Математическая модель эволюционно стабильной стратегии для популяции в условиях конкуренции на неоднородном ареале

Автор: Зеленчук П.А.
Журнал: Российский журнал биомеханики @journal-biomech
Статья в выпуске: 2 т.29, 2025 года.
Бесплатный доступ
На основе уравнений диффузии – реакции рассмотрена система двух конкурирующих за единый ресурс родственных популяций. Дана простая математическая формулировка исследуемой задачи, учитывающая диффузионные процессы и локальное взаимодействие между членами изучаемого биологического сообщества в условиях неоднородности среды обитания. С использованием концепции идеального свободного распределения (ИСР) для одной из двух популяций построена модель эволюционно стабильной стратегии (ЭСС), приводящей к ее долговременному устойчивому существованию. Проанализированы стационарные решения системы и условия их устойчивости на одномерном ареале с неравномерным распределением ресурса. Сформулированы принципы (правила поведения), выполнение которых обеспечивает реализацию ЭСС выбранной популяции. Для численного решения начальнокраевой задачи применяется модифицированный интегроинтерполяционный метод с дискретизацией на основе смещенных сеток. На основе реализованного в среде MATLAB комплекса программ SPECIES21 проведена серия вычислительных экспериментов, устанавливающая степень влияния трех ключевых факторов (начальных распределений популяций, коэффициентов диффузии и конкурентного взаимодействия) на поведение системы.
Конкуренция, уравнения диффузии–реакции, идеальное свободное распределение, эволюционно стабильная стратегия
Короткий адрес: https://sciup.org/146283131
IDR: 146283131 | УДК: 519.63 | DOI: 10.15593/RZhBiomeh/2025.2.03
Mathematical model of evolutionary stable strategy for population under conditions of competition in a heterogeneous habitat
Based on the diffusion-reaction equations, a system of two related populations competing for a single resource is considered. A simple mathematical formulation of the problem under study is given, taking into account diffusion processes and local interactions between members of the stud-ied biological community in conditions of a heterogeneous habitat. Using the concept of ideal free distribution (IFD), a model of an evolutionarily stable strategy (ESS) is constructed for one of the two populations, leading to its long-term sustainable existence. Stationary solutions of the system and the conditions for their stability in a one-dimensional area with an uneven distribution of the resource are analyzed. The principles (rules of conduct) have been formulated, the achievement of which ensures the implementation of the ESS of the selected population. A modified integro-interpolation method with discretization based on shifted grids is used for the numerical solution of the initial-boundary value problem. Based on the SPECIES-21 software package implemented in the MATLAB environment, a series of computational experiments was conducted to establish the degree of influence of three key factors (initial population distributions, diffusion coefficients, and competitive interaction) on the behavior of the system.
Текст научной статьи Математическая модель эволюционно стабильной стратегии для популяции в условиях конкуренции на неоднородном ареале
Конкуренция, наряду с хищничеством, – один из наиболее распространненых типов взаимодействия между живыми организмами на всех уровнях, от вирусов [1] и бактерий [2; 3] до растений и животных [4; 5]. Недавние исследования [5; 6] показали, что это явление присуще даже клеткам в эпителиальных и других тканях высших организмов[7; 8]. Клеточная конкуренция участвует в различных физиологических и патологических процессах и является важнейшим гомеостатическим механизмом, необходимым для эмбрионального развития, профилактики заболеваний и старения [9–11].
Основная причина конкуренции – это соперничество за ресурсы (вода, свет, пища, территория и т. д.), количество которых, как правило, ограничено. Конкурентные отношения составляют неотъемлемую часть борьбы за существование в экологическом сообществе, определяемом как совокупность популяций по крайней мере двух различных видов, взаимодействующих в пределах определенной (географической) области [12; 13].
0000-0001-6598-8521


Эта статья доступна в соответствии с условиями лицензии Creative Commons Attribution-NonCommercial 4.0 International
License (CC BY-NC 4.0)
This work is licensed under a Creative Commons Attribution-NonCommercial 4.0 International License (CC BY-NC 4.0)
Внутривидовая и межвидовая конкуренция оказывает сильное влияние на динамику конкретной популяции, видовое разнообразие [14] и структуру всего экологического сообщества [15]. Существуют различные механизмы и формы конкурентных отношений [5]. На самом общем уровне можно выделить прямую конкуренцию, при которой две популяции одного или разных видов вступают в открытую конфронтацию, реализующуюся во всяческих способах взаимного угнетения (схватки, аллелопатия и пр.), и косвенную конкуренцию, когда одна из популяций опосредованно влияет на другую, ухудшая условия ее существования (опережение, монополизация пространства и пр.). В любом случае конкуренция всегда снижает общую приспособленность обоих участников этого процесса [16].
Согласно принципу конкурентного исключения (закон Гаузе) [17], менее приспособленный вид в долгосрочной перспективе обречен либо на вымирание, либо на вытеснение в другую экологическую нишу [18]. Этот принцип предсказывается теоретическими рассуждениями, математическими моделями и лабораторными экспериментами, однако почти не наблюдается в естественных экосистемах [19]. Считается, что одним из основных механизмов, сохраняющим сосуществование популяций и объясняющим разнообразие видов в биологических сообществах, является компромис между конкуренцией и колонизацией [20]. Так, виды, которые больше преуспели в конкурентной борьбе, будут доминировать на конкретном участке ареала или в определенной экологической нише, приобретая более «специализированную» стратегию, в то время как виды, обладающие «универсальной» стратегией, будут активнее колоннизировать другие участки ареала или занимать новые экологические ниши [21].
Важным вопросом является то, какую стратегию выберет каждая из взаимодействующих популяций, и приведет ли она ее к успеху, т.е. будет ли эта стратегия устойчивой в эволюционном смысле [22]. Эволюционно стабильной стратегией (ЭСС) называется такая стратегия поведения, направленная на решение адаптационных задач, при которой популяция обеспечивает свое долговременное существование и не может быть вытеснена с ареала аналогичной популяцией с другой стратегией [23]. ЭСС относится к области теории игр: существует предположение, что индивидуальная приспособленность особей популяции может рассматриваться как аналог теоретико-игрового понятия полезности или выгоды, представляющей собой величину, измеряющую то, что игроки оптимизируют для достижения своих целей [24].
