Массивность фаланг кисти в контексте происхождения неандертальцев Алтая

Автор: Медникова М.Б., Шуньков М.В., Маркин С.В.
Журнал: Археология, этнография и антропология Евразии @journal-aeae-ru
Рубрика: Антропология и палеогенетика
Статья в выпуске: 3 т.45, 2017 года.
Бесплатный доступ
Методом микротомографии исследована геометрия поперечного сечения медиальных фаланг 2-4-го лучей кисти пяти неандертальцев Европы и Азии (Ля Феррасси 1, Киик-Коба 1, Окладникова 2, 5, Чагырская 16-3-12) и пяти кроманьонцев (Костёнки 14, Тельмановская стоянка TII175 и TII173, Сунгирь 1, Абри Пато 26227). В каждом таксоне выявлен широкий размах индивидуальной изменчивости показателей внутренней массивности. У неандертальцев и наиболее массивные, и наиболее грацильные варианты костей кисти обнаружены на Алтае (в пещерах Окладникова и Чагырской соответственно), что подтверждает сделанный ранее вывод о высоком морфологическом разнообразии неандертальцев в этом регионе и наличии среди них как минимум двух морфотипов. Фаланги верхнепалеолитических сапиенсов Европы, в целом более толстостенные, чем у неандертальцев, в частности у мужчины Костёнки 14 наблюдается медуллярный стеноз. В изученной выборке на степень массивности фаланг не влияли половая и таксономическая принадлежность, а также биологический возраст и даже, по-видимому, биомеханический стресс. При этом гипермассивные варианты встречены нами у кроманьонцев и неандертальцев «гибридного» происхождения. Генетическими исследованиями, проведенными ранее, доказано, что Костёнки 14 относился к предковой для европейских верхнепалеолитических сапиенсов метапопуляции и унаследовал больший процент неандертальского генофонда; в формировании алтайских неандертальцев приняли участие не только денисовцы, но и ранние анатомически современные люди. В статье выдвинута гипотеза о связи гипермассивных вариантов строения трубчатых костей у населения среднего и верхнего палеолита Евразии с генетическим влиянием ранних анатомически современных людей.
Микротомография, медиальные фаланги кисти, неандертальцы, кроманьонцы
Короткий адрес: https://sciup.org/145145818
IDR: 145145818 | УДК: 572.77 | DOI: 10.17746/1563-0102.2017.45.3.126-135
Текст научной статьи Массивность фаланг кисти в контексте происхождения неандертальцев Алтая
Диафизарная структура трубчатых костей часто рассматривается сквозь призму характерной для современных и ископаемых людей двигательной активности. В последние десятилетия наиболее часто проводится биомеханический анализ человеческих останков из археологических раскопок, основанный на «инженерных» подходах оценки степени прочности элементов скелета [Ruff, 2000а, p. 71]. Форма кости определяется многими факторами. В раннем онтогенезе ведущая роль принадлежит преимущественно наследственным (генетическим) факторам. Негенетические факторы также могут оказывать генерализованное и локальное влияние. Системное изменение массивности скелета происходит в результате гормональной регуляции, пищевой недостаточности и патологий. Локальные гипертрофии отражают более ограниченные и специфические воздействия на костную ткань и иногда позволяют реконструировать поведенческие особенности.
Наблюдения за эпохальной динамикой внутренней массивности крупных трубчатых костей послужили основой для создания концепции ведущей роли механического воздействия при формообразовании [Ruff et al., 1993]. По мнению ее авторов, от раннего плейстоцена до современности происходит резкое снижение внутренней массивности бедренных и плечевых ко стей. По уровню внутренней массивности костей анатомически современные люди эпохи палеолита ближе к «архаическим», чем к ныне живущим. Но наиболее своеобразная картина характерна для эректусов: у них диафизарная массивность является результатом сужения медуллярного канала и расширения перио стального слоя. Хронологические тенденции изменения степени кортикализации крупных трубчатых ко стей создатели концепции объясняли постепенным снижением уровня механического воздействия на посткраниальный скелет. Они отмечали, что параллельно шел процесс увеличения головного мозга и развивалась культура, но при этом подчеркивали: когнитивные способности напрямую не связаны со степенью кортикализации скелета.
Археологические, палеогенетические и антропологические исследования последних лет способство- вали открытию в палеолитической ойкумене новых групп ископаемого человечества (таких как денисов-цы, алтайские неандертальцы, «хоббиты» с острова Флорес, Homo naledi). Останки некоторых из них представлены весьма фрагментарно, например, изолированными фалангами кисти. Важными задачами являются всестороннее описание и сравнительный анализ этих находок, в т.ч. с применением современных технологических приемов.
