторностях. Всего собрано 75 проб макробентоса (табл. 1). Фиксацию бентоса проводили 4%-м нейтрализованным формалином.
Таблица 1. Объем материала, собранного на р. Горная в июле 2022 г.
Table 1. The volume of material collected on the Gornaya River in July, 2022
|
Разрез |
Удаление от устья, км |
Максимальная глубина, м |
Элемент русла |
Количество станций |
Количество проб |
|
1 |
0,00 |
0,2 |
устье |
5 |
15 |
|
2 |
0,027 |
0,43 |
эстуарный плес |
5 |
15 |
|
3 |
0,075 |
0,57 |
эстуарный плес |
5 |
15 |
|
4 |
0,173 |
0,87 |
эстуарный плес |
5 |
15 |
|
5 |
0,279 |
0,62 |
перекат |
5 |
15 |
|
Всего |
75 |
||||
По всей толще воды на каждом разрезе параллельно с отбором проб бентоса с помощью мультипараметрического зонда Horiba U 5000G производили измерения солености водных масс (psu) и температуры (°C).
Структуру донных сообществ характеризовали по следующим параметрам: число видов ( S ); удельная численность (плотность) ( N , экз./м2); биомасса ( B , г/м2); частота встречаемости (ЧВ, %). При описании видового состава "видом" обозначали "низший определяемый таксон" ( Протасов, 2012 ). Удельную численность и биомассу определяли с учетом всех проб. Массовыми считались виды при ЧВ более 50 %.
Для классификационных и ординационных процедур в качестве меры обилия видов использовали показатель интенсивности метаболизма Q (кал/м2 х ч), связывающий оценки численностей и биомасс в единую переменную. Этот показатель эквивалентен энергетическим затратам на дыхание всех особей и определяется по формуле ( Кучерук и др., 1985; Мокиевский и др., 2012 )
q = к. в0.75. n'25, где Bi (г/м2) – удельная биомасса вида; Ni (экз./м2) – удельная плотность вида.
Коэффициент k для разноногих ракообразных (Amphipoda) принят равным 0,302; для равноногих ракообразных (Isopoda) – 0,133; для личинок насекомых (Diptera, Ephemeroptera, Plecoptera, Trichoptera, Coleoptera) – 0,189 ( Голубков, 2000; Алимов и др., 2013 ).
Оценку сходства между станциями и разделение их по сообществам проводили, используя индекс Чекановского ( Максимович и др., 1986 ). Станции отнесены к единому сообществу при превышении значения индекса 40 %. Расчет матриц и их кластеризацию выполняли в программе Statistica version 8. Кластеризацию исходных матриц осуществляли по методу невзвешенных парно-групповых средних (unweighted pair-group average).
Для описания структуры сообщества и определения доминирующих таксонов использовали коэффициент относительности (КО) – произведение относительной средней В (%) или показателя Q (%) на частоту встречаемости (%) ( Палий, 1961 ). Вид считали доминирующим (превалирующим), если значение КО находилось в интервале 1 000–10 000 (доля доминирующего вида от общей Q сообщества составляла более 10 % при ЧВ 100 %). Названия сообществ приведены по доминирующим видам.
Видовое разнообразие водных сообществ оценивали с использованием индекса видового разнообразия (энтропийного индекса) Шеннона – Винера ( I ВО , бит/экз.) ( Shannon, 1948 ). Размерную структуру сообществ и степень ее трансформации оценивали ABC-методом (abundance-biomass comparison method) ( Warwick, 1986 ) по ABC-индексу ( Meire et al., 1990 ).
Для определения типа питания отдельных видов макрозообентоса применяли следующую номенклатуру трофических группировок: Br – макроизмельчитель, De – грунтофаг, Gr – соскребатель, Dt – собирающий детритофаг, Pr – хищник, Sc – падальщик, Su – сестонофаг (фильтратор). Для некоторых видов характерно совмещение нескольких типов питания, что выражалось в смешанной характеристике, например: De, Dt или Dt, Su или Dt, Br, Sc.
Результаты исследований
Гидрологическая характеристика эстуария р. Горная
В период проведения работ (7.07.2022) скорость течения на обследованном участке русла варьировала от 0,008 до 1,035 м/с. Максимальные значения отмечали в устье. Глубина изменялась от 0,05 до 0,87 м, батиметрический профиль вдоль продольной оси эстуария представлен на рис. 3. Там же показано вертикальное распределение солености водных масс.
В прилив морские воды с соленостью до 12 psu отмечали у дна на первом и втором разрезах. Проникновения морских вод вверх по течению реки выше 20–30 м от устья не происходило. В отлив речной сток полностью вытеснил морские воды с эстуарной зоны и соленость на всех разрезах составляла 0,1 psu.