В неоднородной среде обитания интересным представляется также возникновение и эволюция пространственного распределения видов [25–27]. Одним из основных математических инструментов для анализа пространственно-временных процессов в биологических сообществах являются уравнения в частных производных, позволяющие описать процессы формирования устойчивых пространственных структур [28; 29]. В качестве одной из теоретических моделей реализации пространственного распределения популяции на неоднородном ареале, приводящей к ЭСС, является идеальное свободное распределение (ИСР) [30]. Согласно теоретической модели ИСР, все особи популяции размещаются пропорционально доступному на ареале ресурсу, имеют идеальное представление о его распределении и возможном изменении со временем и могут свободно перемещаться по ареалу с целью повысить свою индивидуальную приспособленность [31]. ИСР, изначально введенное для одного вида, рассматривалось для систем «хищник – жертва» [32–34] и нескольких видов, конкурирующих за общий ресурс [35]. В работах [36; 37] для системы «хищник – жертва» ИСР рассматривается как пропорциональность распределения обоих видов в стационарном или осциллирующем режиме. В статье [38] описан более общий случай ИСР-подобного распределения, когда обобщенному ресурсу пропорциональна только «жертва», а в распределении «хищника» возможны различные варианты.
В исследовании рассматривается математическая модель стратегии для одной из двух конкурирующих за общий ресурс родственных популяций на неоднородном ареале. Построение модели ведется на основе принципов ИСР с целью получения ЭСС для выбранной популяции.
Материалы и методы
Модель двух конкурирующих популяций
Рассмотрим для описания динамики численности двух конкурирующих за единый ресурс популяций V , V систему дифференциальных уравнений в частных производных (одномерная модель диффузии – реакции) [39; 40]
‘V- = D 1 — VV + a l V 1 p ( x )- b 11 V 1 2 - d t О x
b i VV ,
S
V = D 2 ^ VT + a 2 V 2 P ( x ) - b 22 V -d t dx
b 2 V 1 V 2 ,
где D, D2 — коэффициенты диффузии, a{, a2 — коэффициенты роста обеих популяций соответственно, а p ( x ) -неравномерно распределенный на ареале ресурс. Отметим, что функция ресурса нигде на ареале не обращается в нуль и имеет достаточное количество производных, что облегчает общий анализ системы. Коэффициенты b 1, b22 отвечают за конкуренцию между особями внутри каждой популяции, а b 2, b21 - за конкуренцию между популяциями. Переменная x изменяется на отрезке [ 0;1 ] , на концах которого задаются какие-либо граничные условия. Так же, система (1) дополняется начальными условиями распределения популяций на ареале.
Обилие коэффициентов затрудняет анализ модели.
Заменой
t = i, k , = Dl, k 2 = D a 1 a 1 a 1
,
a 1
a 2
S Г, = -U U1,V2 = -U U 2, b11 b22
Y = a 2 , e , = b 2 , ₽ 2 = b22
a 1 b 22 b 11
система (1) может быть сведена к виду
U = к
5 г 1
S
U = k ar •
d2U , 5 x 2
d 2 U 2
2 5 x 2
+ U 1 [ Р ( x ) - U 1 - e i Y U 2 ] ,
, (3)
+ Y U2 Р ( x )- — U1
_ Y
- U 2
.
Будем считать, что рассматриваемые популяции относятся к одному виду или к малоотличающимся родственным видам [41], так что их коэффициенты роста одинаковы ( a = a2 , Y = 1), а коэффициенты диффузии достаточно малы ( k , k2< 1) и подчиняются условию
k2 = е к , , ( 0 < £ < 1 ) . ( 4 )
Обозначая производную по времени точкой, а по координате штрихом и учитывая вышесказанное, получим
u 1 = kU 1' + u 1 [ p ( X ) - u 1 - в и 2 ] ,
U 2 = £ kU 2 + U 2 [ Р ( X ) - в 2 U 1 - U 2 ] ,
где PP P2 - коэффициенты межпопуляционной конкуренции в нормированных переменных.
Дополним систему (5) условиями периодичности
U 1 ( 0,т ) = U 1 ( 1,т ) , U 2 ( 0,т ) = U 2 ( 1,т ) ( 6 )
и начальными условиями
Ц ( x , 0 ) = U 0 , U 2 ( x , 0 ) = U 2 0 . ( 7 )
Постановка задачи
Цель настоящего исследования состоит в попытке построить эволюционно стабильную стратегию для популяции, находящейся в условиях конкуренции со стороны родственной популяции на неоднородном ареале. Суть такой стратегии заключается в том, чтобы выработать некоторое правило поведения (одно или несколько), обеспечивающее долговременное существование указанной популяции при воздействии неблагоприятных факторов, например, увеличения негативного влияния со стороны конкурента, скачки численности, вызванные
внешним влиянием, и т. д. Математически это должно выражаться в четком понимании того, какие параметры модели должны быть изменены, для того, чтобы выполнялись упомянутые выше правила, формирующие стратегию.
Исходя из начально-краевой задачи (5)-(7), выберем в качестве фокус-популяции, т. е. популяции носителя выстраиваемой эволюционно-стабильной стратегии популяцию U . Согласно приведенной математической модели, наша фокус-популяция обладает всего тремя возможными механизмами управления своей стратегией, заключающимися в изменении: 1) параметра диффузии е; 2) коэффициента конкурентного взаимодействия Р2 ; 3) начальных условий U 0 . Отметим, что величина популяции и ее распределение в начальный момент времени важны в основном при рассмотрении стартовой задачи, возникающей в случае инвазии или заселении вида на новую территорию (место обитания). Разнообразные сценарии развития пространственно-временной динамики системы (5)–(7), возникающие при вариации парметров указанных пунктов, будут рассмотрены ниже.