В фокусе внимания нашей работы – морфологические особенности мелких трубчатых ко стей кисти – медиальных фаланг – у представителей средне-и верхнепалеолитического населения Европы и Азии. При выборе объекта исследования мы исходили из того, что фаланги, связанные значимыми корреляционными связями с длинными костями, могут служить своеобразной «моделью» всего скелета. Это особенно важно при работе с изолированными и уникальными находками, представляющими ископаемых гомининов. Прочность костей верхней конечности по сравнению с нижней в меньшей степени зависит от общей массы тела [Ruff, 2000b, p. 285]. При этом именно ко сти кисти ископаемых людей, изготавливавших и активно использовавших каменные орудия, должны не сти следы структурных перестроек, которые связаны с рабочей гипертрофией костной ткани.
Целью данного исследования является изучение показателей внутренней массивности медиальных фаланг кисти представителей разных таксонов – неандертальцев и кроманьонцев и обсуждение спектра возможных факторов изменчивости.
Материал и методы
Сравнению были подвергнуты фаланги преимущественно 2-го или 4-го луча кисти ископаемых представителей рода Homo разной таксономической принадлежности – неандертальцев Европы и Азии, а также ранних и поздних верхнепалеолитических сапиенсов (табл. 1). Подчеркнем, что не все обследованные индивидуумы жили в перигляциальных условиях: в низкогорной зоне Алтая климат был мягче, чем в Европе, и ледника там не было [Природная среда…, 2003].
Таблица 1. Изученные материалы
|
№ образца |
Ископаемая находка |
Таксон |
Медиальная фаланга кисти |
Пол |
Возраст, лет |
|
1 |
Ля Феррасси 1 |
Европейский неандерталец |
4-й луч, правая |
Муж. |
40–55 |
|
2 |
Киик-Коба 1 |
» |
2-й луч, правая |
» |
40-49 |
|
3 |
Окладникова 2 |
Алтайский неандерталец |
4-й луч, левая |
» |
30–39 |
|
4 |
Окладникова 5 |
» |
2-й луч, правая |
Жен. |
25–39 |
|
5 |
Чагырская пещера 16-3-12 |
» |
То же |
» |
25–39 |
|
6 |
Костёнки 14 |
Верхнепалеолитический H. sapiens |
4-й луч, правая |
Муж. |
25–29 |
|
7 |
Костёнки 8 TII 175 |
» |
3-й луч (?) |
» |
25–39 |
|
8 |
Костёнки 8 TII 173 |
» |
4-й луч, правая |
» |
25–39 |
|
9 |
Сунгирь 1 |
» |
2-й луч, правая |
» |
35–45 |
|
10 |
Абри-Пато 26227 |
» |
То же |
Жен. |
20–29 |
Основу публикуемого исследования составляют наши наблюдения. Европейские неандертальцы представлены материалами из коллекции Музея естественной истории (Музея человека) в Париже и Музея антропологии и этнографии (Кунсткамеры) РАН в Санкт-Петербурге. Останки взрослого мужчины Ля Феррасси 1 были обнаружены в Дордони в 1909 г. [Capitan, Peyrony, 1912]. Останки взрослого неандертальца Киик-Коба 1, найденные в Крыму в 1925 г., были идентифицированы и подробнейшим образом описаны автором раскопок [Бонч-Осмоловский, 1941].
Группу алтайских неандертальцев характеризуют антропологические материалы из раскопок в пещерах Окладникова и Чагырской. Археологические исследования убедительно свидетельствуют о мустьероидном облике каменных индустрий этих пещерных объектов, получивших название «сиби-рячихинский вариант среднего палеолита Алтая» [Д еревянко, 2011; Деревянко, Маркин, Шуньков, 2013; Деревянко, Шуньков, Маркин, 2014]. Антропологические находки из пещеры Окладникова ранее по служили источником для выделения мтДНК неандертальского типа [Krause et al., 2007]. Морфологическое исследование палеоантропологических объектов подтвердило их принадлежно сть к неандертальцам [Viola, 2009; Медникова, 2011а, б; Medni-kova, 2011]. Фрагменты нижней челюсти [Viola et al., 2012] и локтевой кости [Медникова, 2013а] из раскопок Чагырской пещеры также соответствуют особенностям неандертальской анатомии.
Окладникова 2. Медиальная фаланга 3-го или 4-го лучей левой (?) кисти обнаружена в 1984 г. в пещере Окладникова, слое 3, кв. Г-4. Благодаря относительно небольшой длине и увеличенным поперечным размерам она попадает в поле изменчивости неандертальского населения [Медникова, 2011а, с. 55]. Если это фаланга 3-го пальца, то по длине, а также по ширине верхнего и нижнего эпифизов она соответствует мужчине, хотя по ширине и высоте тела тяготеет к неандертальским женщинам. Если это фаланга 4-го пальца, то она тем более мужская, поскольку крупнее соответствующих фаланг мужчин шанидар-ской группы [Там же, с. 71].