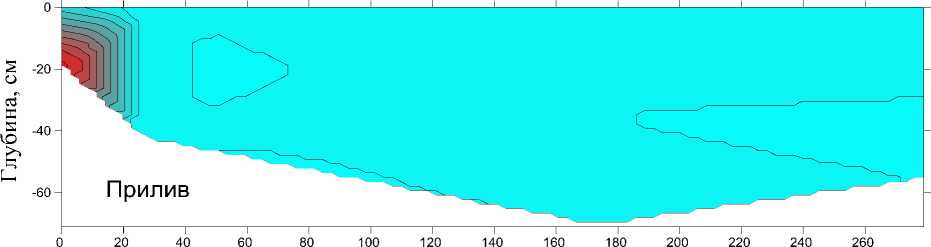
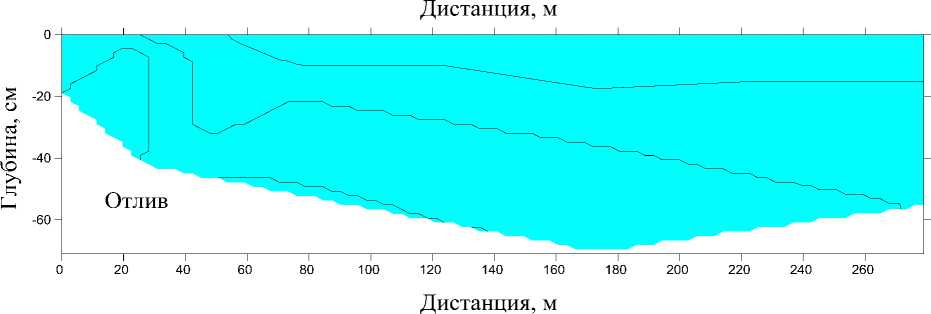
Рис. 3. Батиметрический профиль и вертикальное распределение солености водных масс вдоль эстуария р. Горная, 7 июля 2022 г. во время прилива (вверху) и отлива (внизу) Fig. 3. Bathymetric profile and vertical distribution of salinity of water masses along the estuary of the Gornaya River, July 7, 2022 during high tide (above) and low tide (below)

О 0,05 0,1 1 23456789 10 11
Видовой состав и особенности пространственного распределения макробентоса
В составе донной фауны отмечено 44 вида донных беспозвоночных (табл. 2). Основу видового списка формировали личики и куколки хирономид (22 вида). Второй по значимости группой таксоцена являлись личинки поденок – 10 видов, в составе прочих групп отмечено от 1 до 3 таксонов.
Таблица 2. Видовой состав макрозообентоса эстуария р. Горная, июль 2022 г. Table 2. Species of macrozoobenthos in the estuary of the Gornaya River, July 2022
|
№ |
Группа |
Вид |
Разрезы |
||||
|
1 |
2 |
3 |
4 |
5 |
|||
|
1 |
Amphipoda |
Ampelisca macrocephala Liljeborg, 1852 |
+ |
– |
– |
– |
– |
|
2 |
Amphipoda |
Eogammarus kygi (Derzhavin, 1923) |
+ |
+ |
+ |
+ |
+ |
|
3 |
Amphipoda |
Photis spasskii Gurjanova, 1951 |
+ |
– |
– |
– |
– |
|
4 |
Coleoptera |
Esolus sp. (larv.) |
– |
– |
– |
+ |
– |
|
5 |
Diptera |
Chironomidae gen. sp. (larv.) |
+ |
– |
– |
– |
– |
|
6 |
Diptera |
Chironomus (Lobochironomus) dorsalis Meigen, 1818 (larv.) |
– |
– |
+ |
+ |
– |
|
7 |
Diptera |
Chironomus sp. (pup.) |
– |
– |
+ |
+ |
– |
|
8 |
Diptera |
Corynoneura arctica Kieffer, 1923 (larv.) |
+ |
– |
– |
– |
+ |
|
9 |
Diptera |
Demicryptochironomus vulneratus (Zetterstedt, 1838) (larv.) |
– |
– |
+ |
– |
– |
|
10 |
Diptera |
Doncricotopus bicaudatus Saether, 1981 (larv.) |
– |
– |
+ |
– |
– |
|
11 |
Diptera |
Eukiefferiella gr. gracei (larv.) |
– |
– |
– |
+ |
– |
|
12 |
Diptera |
Glyptotendipes gripekoveni (Kieffer, 1913) (larv.) |
– |
– |
+ |
– |
– |
|
13 |
Diptera |
Hexatoma sp. (larv.) |
– |
– |
+ |
– |
– |
|
14 |
Diptera |
Orthocladius frigidus (Zetterstedt, 1838) (larv.) |
– |
– |
+ |
– |
– |
|
15 |
Diptera |
Orthocladius (Orthocladius) gr. saxicola (larv.) |
– |
+ |
+ |
– |
– |
|
16 |
Diptera |
Orthocladius sp. (pup.) |
— |
— |
+ |
— |
— |
|
17 |
Diptera |
Paratanytarsus inopertus Walker, 1856 (larv.) |
– |
+ |
– |
– |
– |
|
18 |
Diptera |
Polypedilum bicrenatum Kieffer, 1921 (larv.) |
– |
– |
+ |
+ |
+ |
|
19 |
Diptera |
Polypedilum (Polypedilum) pedestre (Meigen, 1830) (larv.) |
— |
— |
+ |
+ |
+ |
|
20 |
Diptera |
Polypedilum sp. (pup.) |
– |
– |
– |
– |
+ |
|
21 |
Diptera |
Procladius gr. choreus (larv.) |
— |
— |
— |
+ |
+ |
|
22 |
Diptera |
Psectrocladius (s. str.) bisetus Goetgh. (larv.) |
— |
— |
+ |
+ |
+ |
|
23 |
Diptera |
Psectrocladius (s. str.) z etterstedti Brundin, 1949 (larv.) |
– |
– |
+ |
– |
– |
|
24 |
Diptera |
Sergentia baueri Wülker et al., 1998 (larv.) |
— |
— |
+ |
— |
— |
|
25 |
Diptera |
Tanytarsus sp. (larv.) |
+ |
– |
+ |
+ |
+ |
|
Diptera |
Tanytarsus sp. (pup.) |
+ |
— |
+ |
+ |
+ |
|
|
26 |
Diptera |
Tokunagaia ambigua Makarchenko et Makarchenko, 2006 (larv.) |
— |
— |
+ |