Методы решения
Для поиска и исследования устойчивости стационарных решений модели (5)–(7) в работе используются стандартные методы математического анализа, теории динамических систем на фазовой плоскости [42; 43] и асимптотического анализа дифференциальных уравнений с малым параметром при производной [44].
Для численного решения сформулированной начально-краевой задачи, основу которой составляет система дифференциальных уравнений в частных производных типа «диффузия - реакция», применяется модифицированный интегро-интерполяционный метод (метод баланса) с дискретизацией на основе смещенных сеток и метод прямых с интегрированием по схеме Рунге - Кутты четвертого порядка, детально описанный в работах [45; 46]. Для реализации метода в среде MATLA B разработан комплекс программ SPECIES -21, защищенный свидетельством о государственной регистрации программ для ЭВМ [47]. Данный комплекс позволяет проводить вычислительные эксперименты для нелинейных диффузионно-адвективных моделей, описывающих пространственно-временную динамику взаимодействующих популяций в условиях неоднородности среды обитания.
Результаты и обсуждение
Начнем анализ модели (5)–(7) в предположении, что коэффициент диффузии настолько мал, что им можно пренебречь ( k = 0). Тогда уравнения (5) могут быть записаны в виде
и 1 = U 1 [ p ( x ) - U 1 - e i U 2 ] ,
U 2 = U 2 [ P ( x ) - в 2 и 1 - U 2 ] .
Система (8) имеет тривиальное стационарное решение { U , U2 } = { 0,0 } и три стационарных решения, отвечающих ИСР
{ P ( x ),0 }
{ °, P ( x ) }
I 1(1 -ил
p ( x ),
1 -0 1
1 — 0 1 0 2
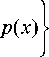
(9, а )
(9, б )
(9, в )
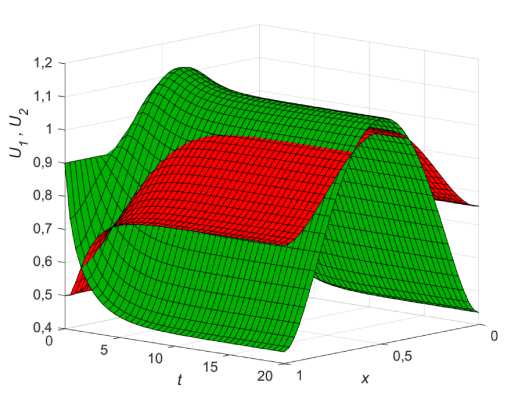
Выполнение условия (10, в ) приводит к стабильному сосуществованию обеих популяций на ареале с пространственным распределением, пропорционам ресурсу p ( x ) , причем та популяция, чей коэффициент конкурентного взаимодействия (Р;, z = 1,2 . ) меньше, будет доминировать по своей численности. Если, например, 02 < 01 < 1, то независимо от начальных условий, в системе (8) установится стационарное состояние (9, в ), при котором фокус-популяция будет располагаться выше конкурента, как показано на (рис. 1), где начальные условия U 0 ( x ) = 0,6, U 0 ( x ) = 0,3, а функция ресурса задается выражением:
Анализ стационарных решений (9) показывает, что их устойчивость определяется условиями:
P(x)
= + Sin nx .
в1 < 1, | в1 >1 , в1 < 1 , в2 > 1 , в2 < 1, в2 < 1 .
( 10, а )
( 10, б )
( 10, в )
Условие (10, а ) означает, что вторая популяция U2 уступает в конкурентной борьбе первой U и в конечном счёте полностью вытесняется ею с ареала. Поскольку U2 — это наша фокус-популяция, то такой вариант развития событий с точки зрения выстраивания эволюционно стабильной стратегии совершенно неприемлем.
Условие (10, б ) - полная противоположность условию (10, а ) и соответствует эволюционным задачам фо-кус-популяции - по возможности победить в конкурентной борьбе и сохранить свое существование в течение длительного временного промежутка.
Отметим, что даже в случае Р[ < Р2 < 1 стратегию фокус-популяции можно также считать эволюционно устойчивой, так как она позволяет сохранять U 2 ( x ) > 0 на ареале бесконечно долго, пусть даже и уступая U j ( x ) по численности. При Р[ = Р2 = 1 возникает очень интересная ситуация, приводящая к появлению целого семейства стационарных решений, описываемого теорией косиметрии [48; 49]. Однако в реальности такое совпадение маловероятно и здесь рассматриваться не будет. Условие же 0i,02 > 1 описано ниже.
Исследуем теперь поведение системы (5)–(7) при наличии диффузии и рассмотрим вопрос о ее влиянии на стационарные состояния (9). Будем искать решение возмущенной (малым параметром k ) системы (5) в виде:
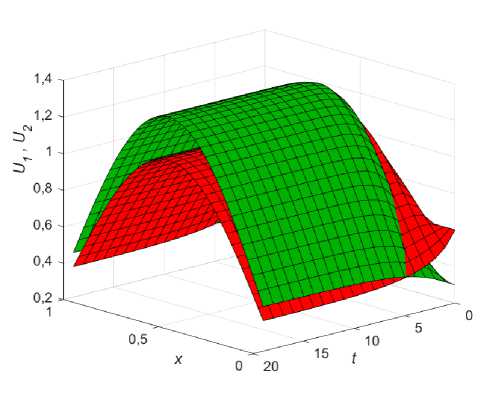
Рис. 1. Распределение фокус-популяции (зеленый) и популяции конкурента (красный) на ареале с неоднородным распределением ресурса p ( x ) для стационарного состояния (9, в ) при р 1 = 0,3; р 2 = 0,15
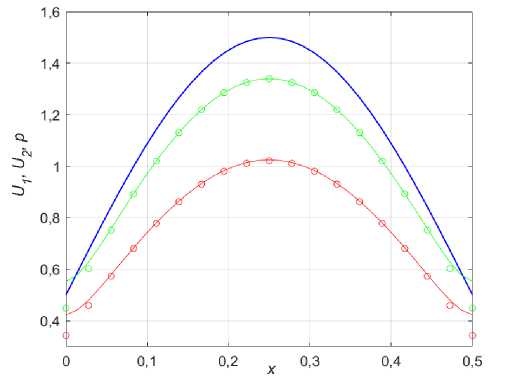
Рис. 2. Сравнение распределений U (красный) и U2 (зеленый) по отношению к ресурсу p (синий) рассчитанных численно (линии) и асимптотически (шарики) при k 1 = 0,001; £ = 1, Р 1 = 0,35; р 2 = 0,15
и 1 = U 10 + k i U 11 + kyU 12 +••• , <
_ U 2 = U 20 + k 1 U 21 + k\U 22 +• .