Окладникова 5 . Медиальная фаланга из слоя 3–1. Определена как женская правая фаланга 2-го луча [Там же, с. 67–68]. Как и предыдущая, характеризуется уплощенностью головки и относительно объемным диафизом [Там же, с. 70–71].
Чагырская 16-3-12 . Медиальная фаланга, обнаруженная в Чагырской пещере в 2012 г., также демонстрирует отдельные «архаические» особенности [Mednikova, Potrakhov, Bessonov, 2012]. Определена нами как женская правая фаланга 2-го луча.
Верхнепалеолитических сапиенсов в данном исследовании представляет один из наиболее ранних анатомически современных людей в Европе, о станки которого обнаружены на стоянке Костёнки 14 (Маркина Гора) [Marom et al., 2012; Nalawade-Chavan, McCullagh, Hedges, 2014]. Менее известны материалы с близлежащей стоянки Костёнки 8 (Тельманская стоянка), найденные экспедицией Ленинградского отделения Института археологии АН СССР под руководством А.Н. Рогачева предположительно в 1959 г. Более крупная, разрушенная в проксимальной части находка TII 175 идентифицирована как медиальная фаланга 3-го (?) луча кисти. Медиальная фаланга ТII 173 сохранилась целиком, она отно сится к 4-му лучу правой кисти [Медникова, Моисеев, Хартанович, 2016]. Останки другого раннего восточно-европейского кроманьонца – мужчины Сунгирь 1 – остаются в фокусе внимания исследователей на протяжении десятилетий. Так, в недавно опубликованных работах обсуждались главные особенности строения его кисти [Медникова, 2012; Trinkaus et al., 2014]. Медиальная фаланга Абри Пато из Музея естественной истории (№ 26227) позволяет составить представление о морфологических особенностях более поздних западноевропейских кроманьонцев периода протомадлена.
Медиальные фаланги всех перечисленных индивидуумов были подвергнуты микротомографии с целью исследования внутренней структуры объектов без их разрушения. Все находки, за исключением образцов Ля Феррасси 1, 2 и Абри Пато, которые изучались с помощью оборудования Музея естественной истории, были отсканированы на рентгеновском 3D-микроскопе Xradia Versa XRM-500 в лаборатории ООО «Системы микроскопии и анализа» (Москва). Ранее этот же прибор использовался для рентгеновской микроскопии фаланги девочки из Денисовой пещеры [Медникова и др., 2013].
В ходе исследования сравнивались виртуальные поперечные срезы в середине диафиза. Определялись стандартные показатели, описывающие геометрию поперечного сечения объектов (использована формула эллипса): площадь кортикального слоя (СА) – позволяет оценить устойчивость трубчатой ко сти к сжатию или растяжению; общая площадь сечения (ТА) – характеризует внешнюю массивно сть диафиза фаланги, площадь сечения медуллярного канала (МА); показатель кортикализации (% СА) – характеризует внутреннюю массивность диафиза, т.е. отно сительную толщину стенок. Полярный момент инерции (J) – характеризует устойчивость кости к сжатию и скручиванию. Применение по следнего показателя для реконструкции биомеханических нагрузок у человека вызывало критику [Lieberman et al., 2004], поэтому мы используем его с осторожностью. Кроме того, полярный момент инерции и длина тела (длина трубчатой кости) связаны более тесной корреляцией, чем длина и площадь кортикального слоя [Ruff, 2000b, p. 284]. Наиболее информативным представляется не зависящий от абсолютных размеров индекс – показатель относительного развития кортекса медиальных фаланг. Чтобы наши выводы не были искажены под влиянием фактора общих размеров трубчатых костей, мы остановимся на его значениях подробнее.
Результаты исследования
Среди обладателей изученных десяти фаланг кисти – мужчины и женщины, принадлежавшие к крупным и миниатюрным, внешне массивным и грацильным морфологическим вариантам. Самыми длинными фалангами с объемными диафизами характеризовались неандертальцы Ля Феррасси 1, Окладникова 2, кроманьонец ТII 173, самыми короткими и узкими – индивиды Костёнки 14 и Абри Пато 26227 (рис. 1).
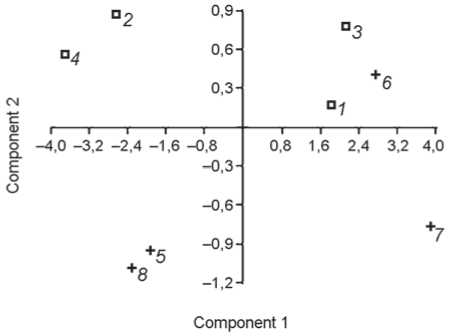
Рис. 1. Дифференциация медиальных фаланг по общим размерам методом главных компонент. Признаки – сочленовная длина, высота середины, ширина середины, окружность середины.
1 – Ля Феррасси 1; 2 – Киик-Коба 1; 3 – Окладникова 2; 4 – Чагырская 16-3-12; 5 – Костёнки 14; 6 – Костёнки 8 (ТII 173);
7 – Сунгирь 1; 8 – Абри Пато 26227.