— |
+ |
|
27 |
Ephemeroptera |
Baetis (Acentrella) sibiricus (Kazlauskas, 1963) (larv.) |
+ |
– |
– |
– |
+ |
|
28 |
Ephemeroptera |
Baetis (Baetis) sp. (larv.) |
+ |
+ |
+ |
+ |
+ |
|
29 |
Ephemeroptera |
Baetis ursinus Kazlauskas, 1963 (larv.) |
– |
– |
– |
+ |
+ |
|
30 |
Ephemeroptera |
Ecdyogymnurus aspersus Kluge, 1980 |
+ |
– |
– |
– |
+ |
|
31 |
Ephemeroptera |
Ecdyonurus sp. |
– |
– |
– |
– |
+ |
|
32 |
Ephemeroptera |
Epeorus sp. (larv.) |
– |
– |
– |
– |
+ |
|
33 |
Ephemeroptera |
Epeorus (Belovius) sp. (larv.) |
– |
– |
– |
– |
+ |
|
34 |
Ephemeroptera |
Ephemerella aurivillii Bengtsson, 1908 (larv.) |
+ |
+ |
+ |
– |
+ |
|
35 |
Ephemeroptera |
Neoleptophlebia japonica (Matsumura, 1931) (larv.) |
– |
– |
+ |
– |
+ |
|
36 |
Ephemeroptera |
Rhithrogena (Rhithrogena) gr. lepnevae (larv.) |
– |
+ |
– |
– |
– |
|
37 |
Isopoda |
Synidotea cinerea Gurjanova, 1933 |
+ |
– |
– |
– |
– |
|
38 |
Nematoda |
Nematoda gen. sp. |
+ |
– |
+ |
– |
+ |
|
39 |
Oligochaeta |
Specaria josinae (Vejdovský, 1884) |
+ |
+ |
+ |
+ |
+ |
|
40 |
Plecoptera |
Amphinemura borealis (Morton, 1894) (larv.) |
– |
– |
– |
– |
+ |
|
41 |
Plecoptera |
Sweltsa sp. (larv.) |
– |
+ |
– |
– |
– |
|
42 |
Plecoptera |
Suwallia sp. (larv.) |
+ |
+ |
+ |
– |
+ |
|
43 |
Trichoptera |
Apatania sp. (larv.) |
– |
– |
+ |
– |
+ |
|
44 |
Trichoptera |
Ceratopsyche orientalis Martynov, 1934 (larv.) |
– |
– |
+ |
– |
– |
|
Итого |
15 |
9 |
26 |
14 |
24 |
Длина видового списка по разрезам изменялась от 9 до 26 видов – с минимумом на втором и максимумом на третьем створах. В устьевой зоне эстуария были отмечены специфические морские беспозвоночные, занесенные с приливными водами, – амфиподы Ampelisca macrocephala , Photis spasskii и изопода Synidotea cinerea . Все остальные виды являлись обычными представителями нижней ритрали рек о-ва Сахалин. Массовым видом с частотой встречаемости 72 % являлась олигохета Specaria josinae .
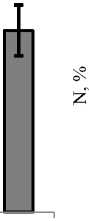
Распределение количественных показателей вдоль продольной оси эстуария имело схожую динамику как по численности, так и по биомассе (рис. 4). От нижнего створа к верхнему плотность и биомасса беспозвоночных увеличивались. Этот рост достигал одного порядка по численности – с 36 ± 5,8 до 342 ± 47,7 экз./м2, и почти порядка по биомассе – с 0,064 ± 0,009 до 0,502 ± 0,059 г/м2.
В устье реки и на створе выше основу плотности формировали олигохеты (38–70 %). На третьем разрезе наблюдалась смена доминирующих групп и основной вклад в общую плотность вносили двукрылые (63 %). Схожей была ситуация и на четвертом разрезе, где также доминировали двукрылые (78 %). На верхнем створе численность двукрылых резко сократилась и доминирующее положение вновь заняли олигохеты (рис. 4, б ).
Структура доминирования по биомассе изменялась аналогичным образом (рис. 4, в , г ) с увеличением роли амфипод. В устье реки и створом выше превалировали поденки (49 %) и амфиподы (35 %). На третьем разрезе произошла смена доминант и основной вклад в общую биомассу вносили двукрылые (37–67 %). На верхнем разрезе структура доминирования восстановилась и лидирующее положение вновь заняли амфиподы и поденки.