Подставляя (12) в (5) и ограничиваясь первыми двумя членами асимптотического разложения, получим приближенное решение:
|
U 1 1 — в 1 в 2 ( |
. p "( x ) p ( x )+ k 1 p ( x ) l |
1 -. 1 -P 1 |
£в 1 1 — в 2 . |
, 1 ( 13 ) |
|
U 2 = |
/ \ P ”( x ) - p ( x ) + k j 1 |
£ |
β 2 - |
. |
|
2 1 - в 1 в 2 |
1 p ( x ) |
. 1 - в 2 |
1 — в 1. |
] |
На рис. 2 изображены распределения популяций в сравнении с ресурсом при наличии малой ( к 1 = 0,001) диффузии. Сплошными линиями показано решение, полученное с помощью численного метода, а шариками -по формулам (13). Видно, что диффузия «растаскивает» исходное, пропорциональное p ( x ) решение (9 в ) по краям, уменьшая его максимум в центре ареала. По мере увеличения диффузии этот эффект становится все сильнее, правда результаты, полученные с помощью выражений (13) становятся менее достоверными, так как к перестает быть малым параметром. Наличие диффузии в системе (5)-(7) нарушает ИСР, но все же при к1<^1 стационарные решения такого рода могут быть названы ИСР-подобными [36-38].
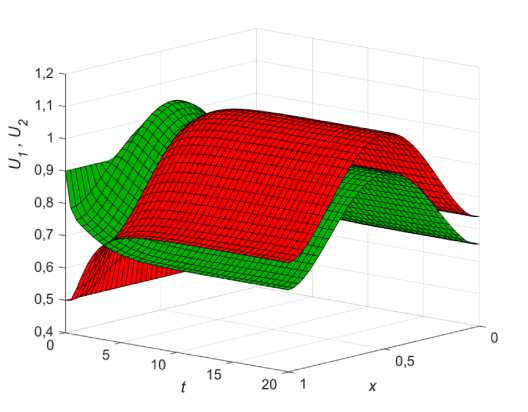
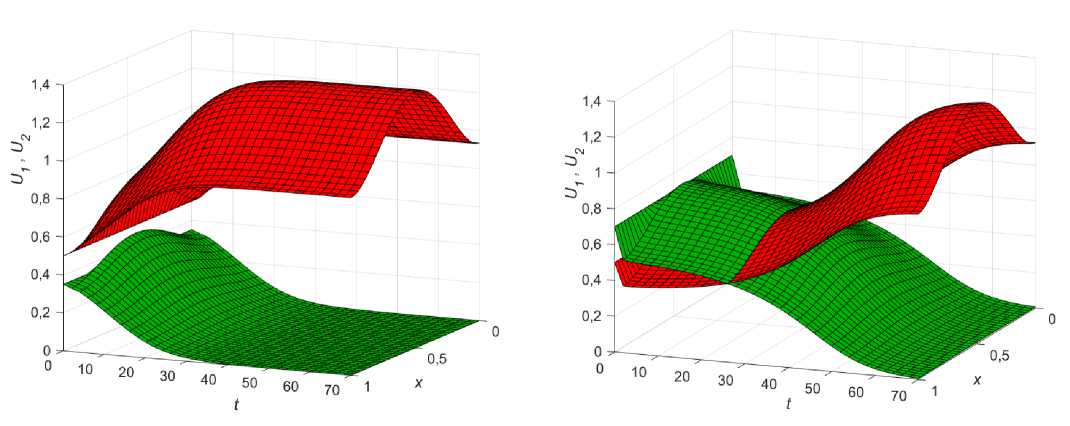
Общий анализ уравнений (13) показывает, что изменение параметра диффузии £ может быть использовано фокус-популяцией для улучшения своей приспособленности на ареале, так, например, при в1 < в2 < 1 устанавливается равновесие, при котором численность первой популяции больше, чем численность второй, однако если последняя уменьшит свою диффузию путем изменения ε , то с течением времени сможет добиться локального перевеса в месте максимума ресурса, сохраняя при этом общее равновесие системы (рис. 3).
Заметим, что, несмотря на разницу стартовых условий ( U 0 > U 0 ), снижение коэффициента конкурентного взаимодействия в2 < в 1 и сильные изменения в коэффициенте диффузии ( £ = 0,1... 0,01), фокус-популяция не сможет полность вытеснить конкурента с ареала до тех пор, пока будет выполнено условие (10 в ). Это связано с тем, что собственные значения линеаризованной системы (5)-(7) лежат строго в левой комплексной полуплоскости и не могут перейти в правую при условии в12 < 1 [50]. Потеря устойчивости положения равновесия (9 в) может произойти лишь при нарушении условия (10 в ).
Рассмотрим поведение системы (5)–(7) в случае, когда в 12 > 1. Как уже было сказано выше, стационарное состояние, при котором на ареале сосуществуют оба вида, станет неустойчивым, и малейшее возмущение приведет со временем к гибели одного из видов. Можно сказать, что конкурентная борьба сильно обостряется, теперь уже не может быть мирного сожительства с разными численностями Ux ( x ) и U 2 ( x ) , выживает та популяция, которая сможет эффективнее снизить конкурентное влияние другой. В этом смысле для того, чтобы фокус-популяции сохранить свою стратегию эволюционно стабильной, необходимо просто побеждать в кон-куретной борьбе (1 < в2 < в 1 ). Однако возможны и другие варианты.