При визуальном рассмотрении виртуальных поперечных срезов на уровне середины диафиза обращают на себя внимание различия в форме сечения фаланг (рис. 2). В подавляющем большинстве случаев она вытянутая (подовальная). Наиболее близки к овалу сечения фаланг неандертальцев Ля Феррасси 1 и Окладникова 2 и 5, а также (в меньшей степени) верхнепалеолитических сапиенсов с Тельмановской и Сунгирской стоянок. Это объясняется прежде всего тем, что развитие ладонных гребней фиксируется на уровне середины диафиза далеко не у всех индивидуумов: внешние структуры максимально выражены у мужчин Ля Феррасси 1 и Костёнки 14, значительно слабее гребни проявляются на срезах фаланг Киик-Коба 1, Чагырская, Костёнки 8 и сунгирца.
По величинам общей площади сечения наиболее объемные диафизы среди неандертальцев наблюдаются у мужчин из грота Ля Феррасси во Франции и из пещеры Окладникова на Алтае. Среди кроманьонцев нашей выборки общая площадь сечения уменьшается в ряду Костёнки 8 (обе фаланги), Сунгирь, Костён-ки 14, Абри Пато.
Среди форм с самыми узкими диафизами Абри Пато 26227, Костёнки 14, Окладникова 5. Первый и третий образцы представляют женщин, относящихся к разным таксонам, второй – согласно данным генетического анализа, мужчину.
При рассмотрении показателя массивности стенок обращает на себя внимание высокий диапазон индивидуальной изменчивости, характерный и для неандертальцев, и для кроманьонцев (рис. 3, табл. 2).
У неандертальских мужчин из Европы и Сибири различия в уровне кортикализации фаланг не столь
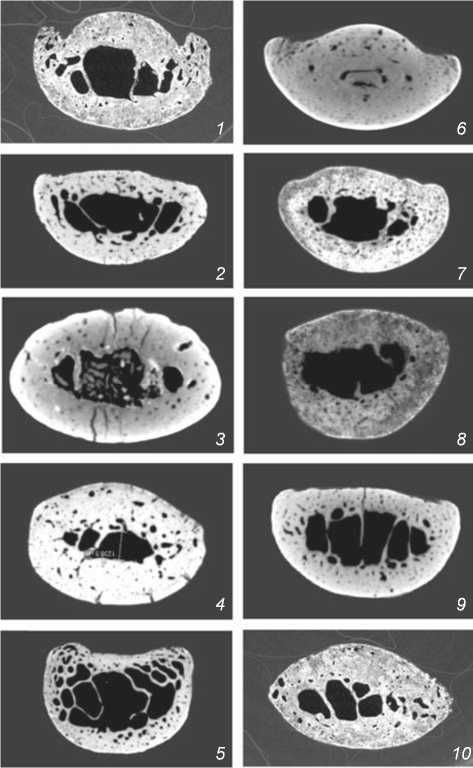
Рис. 2. Поперечные срезы медиальных фаланг кисти на уровне середины диафиза у неандертальцев ( 1-5 ) и кроманьонцев ( 6-10 ). Ладонная поверхность сверху. Нумерация ископаемых гомининов соответствует таковой в табл.1.
значительны по сравнению с представительницами этого таксона, обитавшими на Алтае в пещерах Окладникова и Чагырская. Данные именно по этим женщинам образуют границы изменчивости признака для неандертальцев: гипермассивный вариант в пещере Окладникова и гиперграцильный в Чагырской.
По этому признаку кроманьонцы также весьма вариативны, хотя уровень расхождений между наиболее толстостенной фалангой Костёнки 14 и самой тонкостенной фалангой Сунгирь 1 меньше, чем у неандертальцев.
В целом, вопреки ожиданиям, внутренняя массивность медиальных фаланг у большинства изученных нами кроманьонцев выше, чем у большинства неандертальцев. К тому же, самый тонкостенный диафиз выявлен у неандертальцев, у индивида из Чагырской пещеры.
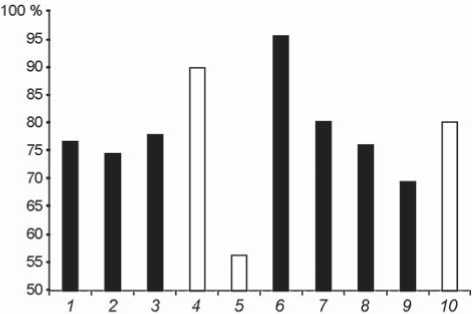
Рис. 3. Распределение значений показателя кортикали-зации медиальных фаланг кисти по полу. Нумерация ископаемых гомининов соответствует таковой в табл. 1. Черный цвет - мужчины, белый - женщины.