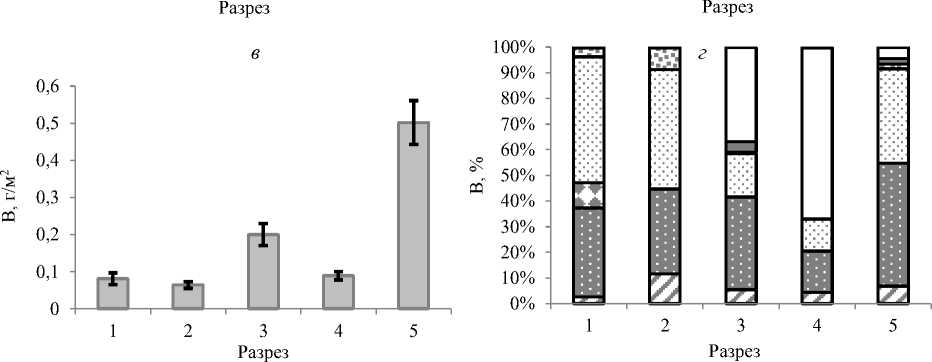
Расположение критических для макробентоса участков оценивали по распределению вдоль оси эстуария значений индекса Шеннона – Винера (рис. 5). В р. Горная отклонения от стандартного соотношения индексов характеризовали весь эстуарий реки, кроме устьевого разреза.
а
б
Z
50 0



100%
90%
80%
70%
60%
50%
40%
30%
20%
10%
0%


га







-
□ Oligochaeta
-
□ Amphipoda
в Isopoda
-
□ Ephemeroptera
-
□ Plecoptera
-
■ Trichoptera
-
□ Diptera
-
■ Прочие
Рис. 4. Изменение общей плотности ( а , б ) и биомассы ( в , г ) макробентоса вдоль эстуария р. Горная Fig. 4. Changes in the total density ( а , б ) and biomass ( в , г ) of macrobenthos along the estuary of the Gornaya River
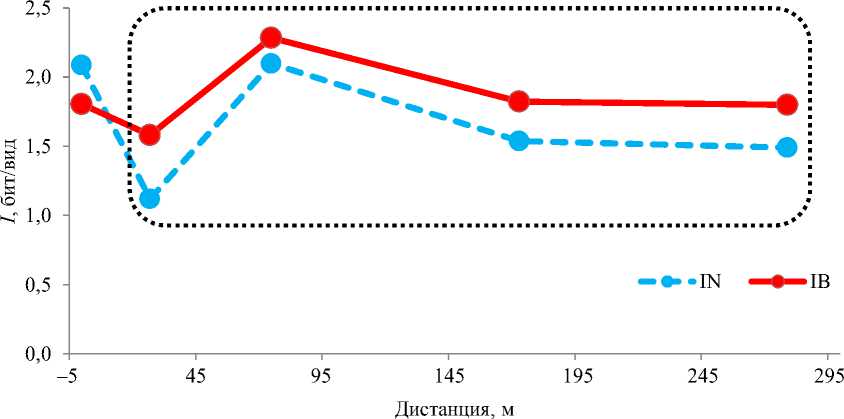
Рис. 5. Динамика индекса видового разнообразия по численности (IN) и биомассе (IB) макробентоса вдоль эстуария р. Горная
Fig. 5. Dynamics of the index of species diversity in terms of abundance (IN) and biomass (IB) of macrobenthos along the estuary of the Gornaya River
В распределении АВС-индекса (рис. 6) наблюдалось снижение на границах переходных зон. Положение нижней критической (граничной) точки на приустьевом створе закономерно показывало границу с морским прибрежьем. Критическая точка на разрезе 4 отмечала границу перехода эстуария к нижней ритрали реки.
20,0
18,0
16,0
14,0
12,0
10,0
8,0
6,0
4,0
2,0
0,0
–5
Дистанция, м
Рис. 6. Динамика АВС-индекса IABC макробентоса вдоль эстуария р. Горная
Fig. 6. Dynamics of the ABC-index IABC of macrobenthos along the estuary of the Gornaya River
Основные сообщества
На дендрограмме сходства станций по структуре макробентоса выделено шесть кластеров (рис. 7). Кластер "А" соответствовал донному сообществу с доминированием ракообразных Eogammarus kygi , доля доминанта от общей биомассы составляла 46 %. Диапазон глубин, на котором было выделено данное сообщество, варьировал от 0,1 до 0,87 м. Структура грунтов кластера включала мелкий песок с алевритом, мелкую и крупную гальку и булыжники (табл. 3).