а
Рис. 3. Влияние коэффициента диффузии фокус-популяции на решение (9, в ) при в 1 = 0,3; в 2 = 0,38; к 1 = 0,03;
б
U 1 0 = 0,5; U 0 = 0,9 : а - диффузия одинакова (£ = 1); б - диффузия фокус-популяции меньше ( £ = 0,1)
Предположим, что фокус-популяция проигрывает в конкурентной борьбе (1 < в1 < вг ), но У нее остается возможность варьировать начальные условия и коэффициент диффузии. Покажем на конкретных вычислительных примерах, что оба этих параметра могут привести к смене стационарного решения.
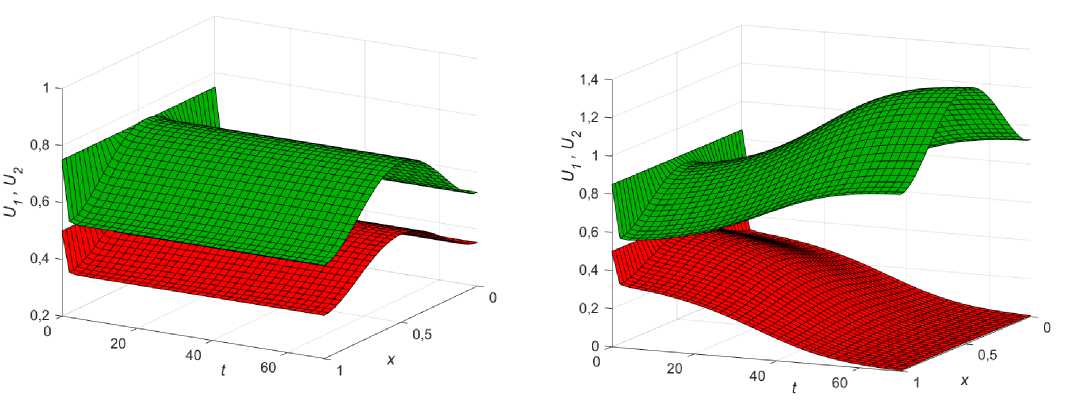
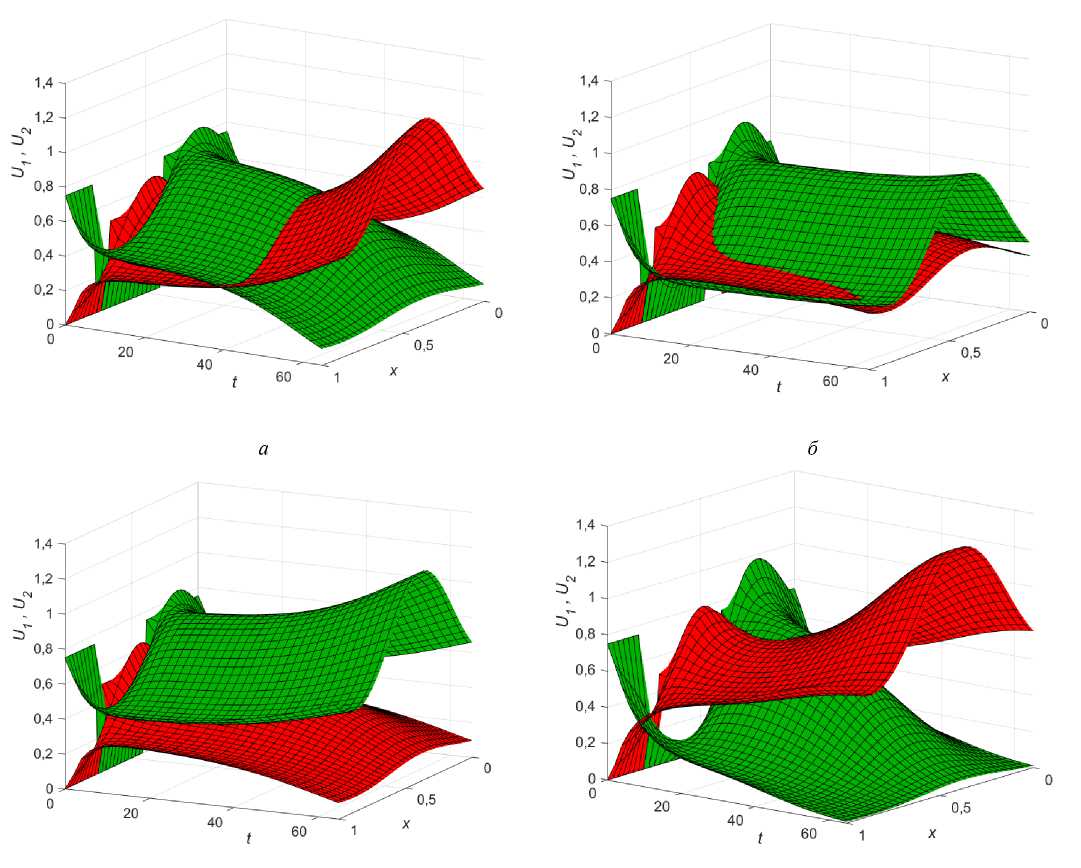
Пусть изначально коэффициенты диффузии равны, е = 1, и Р[ = 1,15 < Р2 = 1,2 , тогда если начальное распределение U 0 > U 0 , то первая популяция однозначно выигрывает конкурентную борьбу у второй (рис. 4, а ).
Данное стационарное решение сохраняется даже при небольшом начальном перевесе фокус-популяции (рис. 4, б ), но начиная с некоторой разницы в численности, может установиться стационарное состояние, соответствующее сосуществованию видов (рис. 4, в ), которое будет неустойчивым по отношению к возмущению других параметров. Наконец, при дальнейшем увеличении разницы в начальных распределениях фокус-популяция быстро вытесняет конкурента со всего ареала (рис. 4, г ).