Таблица 2. Сравнительная характеристика степени внутренней массивности медиальных фаланг кисти на уровне середины диафиза у ископаемых Homo разной таксономической принадлежности
|
№ образца |
СА |
ТА |
МА |
%СA |
J |
|
1 |
41,48 |
54,17 |
12,69 |
76,57 |
507,52 |
|
2 |
28,96 |
38,93 |
9,97 |
74,39 |
255,19 |
|
3 |
35,36 |
45,41 |
10,05 |
77,88 |
330,65 |
|
4 |
33,88 |
37,7 |
3,82 |
89,87 |
231,58 |
|
5 |
22,92 |
40,70 |
17,78 |
56,32 |
231,71 |
|
6 |
34,33 |
35,93 |
1,61 |
95,6 |
2,47 |
|
7 |
38,17 |
47,62 |
9,45 |
80,15 |
373,22 |
|
8 |
34,53 |
45,45 |
10,92 |
75,97 |
333,52 |
|
9 |
26,37 |
37,89 |
11,52 |
69,61 |
230,82 |
|
10 |
26,47 |
33,02 |
6,55 |
80,17 |
168,46 |
Таким образом, по предварительной оценке на степень внутренней массивности мелких трубчатых костей ископаемых гомининов не влияют: половая и таксономическая принадлежность, а также биологический возраст. Это вынуждает нас рассмотреть полученные данные и возможные интерпретации более подробно.
Обсуждение
Внутренняя массивность трубчатых костей как функция биологического возраста. Толщина стенок трубчатых костей зависит от биологического возраста, т.е. от индивидуального состояния организма. В процессе онтогенеза толщина стенок сначала увеличивается, затем после стабильного периода зрелости происходят постдефинитивные изменения, связанные с постепенной потерей костного вещества и истончением кортекса. Развивается остеопороз.
Немаловажно, что благодаря хорошей сохранности останков принадлежность неандертальцев Ля Феррас-си 1, Киик-Коба 1 и кроманьонцев Сунгирь 1, Абри Пато к определенной возрастной когорте подтверждается по другим элементам скелета. Так, среди изученных нами представителей среднего и верхнего палеолита старшим является «классический» неандерталец Ля Феррасси 1. Он демонстрирует многочисленные проявления дегенеративно-дистрофических изменений в разных отделах скелета, облитерацию черепных швов. Его биологический возраст оценен в диапазоне 40–55 лет [Fennell, Trinkaus, 1997, р. 987]. Однако микротомография его медиальной фаланги не выявила костную резорбцию, связанную с остеопорозом, ни в дорзальной, ни в вентральной стенках диафиза, которые весьма массивны. Зоны интенсивных костных перестроек, наблюдаемые на виртуальном поперечном срезе фаланги, примыкают к ладонным гребням, и очевидно но сят функциональный характер, отражая натруженность мышц-сгибателей пальцев (см. рис. 2, 1 ).
Среди кроманьонцев старшим является мужчина Сунгирь 1, но он моложе неандертальца Ля Феррас-си 1. По первоначальной оценке Г.Ф. Дебеца [1967, с. 160], состояние черепных швов Сунгирь 1 соответствовало возрасту 55–65 лет. В.В. Бунак [1973, с. 3] определял возраст этого индивидуума как «позднюю зрелость». По разным системам признаков были исключены старческие дегенеративные изменения [Бу-жилова, Козловская, Медникова, 2000, с. 56]. Проведенное недавно наиболее дифференцированное рассмотрение возрастных маркеров позволило установить, что сунгирец скончался в промежутке от 35 до 45 лет [Trinkaus et al., 2014, p. 76]. Новые данные гистологического исследования его костной ткани целиком соответствуют этому заключению [Добровольская, Медникова, 2015].
Таким образом, в нашей выборке даже старшие по возрасту индивидуумы не обнаруживают признаков далеко зашедшего остеопороза или, наоборот, максимальной (функциональной) гипертрофии, т.е. фактор биологического возраста не оказывает влияния на степень массивности изучаемых фаланг.
Функция физических нагрузок. В 1892 г. немецкий анатом Юлиус Вольф предположил, что форма кости является результатом перемещения костных элементов в направлении функционального давления, его отражением является увеличение или снижение массы кости [Wolff, 1892]. Возможность применения «зако- на Вольфа» для определения уровня физических нагрузок вызвала дискуссию. С учетом того, что на процесс увеличения/снижения массы кости влияют еще и возраст, болезни, гормональный статус и наследственность [Ruff, Holt, Trinkaus, 2006, p. 485], сегодня предлагается применять термин «функциональная адаптация кости».