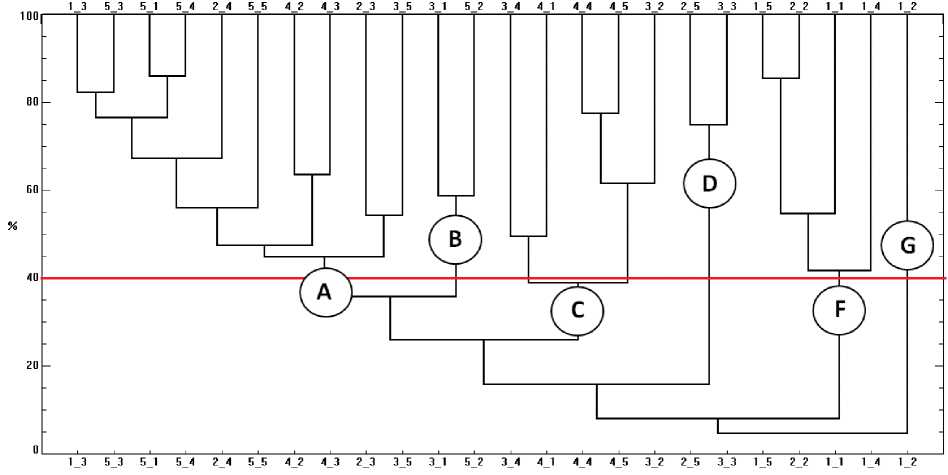
Рис. 7. Дендрограмма сходства станций макрозообентоса эстуария р. Горная
Fig. 7. Dendrogram of similarity of macrozoobenthos stations in the estuary of the Gornaya River
Таблица 3. Показатели обилия макробентоса донных сообществ эстуария р. Горная Table 3. Abundance of macrobenthos in benthic communities of the estuary of the Gornaya River
|
Сообщество (по доминанту) |
Eogammarus kygi |
Ephemerella aurivillii + Baetis sp. |
Tanytarsus |
Specaria josinae |
Baetis sp. |
Baetis (Асentrella) sibiricus |
|
|
Номер станций |
1_3, 5_3, 5_1, 5_4, 2_4, 5_5, 4_2, 4_3, 2_3, 3_5 |
3_1, 5_2 |
3_4, 4_1, 4_4, 4_5, 3_2 |
2_5, 3_3 |
1_5, 2_2, 1_1, 1_4 |
1_2 |
|
|
Глубина, м |
0,1–0,87 |
0,32–0,62 |
0,24–0,57 |
0,15–0,48 |
0,05–0,4 |
0,15 |
|
|
Тип грунта |
от мелкого песка с алевритом до крупной гальки |
от гальки с алевритом до крупной гальки |
алеврит, песок |
песок, алеврит, мелкая галька |
галька, песок |
галька |
|
|
S , psu |
0,1–12 |
0,1 |
0,1 |
0,1 |
0,1–12 |
6 |
|
|
S , видов |
34 |
15 |
21 |
4 |
8 |
2 |
|
|
N , экз./м2 |
226 ± 32 |
165 ± 16,6 |
253 ± 31 |
101 ± 11,5 |
29 ± 39 |
6 |
|
|
В , г/м2 |
0,280 ± 0,039 |
0,201 ± 0,020 |
0,130 ± 0,015 |
0,017 ± 0,002 |
0,51 ± 0,007 |
0,016 |
|
|
В доминанты , % |
46,2 |
51,4 |
56,5 |
88,8 |
68,9 |
96,6 |
|
|
QJ К 5 и m |
Oligochaeta |
5,5 |
6,0 |
5,6 |
88,8 |
1,9 |
3,4 |
|
Amphipoda |
46,2 |
10,7 |
6,6 |
0,0 |
5,5 |
0,0 |
|
|
Isopoda |
0,0 |
0,0 |
0,0 |
0,0 |
19,6 |
0,0 |
|
|
Ephemeroptera |
30,6 |
56,9 |
17,1 |
0,0 |
68,9 |
96,6 |
|
|
Plecoptera |
2,0 |
2,8 |
0,0 |
0,0 |
3,8 |
0,0 |
|
|
Trichoptera |
1,4 |
14,5 |
1,7 |
0,0 |
0,0 |
0,0 |
|
|
Diptera |
14,2 |
9,2 |
68,7 |
11,2 |
0,3 |
0,0 |
|
|
I , бит/вид |
N |
2,00 |
1,98 |
1,60 |
0,45 |
1,66 |
0,69 |
|
B |
2,04 |
2,04 |
2,11 |
0,45 |
0,98 |
0,15 |
|
|
I ABC |
4,1 |
17,6 |
–1,1 |
0,1 |
16,8 |
23,3 |
|
Следующий кластер "В" идентифицирован как сообщество поденок Ephemerella aurivillii и Baetis ( Baetis ) sp., в котором на долю доминирующих видов приходилось 51 % от общей биомассы. Состав грунтов сообщества был представлен галькой, алевритом, крупным песком и булыжниками на глубинах 0,32–0,62 м.
Кластер "С" объединял станции, локализованные на мелкодисперсных грунтах (алеврит, песок) с диапазоном глубин от 0,24 до 0,57 м. Доминировали в данном сообществе личинки и куколки двукрылых Tanytarsus sp., которые в сумме составляли 57 % от общей биомассы.
Следующий кластер "D" соответствовал сообществу малощетинковых червей Specaria josinae , которые формировали основную часть биомассы (89 %) на глубинах 0,15–0,48 м. Структура грунтов, на которых располагалось сообщество, включала песок, алеврит и мелкую гальку.