Вопрос о нахождении точного соотношения между начальными распределениями популяций задающего условие смены стационарных решений интересен, но выходит далеко за рамки данной работы.
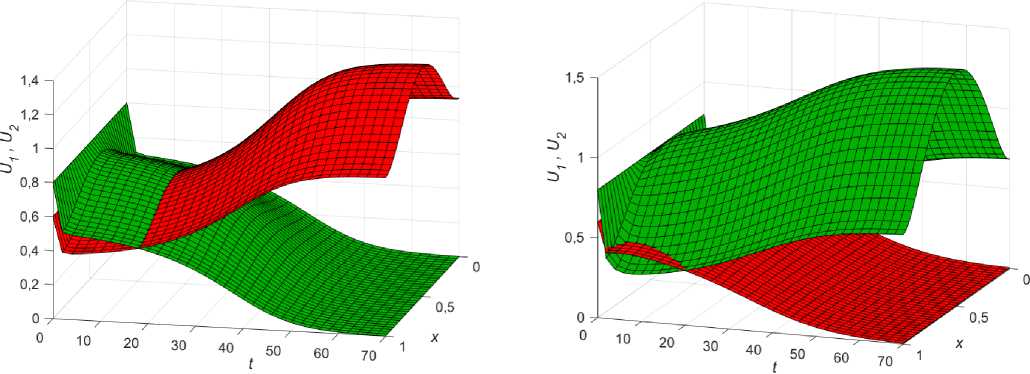
Известно [42; 50], что разность в коэффициентах диффузии может также привести к смене равновесия (рис. 5). В таких случаях говорят о так называемой неустойчивости Тьюринга [51].
Естественно, что получить перевес в конкурентной борьбе при 1 < в1 < вг фокус-популяция может, комби- нируя изменения упомянутых выше двух параметров (U0,е ). Особенно интересен этот вопрос в случае инвазии одного из видов или освоения (заселения) новых территорий, когда начальные распределения по ареалу не постоянны (U02 = f (x)).
Рассмотрим вначале случай, когда p (x) = 1 + “ sin f 2nx
π
—
а б
в г
Рис. 4. Влияние начальных условий на решение системы (5)-(7) при в1 = 1,1; вг = 1,15 : а - U 0 = 0,5; U 0 = 0,35; б - U 0 = 0,5; U 0 = 0,7 ; в - U 0 = 0,5; U 0 = 0,75 ; г - U 0 = 0,5; U 0 = 0,85
а
б
Рис. 5. Смена стационарного решения (вх = 1,1; вг = 1,15; к = 0,03; U 0 = 0,6; U 0
= 0,8 :) а - е = 1; б - е = 0,1
в г
Рис. 6. Смена стационарного решения (Р[ = 1,1; Р2 = 1,15; к = 0,01, U 0 , U 0 определены из (15)-(16):
а - е = 1; б - е = 0,72 ; в - е = 1, но U 0 = 0,75 при x е
9;9
; г - е = 0,2
а начальные численности популяций заданы на постоянном уровне, но на всем интервале:
0,5
U0 (x ) = 1
0,
( 1 7
при x el -;-
_ ( 1 7
при x gl —
0,
при x e
U0 (x Н
0,75
при x g
9;6
9;6
Как показано на рис. 6, а , несмотря на интегральное превосходство в начальный момент времени, фокус-популяция тем не менее проигрывает в долгосрочной перспективе при равных коэффициентах диффузии и 1 < в1 < вг ■ С уменьшением коэффициента диффузии
(е < 1) ситуация начинает меняться, и при определенном пороговом значении (е < 0,73) фокус-популяция переигрывает конкурента (рис. 6, б ). Этого же результата можно добиться и увеличением начальной численности U 0 ( x ) , например сужая отрезок с нулевым количеством фокус-популяции с [ 4/9;5/6 ] до [ 4/9;7/9 ] , как это видно на (рис. 6, в ). Отметим, что увеличение U 0 ( x ) повышает пороговое значение s и позволяет второй популяции добиваться превосходства при меньшем изменении коэффициента диффузии. Однако чрезмерное снижение коэффициента диффузии может привести к обратному эффекту, например, если е = 0,2 , то превосходства на ареале добивается первая популяция (рис. 6, г ). Это связано с законом распределения ресурса на ареале, сильная диффузия приводит к тому, что фокус-популяция «проскакивает» максимум и не может до достичь критического перевеса в локальных точках до-
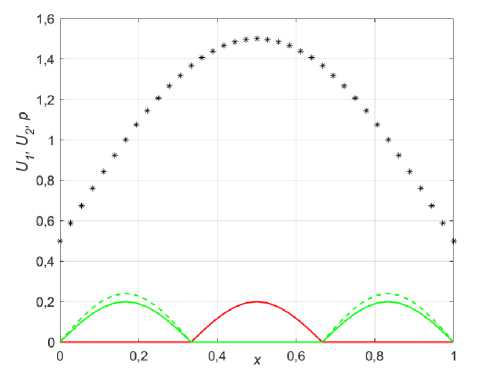
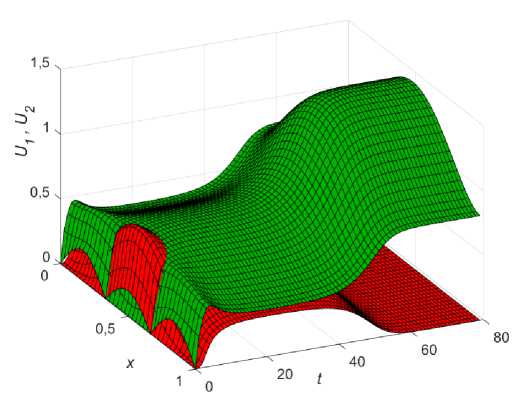
Рис. 7. Начальное распределение и сценарии конкурентного взаимодействия популяций при колонизации р! = 1,35; р2 = 1,4; к = 0,01; p ( x ) = 1/2 + sinn x : а - сравнение распределения ресурса (звездочки) с U 0 (красный), исходным U 0 (сплошной зеленый) и увеличенным U 0 (зеленый пунктир); б - сценарий взаимодействия при е = 1; в - параметр е = 1, но U 0 возросло; г - диффузия фокус-популяции сильно меньше (е = 1,2)
г
стичь критического перевеса в локальных точках пространства, в результате чего и проигрывает в долгосрочной перспективе в целом.