Развитие боковых гребней на ладонной поверхности фаланг кисти интерпретируется как несомненное проявление тренированности мышц-сгибателей пальцев. Наши данные свидетельствуют о разнообразной выраженности этого признака у людей верхнего и среднего палеолита. Можно предположить, что с возрастом гребни акцентированы сильнее. Действительно, Ля Феррасси 1 демонстрирует сильную гипертрофию гребней на уровне середины диафиза. По этому признаку к нему максимально близок молодой мужчина Костёнки 14. Если рассматривать данную особенность с позиций биомеханики, то следует сделать вывод: Костёнки 14 трудился упорнее сунгирца, у которого за долгие годы физических усилий не накопилось подобной суммы ответов костной ткани в виде рабочей гипертрофии. Это, безусловно, парадоксальный результат. Впрочем, такой формальный подход представляется неудачным, поскольку он не учитывает индивидуальных вариаций в местах прикрепления мышц-сгибателей пальцев. Например, у неандертальца из Киик-Кобы ладонные гребни есть, они сильно развиты, но сформированы проксимальнее уровня середины диафиза.
Несомненно, использование и изготовление каменных орудий труда оказывало особое биомеханическое воздействие на кисть палеолитического человека. Среди многочисленных научных работ по этой проблематике, вышедших в последние 150 лет, обращают на себя внимание недавние публикации, в которых приводится набор морфологических признаков, благоприятствовавших наиболее эффективному изготовлению и применению скребков и рубил [Marzke, 2013, p. 3]. М. Марцке задает вопрос, эволюционировала ли человеческая кисть, адаптируясь к изготовлению и применению каменных орудий. Сегодня, по мнению специалистов, нельзя утверждать, что кисть го-мининов эволюционировала путем естественного отбора только в результате адаптации к использованию орудий труда. Отдельные особенности человеческой кисти формировались в разные периоды в соответствии с собирательской стратегией выживания ранних гоми-нинов. Вместе с тем очевидно, что кисть ископаемого человека испытывала сильное и продолжительное воздействие, связанное с трудовой деятельностью.
Эксперименты с участием современных добровольцев позволили проследить последствия использования каменных артефактов. Например, оценивалась активность 17 мышц, управляющих движениями кисти и пальцев, при использовании олдованских орудий [Marzke et al., 1998]. Было установлено, что у ранних Homo максимальные силовые нагрузки испытывали 1-й и 5-й лучи.
Эксплуатация большинства палеолитических орудий предполагала использование двух техник удержания [Marzke, Shackley, 1986]. При применении первой – охват орудия ладонью сочетался с движениями пальцев, второй – орудие с силой удерживалось большим пальцем и кончиками 2-го и 3-го лучей кисти. Массивность кисти неандертальца была обусловлена функцией силового контроля определенного положения пальцев и постоянной манипуляцией орудиями. Неандертальцам, возможно, лучше, чем сапиенсам, удавался силовой поперечный зажим орудия; на это указывает гипертрофия широтных размеров фаланг, особенно дистальных [Churchill, 2001; Niewoehner et al., 2003].
Даже если учитывать всю сумму культурных и технологических различий населения эпох среднего и верхнего палеолита, то следует констатировать, что любые физические нагрузки неизбежно вели к рабочей гипертрофии мелких трубчатых костей кисти. Некоторые верхнепалеолитические люди (например, Сунгирь 1) при общей сапиентности в этой связи имели отдельные «неандерталоидные» черты: расширение ногтевых фаланг, мощные гребни на ладонной поверхности проксимальной фаланги, свидетельствующие об интенсивности биомеханического воздействия на кисть правой руки [Медникова, 2012].
Другим фактором, определявшим формирование кисти, был ландшафт, типичный для проживания многих популяций Homo в эпоху как среднего, так и верхнего палеолита. Экспериментальные данные по скалолазанию могут оказаться важными для понимания особенностей, связанных с двигательной активностью ископаемых людей, которые часто тяготели к гористой или холмистой местности. Структурные изменения кисти и отдельных пальцев обнаружены у современных спортсменов-скалолазов [Sylvester, Christensen, Kramer, 2006]. Установлено, что скалолазание оказывает интенсивное и долговременное механическое воздействие на ко сти кисти. Происходит перестройка метакарпалий и фаланг, что проявляется в утолщении кортекса. По сравнению с контрольной группой у скалолазов увеличена площадь поперечного сечения и вторичный момент площади. У них больше общая ширина костей кисти за счет добавочных субперио стальных отложений, а не увеличения пространства костномозгового канала. Прочность костей кисти коррелирует с современными стилями скалолазания и т.н. карабканьем по крупным валунам, требующими максимальной атлетической подготовки, в ходе которой происходят наиболее сильные изменения кисти.
Обследованные нами неандертальцы жили в горных условиях (скалы Дордони, Крыма, Алтая) и могли испытывать соответствующие им физические нагрузки. Кроманьонцев Костёнок и Сунгиря окружал ландшафт Восточно-Европейской равнины, где скалолазанье – значительно менее распространенный тип физической активности.
Как можно видеть, гипотеза исключительного биомеханического влияния на внутреннюю массивность фаланг кисти применительно к нашим наблюдениям не выглядит исчерпывающей.