Сообщество личинок поденок Baetis sp. "F" было зафиксировано на глубинах 0,05–0,4 м преимущественно на галечных грунтах с небольшой примесью песка. Биомасса доминирующего вида составила 69 % от общей.
Последнее выделенное сообщество с доминированием личинок поденок Baetis (Асentrella) sibiricus – кластер "G" – располагалось на галечных грунтах на глубине 0,15 м. Они же слагали основу биомассы (96,6 %).
Распределение основных сообществ макробентоса в эстуарии р. Горная показано на рис. 8.
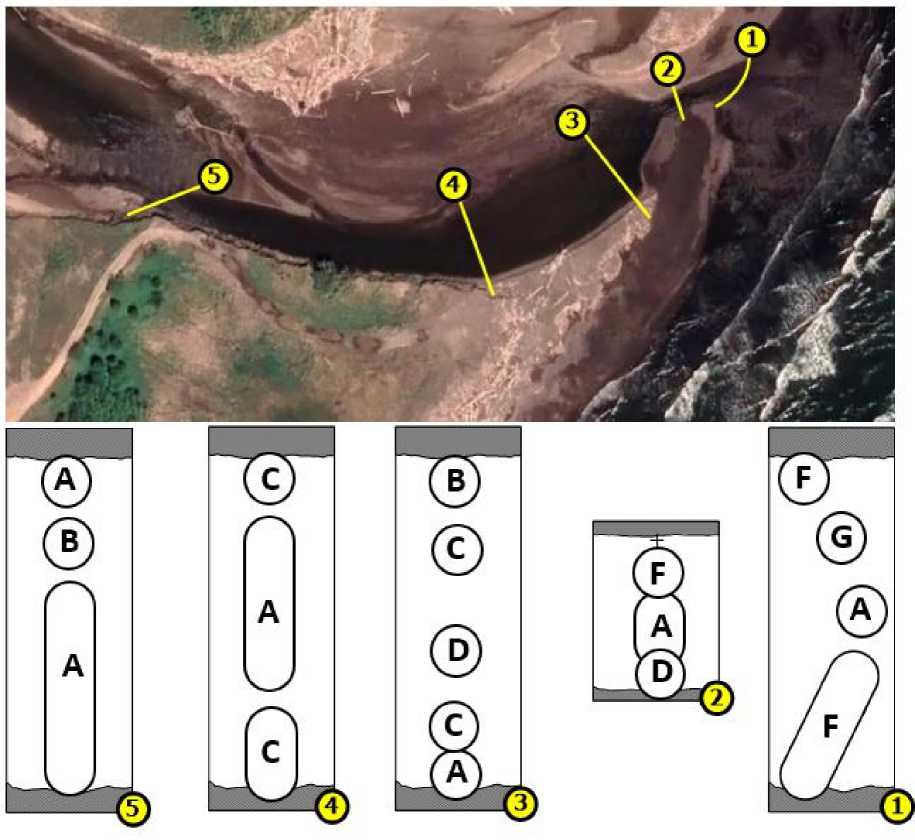
Рис. 8. Распределение основных сообществ макробентоса в эстуарии р. Горная, обозначения сообществ см. на рис. 7
Fig. 8. Distribution of main macrobenthos communities in the estuary of the Gornaya River, for community designations see Fig. 7
На ординационном плоте, показывающем сходство в распространении массовых видов бентоса, показателей обилия макробентоса в целом и известных абиотических факторов, большинство видов объединяются в одну группу (рис. 9), биомасса которых возрастает с уменьшением глубины.
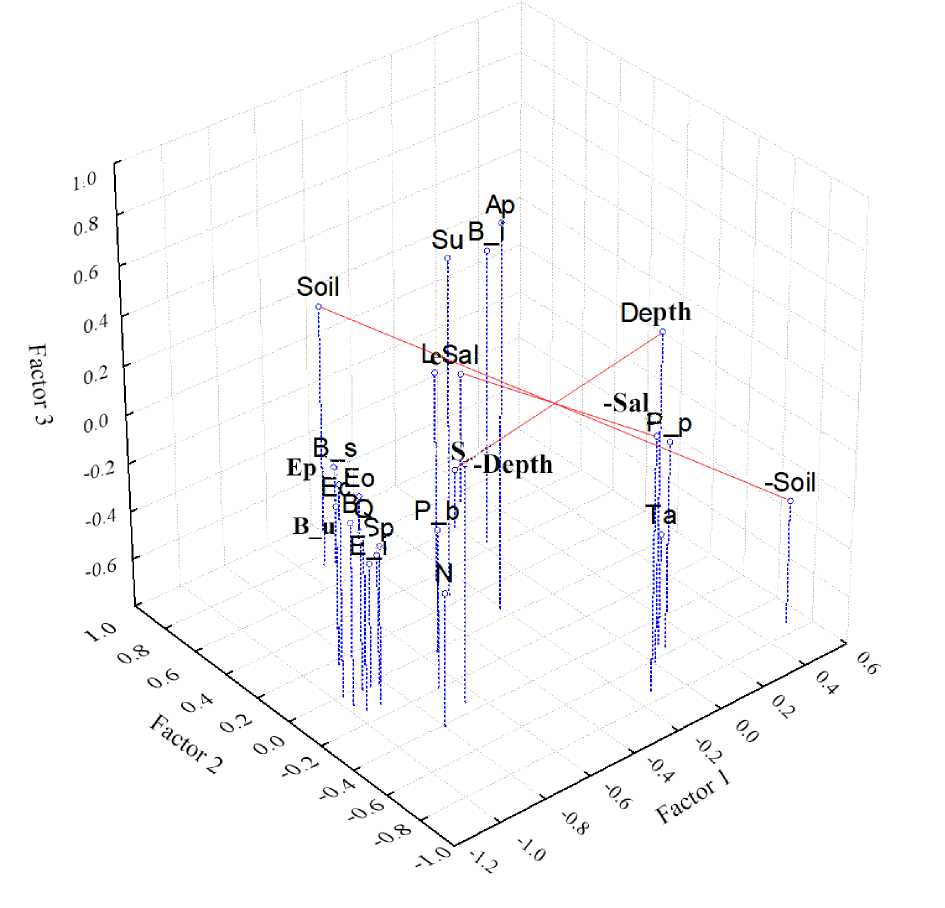
Рис. 9. Ординационный плот сходства массовых видов макробентоса, показателей обилия макробентоса и факторов среды; сокращения: Eo – Eogammarus kygi , P–b – Polypedilum bicrenatum , P–p – Polypedilum (Polypedilum) pedestre , Ta – Tanytarsus sp., B–i – Baetis sp., B–s – Baetis (Асentrella) sibiricus , Ec – Ecdyogymnurus aspersus , E–i – Epeorus sp., Ep – Ephemerella aurivillii , Le – Neoleptophlebia japonica , B–u – Вaetis ursinus , Sp – Specaria josinae , Su – Suwallia sp., Ap – Apatania sp., Sol – соленость воды, Temp – температура воды, Depth – глубина, Soil – тип грунта, S – количество видов, N – плотность, B – биомасса, Q – эквивалент энергетических затрат бентоса на дыхание Fig. 9. Ordination