Рассмотрим теперь случай колонизации (заселения новых территорий), когда оба вида прибывают (высаживаются естественным путем) в неосвоенную область ареала или новую среду обитания. Пусть функция ресурса в этой области задана выражением (11). Предположим также, что первая популяция имеет прямое конкурентное преимущество (1 < в1 < Р2 ) и согласно концепции ИСР высаживается на новом ареале в том месте, где расположен максимум ресурса. Несмотря на интегральное численное превосходство фокус-популяции (площадь под зеленой кривой в два раза больше чем под красной, (рис. 7, а )), при равной диффузии она проигрывает конкуренту (рис. 7, б). Однако уже небольшое увеличение начальной численности ( U 02 - U 0j ® 10 %) приводит к тому, что с течением времени вторая популяция вытесняет первую (рис. 7, в ). Ускорения этого процесса фокус-популяция может добиться, успользуя увеличение своей диффузии. Действительно, поскольку особи второй популяции распологаются по обе стороны от максимума ресурса, то рост диффузии приведет к ускоренному их проникновению в центр ареала, создавая критическое количество, необходимое для победы в конкурентной борьбе (рис. 7, г ).
Интересна также ситуация, когда при колонизации оба вида не имеют представления о функции ресурса в новой среде обитания и распределяются равномерно в шахматном порядке (рис. 8, сплошные кривые).
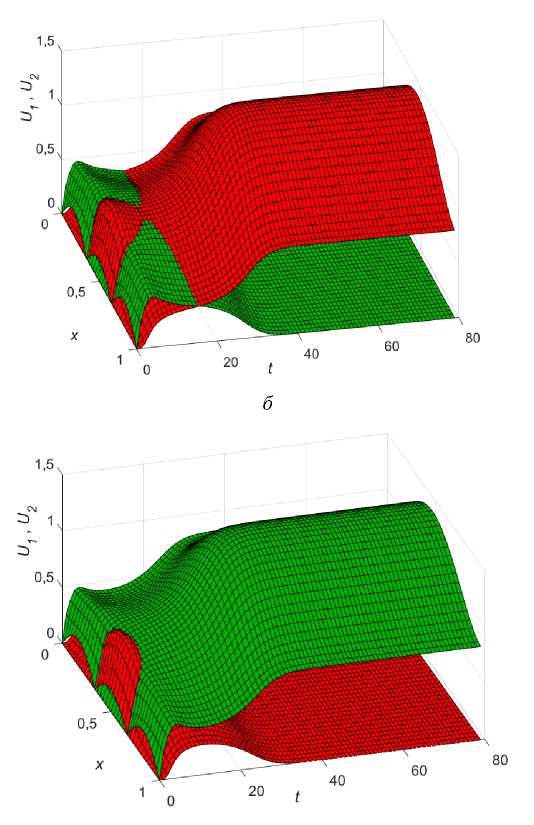
Пусть, как и прежде, 1 < в1 < Р2 и функция ресурса имеет вид (11). Будем для сравнения наблюдать за плотностями популяций в двух точках ареала с координатами x = 0,25 (сплошные линии на рис. 9) и x = 0,45 (пунктирные линии на рис. 9). При равных численностях популяций победу в конкурентной борьбе одержит U , как
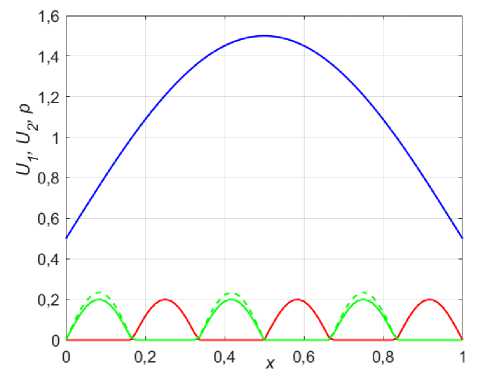
Рис. 8. Начальное распределение Ux (красный)
и U2 (зеленый) по отношению к ресурсу p (синий) при U 0 = U 0 (сплошные) и U 0 > U 0 (пунктир)
это видно на (рис. 9, а). В этом случае ни увеличение, ни уменьшение диффузии (даже очень маленькое) не может помочь фокус-популяции переиграть конкурента. Например, уменьшение k 2 (от е = 1 до е = 0,1) приводит к динамике, изображенной на (рис. 9, б) . Добиться превосходства U можно, лишь увеличив начальное распределение U 0 > U 0 (зеленый пунктир на рис. 8). Динамика такого сценария изображена на (рис. 9, в ). При численном перевесе на старте фокус-популяция может позволить себе изменение коэффециента диффузии и в меньшую и в большую сторону, однако до определенного предела. Например, при увеличении k уже при е = 1,1 (рис. 9, г ) верх одерживает популяция Ux . При уменьшении k ситуация чуть лучше, однако по достижении е = 0,15 U опять выигрывает.
Таким образом, при колонизации в условиях неопределенности и равномерном стартовом распределении популяций (при 1 < в1 < Р2 ) может сложиться ситуация (нельзя исключать иные распределения ресурса), когда ЭСС для фокус-популяции состоит только в численном превосходстве начального распределения.
Заключение
Предложенная в статье модель ЭСС для популяции, находящейся в условиях конкуренции на неоднородном ареале, базируется на системе дифференциальных уравнений в частных производных, относящихся к классу уравнений диффузии - реакции. Модель построена с учетом принципа ИСР, позволяющего получать максимальную приспособленность особей популяции к месту обитания. Сама стратегия сводится к определенному набору правил поведения, обеспечивающих долговременное существование в условиях конкурентной борьбы с родственной популяцией за неравномерно распределенный ресурс.