Массивность костей кисти ископаемых гомини-нов среднего и верхнего палеолита в свете данных об их происхождении. Благодаря генетическим исследованиям определена древность дивергенции групп ископаемого человечества и установлены их генетические контакты. В изучаемой выборке представлены три группы населения среднего и верхнего палеолита: классические неандертальцы Европы, неандертальцы Алтая и анатомически современные сапиенсы.
Рассмотрим полученные результаты в свете имеющихся данных об их происхождении.
«Классический» неандерталец Ля Феррасси 1 (и, по-видимому, неандерталец Киик-Коба 1) демонстрирует особенности населения «доконтактной» с кроманьонцами эпохи.
Алтайские неандертальцы генетически составляют особую группу. Южная Сибирь входит в зону взаимодействия денисовцев и неандертальцев; об их генетических контактах красноречиво говорят результаты расшифровки высококачественных геномов обитателей Денисовой пещеры, принадлежавших к этим таксонам [Krause et al., 2010; Prüfer et al., 2014].
Генетически определенные останки неандертальцев происходят из трех алтайских пещер. Если суммировать информацию по всем посткраниальным находкам с Алтая, то с точки зрения морфологии местные неандертальцы демонстрируют высокую индивидуальную изменчивость, в которой прослеживаются определенные закономерно сти [Mednikova, 2014, 2015]. Проксимальная фаланга стопы и дистальная фаланга кисти из Денисовой пещеры [Медникова, 2011б, 2013б], трубчатые кости разных индивидуумов из пещеры Окладникова [Медникова, 2011а] выделяются, даже на общенеандертальском фоне, очень толстыми стенками. Посткраниальные ко сти из Чагырской пещеры принадлежали более типичным представителям неандертальской анатомии (напр.: [Медникова, 2013a]), без экстаординарной внутренней массивности.
Результаты изучения медиальных фаланг кисти в нашей работе полностью подтверждают ранее сделанные выводы о дифференциации неандертальцев Южной Сибири как минимум на два морфологических варианта. Высказывалось предположение, что эти варианты связаны с разными волнами миграций H. neanderthalensis на Алтай, а также с генетическими контактами неандертальцев с представителями других таксонов, например, с денисовцами [Mednikova, 2015]. Вместе с тем с учетом территориальной близости указанных пещер мы не можем исключить генетических контактов и между группами проживавших в них людей. В любом случае широкий размах изменчивости является фенотипическим отражением сложной истории происхождения алтайских неандертальцев.
Термин «алтайские неандертальцы» появился в процессе изучения полного генома обитательницы Денисовой пещеры [Prüfer et al., 2014], по нашим данным, одной из носительниц гипермассивного варианта внутреннего строения [Медникова, 2011б]. Логичным было бы допустить, что утолщение стенок трубчатых костей у представителей этой популяции определялось не только биомеханическим воздействием, но и наследственными факторами, например, генетическим влиянием реликтовых денисовцев. Однако результаты нашей работы позволили установить, что некоторые кроманьонцы в Европе также характеризовались более утолщенными стенками мелких трубчатых костей по сравнению с неандертальцами «классическими», а также населявшими Чагырскую пещеру. Речь идет прежде всего об обитателях верхнепалеолитических стоянок в долине среднего Дона, среди которых интерес вызывает мужчина Костён-ки 14, фаланги которого отличаются особой массивностью. Поскольку скелет этого человека сохранился очень хорошо (в частности, полностью представлены обе его кисти), были сделаны выводы о том, что по массивности его скелетная система превосходила таковую у современных жителей Евразии [Медникова, Моисеев, Хартанович, 2016]. Радиологическими методами у индивидуума Костёнки 14 были выявлены множественные медуллярные стенозы (вытеснение костного мозга компактой) в дистальных и медиальных фалангах обеих кистей, а также в проксимальных фалангах левой кисти (см. рис. 2, 6 ). В рамках дифференциальной диагностики наиболее вероятным представляется вариант наследственного гипотиреоидо-за. Крайняя степень медуллярного стеноза трубчатых костей кисти среди верхнеплейстоценовых ископаемых форм до этого была определена только у неандертальца из Денисовой пещеры (на ногтевой фаланге) [Медникова, 2013б].
Генетическое исследование показало принадлеж-но сть индивидуума Костёнки 14 к метапопуляции, предковой для многих современных людей и для верхнепалеолитических сапиенсов, включая и образец из Абри Пато. Геном обитателя долины среднего Дона содержит более протяженные участки неандертальской ДНК [Seguin-Orlando et al., 2014].
Таким образом, утяжеление скелета у представителей среднего и верхнего палеолита Евразии за счет утолщения стенок трубчатых костей обнаружено нами в двух гибридных группах. Одна рассматривается как прародитель современных верхнепалеолитических и отчасти современных европейцев, среди предков которого были неандертальцы, другую составляют неандертальцы, контактировавшие с денисовцами (образцы из пещер Денисова и Окладникова).