raft of similarity between mass species of macrobenthos, indicators of macrobenthos abundance and environmental factors; abbreviations: Eo – Eogammarus kygi , P–b – Polypedilum bicrenatum , P–p – Polypedilum (Polypedilum) pedestre , Ta – Tanytarsus sp., B–i – Baetis sp., B–s – Baetis (Асentrella) sibiricus , Ec – Ecdyogymnurus aspersus , E–i – Epeorus sp., Ep – Ephemerella aurivillii , Le – Neoleptophlebia japonica , B–u – Baetis ursinus , Sp – Specaria josinae , Su – Suwallia sp., Ap – Apatania sp., Sol – water salinity, Temp – water temperature, Depth – depth, Soil – soil type, S – number of species, N – density, B – biomass, Q – equivalent energy expenditure of benthos for respiration
Трофическая характеристика
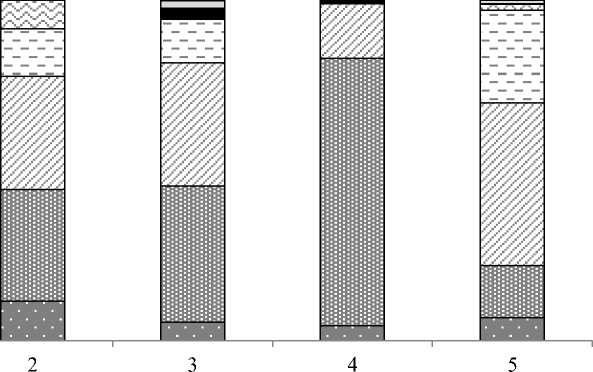
Из семи выявленных трофических групп основу биомассы макрозообентоса формировали всего три группы (рис. 10). В устье р. Горная "собирающие детритофаги", к которым относятся два вида поденок, четыре вида двукрылых и один вид изопод, создавали основу биомассы. Группа со смешанным типом питания "собирающие детритофаги + макроизмельчители + падальщики" была представлена одним видом амфипод Eogammarus kygi и имела второй по значимости вклад в общую биомассу. Третья по значимости группа "соскребатели" была представлена двумя видами поденок Ephemerella aurivillii и Ecdyogymnurus aspersus . Прочие группы – "грунтофаги", "микроизмельчатели" и "сестонофаги" вносили наименьший вклад в общую биомассу как на устьевом разрезе, так и по всему эстуарию в целом. Выше по течению основные группы, которые доминировали в устье реки, сохраняли свой вклад в общую биомассу, меняя пропорции. Хищники в эстуарии р. Горная были представлены только на двух разрезах (3, 4 разрезы) и вносили незначительный вклад в биомассу – от 0,8 до 2,8 %.