Анализ модели (5) показывает, что система может иметь три стационарных решения (9), которые в общей сложности зависят от трех факторов: нормированных коэффициентов межпопуляционной конкуренции Ри Р2, диффузии (к, е) и начальных условий (U[0, U2 ). Долговременное существование фокус-популяции подразумевает несколько вариантов, худший из которых состоит в простом выживании при численном доминировании конкурента, что соответствует одному из вариантов решения (9 в), а лучший - в полной победе над другой популяцией (конкурент погиб или вытеснен за пределы ареала), что соответствует решению (9 б). Приведем ниже основные правила, придерживаясь которых фокус-популяция будет обладать ЭСС.
В отсутствии диффузии (к = 0) при Р12 < 1 фокус-популяция будет всегда закрепляться на ареале при лю- бых начальных условиях (7), а ее распределение по ареалу будет соответствовать ИСР, как, впрочем, и распределение популяции конкурента. Если при этом необходимо, чтобы ее численность была выше численности конкурента, то должно выполняться неравенство в2 < в1 ■
Наличие малой диффузии приводит к отклонению стационарного решения от ИСР и может быть использовано фокус-популяцией для улучшения своей приспособленности на ареале, так, при в1 < вг < 1, уменьшая параметр S , она может добиться локального перевеса в месте максимума ресурса, сохраняя при этом общее равновесие системы см. (рис. 3, б).
В случае обострения конкурентной борьбы, когда в12 > 1 и сосуществование двух популяций на ареале становится неустойчивым, фокус-популяции для сохранения ЭСС необходимо либо полностью побеждать в конкуретной борьбе (1 < Р2 < в1), либо при 1 < в1 < в2 варьировать свой коэффициент диффузии и начальные условия в зависимости от внешних обстоятельств. Так, при равных стартовых условиях необходимо снижать диффузию, уменьшая параметр S . В остальных случаях фокус-популяции следует добиваться численного превосходства в начальных условиях и такого изменения параметра s (в большую или меньшую сторрону), которое приводит к быстрейшему притоку особей в места с максимальным значением ресурса. Такие сценарии хорошо проиллюстрированы (рис. 6 и 7).
В данной работе построение ЭСС для популяции в условиях конкуренции за общий ресурс на неоднородном ареале велось на основе одномерной модели диффузии - реакции в предположении, что популяция-конкурент является родственной ( a [ = a 2, y = 1) , а коэффициенты диффузии достаточно малы ( k , k 2< 1). Изменение любого из этих условий приводит к усложнению задачи и требует дополнительных исследований. Для будущих задач особо интересны случаи конкурентного взаимодействия с нелинейными коэффициентами диффузии [52; 53].
б
в г
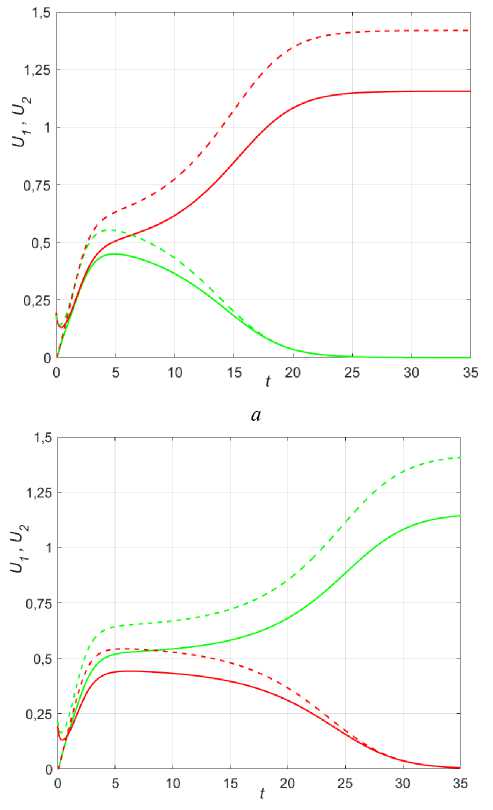
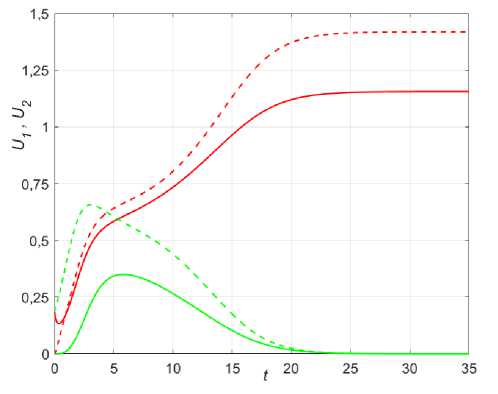
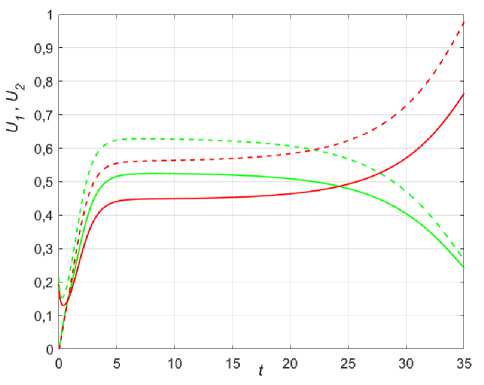
Рис. 9. Динамика различных сценариев популяций при колонизации в условиях неопределенности для двух точек x = 0,25 (сплошные) и x = 0,45 (пунктир), в 1 = 1,35; в2 = 1,4; k = 0,01; Р ( х ) = 0,5 + sinn x .
а - U 10 = U 0 , е = 1; б - U 1 0 = U 0 , е = 0,1; в - U 10 = 0,234sin ( 6nx ) , U 2 = 0,2sin ( 6n x ) , е = 1;
г - U 0 = 0,234sin ( 6nx ) , U 0 = 0,2sin ( 6nx ) , е = 1,2