Впрочем, новое исследование генома алтайских неандертальцев выявило неожиданные особенности их «гибридного происхождения» – контакты с сапиен-сами [Kuhlwilm et al., 2016]. До появления указанной публикации считалось, что метисация неандертальцев и анатомически современных людей произошла за пределами Африки 65–47 тыс. л.н. В работе М. Кал-вилма с соавторами геномы алтайского неандертальца и денисовца были рассмотрены вместе с последовательностями 21 хромосомы двух неандертальцев из Испании и Хорватии. Установлено, что популяция, рано дивергировавшая от остального африканского («сапиентного») населения, внесла генетический вклад в генофонд предков алтайских неандертальцев ок. 100 тыс. л.н. У денисовцев и двух европейских неандертальцев «сапиентное» генетическое наследие не выявлено. С учетом этого сделан вывод о том, что гибридизация предков неандертальцев Алтая с ранними современными людьми происходила, скорее всего, на Ближнем Востоке, т.е. она удревнилась на десятки тысяч лет.
Итак, по скольку внутренняя гипермассивность трубчатых костей Костёнок 14 определенно не является наследием денисовцев, связующим звеном между ним (или шире – его метапопуляцией) и алтайскими неандертальцами можно считать ранних сапиенсов. В этом случае гипертрофия стенок трубчатых костей была обусловлена наследственным фактором. В основе явления – миграция (в случае Костёнки 14) или генетическое влияние (в случае алтайских неандертальцев) представителей современной анатомии из тропических широт.
Заключение
Приступая к исследованию, мы рассчитывали найти подтверждение предположения о повышенной внутренней массивности трубчатых костей кисти у неандертальцев по сравнению с верхнепалеолитическими европейскими сапиенсами. Эти ожидания опирались на представления о бóльшей приспособленности неандертальцев к силовому зажиму орудий труда и, в конечном счете, на концепцию ведущей роли биомеханического воздействия на утолщение костных стенок. Однако наши данные свидетельствуют о том, что фаланги кроманьонского населения, как правило, были более толстостенными. Вопреки ожиданиям внутренняя массивность не зависит от пола и возраста индивидуумов. Видимо, она не зависит и от формального таксона. Например, алтайские неандертальцы из пещеры Окладникова (если учитывать результаты исследования других мелких трубчатых костей, то и из Денисовой пещеры) также демонстрируют внутреннюю гипермассивность.
Согласно палеогенетическим исследованиям, алтайские неандертальцы Денисовой пещеры (и, по нашему предположению, пещеры Окладникова), отчасти являются потомками сапиенсов, живших более 100 тыс. л.н. Поэтому в ответе на вопрос, кто из плейстоценовых Homo стал «источником» формирования у них исключительно тяжеловесных вариантов строения скелетной системы, можно указать ранних анатомически современных тропидов.
Данное исследование подтвердило ранее высказанное предположение о морфологической неоднородности неандертальских обитателей Алтая, отражающей, скорее всего, сложную генетическую историю этой группы, которая вобрала в себя наследие двух других таксонов. В перспективе анализ морфологии алтайских палеоантропов может быть использован для выявления иных следов воздействия денисовцев или ранних анатомически современных людей.
Исследование выполнено за счет гранта Российского научного фонда (проект № 14-50-00036). Авторы выражают благодарность А. Фроману и Музею естественной истории (Музей человека) в Париже за предоставленную возможность микротомографирования фаланг Ля Феррасси 1 и Абри Пато 26227, В.И. Хартановичу (Музей этнографии и антропологии (Кунсткамера) РАН в Санкт-Петербурге) за возможность исследовать антропологические находки Костёнки 14 и 8.
Список литературы Массивность фаланг кисти в контексте происхождения неандертальцев Алтая
- Бонч-Осмоловский Г.А. Кисть ископаемого человека из грота Киик-Коба//Палеолит Крыма. -М.: Л.: Изд-во АН СССР, 1941. -Вып. II. -170 с.
- Бужилова А.П., Козловская М.В., Медникова М.Б. Сунгирь 1//Homo sungirensis: Верхнепалеолитический человек: экологические и эволюционные аспекты исследования. -М.: Науч. мир, 2000. -C. 54-56.
- Бунак В.В. Ископаемый человек из стоянки Сунгирь и его место среди других ископаемых позднего палеолита//IX Междунар. конгресс антропол. и этнограф. наук. -Чикаго, 1973. -С. 117.
- Дебец Г.Ф. Скелет верхнепалеолитического человека из погребения на сунгирской стоянке//СА. -1967. -№ 3. -С. 160-164.
- Деревянко А.П. Верхний палеолит в Африке и Евразии и формирование человека современного анатомического типа. -Новосибирск: Изд-во ИАЭТ СО РАН, 2011. -560 с.