100 %

Разрез
90 %
80 %
70 %
60 %
50 %
40 %
30 %
20 %
10 %
0 %
□ De □ Dt □ Dt, Br, Sc □ Gr □ mBr ■ Pr □ Su □ Прочие
Рис. 10. Изменчивость биомассы ( В , г/м2) трофических групп макробентоса вдоль эстуария р. Горная Fig. 10. Variability of biomass ( B , g/m2) of trophic groups of macrobenthos along the estuary of the Gornaya River
Обсуждение результатов
Исследования гидрологических характеристик р. Горная в июле 2022 г. показали нетипичную для эстуарных зон ситуацию по солености. Здесь практически отсутствовал градиент солености как вертикальный, так и в направлении от устья к верхней части. В прилив морские соленые воды проникали только в приустьевую часть, в отлив из-за сильного речного стока эстуарий полностью заполнялся пресной водой. По нашим наблюдениям, заплеск морских вод в эстуарий р. Горная происходит только под действием нагонных и штормовых волн, что сопровождается резким, но кратковременным увеличением солености. Согласно гидролого-морфологической типизации ( Михайлов и др., 2009 ) данный эстуарий можно отнести к типу русловых эстуариев без устьевого расширения. Гидрологические процессы в таких эстуариях преимущественно находятся под влиянием речного стока, а влияние моря ограничивается сгонно-нагонными явлениями. Для данного случая, который нередко встречается в Сахалино-Курильском регионе, мы предлагаем ввести название "заплесковый эстуарий".
Несмотря на сравнительно малую протяженность эстуария, здесь был зарегистрирован внушительный видовой список донных беспозвоночных (44 вида). Для сравнения в эстуарии р. Мануй (протяженность 6,0 км) длина видового списка составляла 23 вида, в эстуарии р. Сусуя (8,5 км) – 57 видов. Большая часть представленных в эстуарии р. Горная беспозвоночных являлись типичными обитателями ритрали сахалинских рек ( Живоглядова и др., 2012; Лабай и др., 2015 ). Личинки поденок рода Baetis известны как обычная форма дрифта беспозвоночных ( Живоглядова и др., 2015 ) и, по-видимому, были снесены в эстуарий речным потоком.
Изменения в качественном и количественном составе донной фауны по продольному профилю определялись особенностями гидрологического режима створов. Структурирующая роль солености прослеживалась только на первых двух створах. Морские виды выше первого створа не проникали. На втором створе (27 м от устья), представляющим границу приливных вод, был отмечен минимум по числу видов. Здесь на одной из станций донные организмы вовсе отсутствовали. Третий створ (75 м от устья) характеризовался резкой сменой доминирующих групп. Четвертый и пятый створы (280 м от устья) представляли же типичную нижнюю ритраль с преобладанием сообщества Eogammarus kygi (Живоглядова и др., 2012; Лабай и др., 2015). Бокоплав Eogammarus kygi – реликт солоноватоводных плейстоценовых морей Японского и Охотского (Лабай, 2011), широко распространенный в низовьях сахалинских рек, где часто образует одноименное сообщество на участках, богатых детритом (Живоглядова и др., 2012).
Нестандартное соотношение индексов Шеннона – Винера характеризовало весь эстуарий реки, кроме устья. Это указывает на нестабильные условия эстуария, при которых формируются только молодые сообщества, представленные быстроразмножающимися мелкоразмерными видами.
Область критически низкой биомассы в эстуарии р. Горная находилась в приустьевой зоне, максимальная биомасса была отмечена на верхнем створе.
Трофическая структура является важным аспектом организации сообществ, позволяющим определить границы основных функциональных зон эстуариев. В эстуарии р. Горная при сравнительно небольшом числе групп – 7 (в рр. Мануй и Сусуя – 10) на всех разрезах наблюдалась схожая картина с доминированием одних и тех же трофических группировок и сменой вдоль оси эстуария только пропорциональных отношений друг к другу.
Гидрологический режим эстуариев имеет важное значение для молоди лососевых рыб в период ската, поскольку в условиях увеличения солености происходит осморегуляторная адаптация молоди к солености морской воды. Молодь горбуши в начальный период смолтификации более резистентна к условиям резко увеличивающейся солености воды, чем молодь кеты. Для молоди кеты при быстром переходе из пресной воды в морскую с соленостью около 30 ‰ гибель может достигать от 30 до 40 % ( Волобуев и др., 2011 ). Полученные данные по солености указывают на неблагоприятные условия в эстуарии р. Горная для выпуска кеты.
Заключение
На примере р. Горная рассмотрены особенности гидрологического режима и сообществ макрозообентоса эстуария руслового типа, в который в отсутствии штормовых заплесков проникновения морских вод практически не происходит. В прилив только в устье у дна отмечаются воды с соленостью более 12 psu, в отлив эстуарий полностью заполняется пресной водой. Влияние солености прослеживается на расстоянии не более 20–30 м от устья.
Показатели обилия зообентоса увеличиваются от устья эстуария к гравийно-галечному перекату, который является верхней границей эстуария. Нестабильные условия среды определяют существование в эстуарии молодых сообществ, представленных быстроразмножающимися мелкоразмерными видами.
Поскольку от условий эстуариев зависит успех адаптации молоди лососевых рыб в наиболее критический период онтогенеза, мониторинговые исследования гидрологии и кормовых ресурсов эстуарных зон лососевых рек о-ва Сахалин необходимо продолжить.
Авторы работы выражают свою искреннюю благодарность сотрудникам Сахалинского филиала Всероссийского научно-исследовательского института рыбного хозяйства и океанографии ("СахНИРО"), принимавшим участие в полевом этапе исследований и камеральной обработке проб.
