Математическая модель конкуренции популяций на загрязненной территории

Автор: Колпак Е.П.
Журнал: Бюллетень науки и практики @bulletennauki
Рубрика: Физико-математические науки
Статья в выпуске: 9 т.10, 2024 года.
Бесплатный доступ
Техногенное воздействие на окружающую среду приводит к изменению видовой структуры экосистем. Внешнее ингибирующее воздействие на конкурирующие популяции приводит к изменению их численности. Одной из задач прогнозирования является теоретическая проработка направлений изменения численности популяций. Разработана математическая модель конкуренции двух популяций, учитывающая изменение скорости роста численности популяции и изменение емкости экологической ниши. Дана оценка направлений изменения численности популяции. Модель представлена задачей Коши для системы обыкновенных дифференциальных уравнений.
Выживаемость, математическая модель, популяция, антропогенное воздействие, устойчивость, распределение вероятностей
Короткий адрес: https://sciup.org/14131054
IDR: 14131054 | УДК: 51(075.8) | DOI: 10.33619/2414-2948/106/01
A mathematical model of population competition in a polluted area
Anthropogenic impact on the environment leads to a change in the species structure of ecosystems. An external inhibitory effect on competing populations leads to a change in their numbers. One of the tasks of forecasting is the theoretical study of the directions of population change. The paper develops a mathematical model of competition between two populations, taking into account the change in the rate of population growth and the change in the capacity of the ecological niche. An assessment of the directions of population change is given. The model is represented by the Cauchy problem for a system of ordinary differential equations.
Текст научной статьи Математическая модель конкуренции популяций на загрязненной территории
Бюллетень науки и практики / Bulletin of Science and Practice
УДК 51(075.8)
Техногенные вмешательства в природную среду стали соперничать со многими природными процессами, оказывая мощное воздействие на природные комплексы, вызывая нарушения нормального хода протекающих в различных биогеоценозах процессов. Вредные для всего живого вещества антропогенного происхождения наполняют воздушный и водный бассейны, загрязняя обширные территории [1, 2], Загрязнение среды обитания ведет к кардинальным изменениям условий существования населяющих эти территории растений и животных, может изменить направленность и формы естественного отбора [3], способно изменять генетическую структуру природных популяций [4], привести к уменьшению видового разнообразия и исчезновению отдельных видов [5]. В дополнение к этому изменяется и интенсивность межвидовых взаимоотношений, обусловленная различной реакцией особей на внешние воздействия. У конкурирующих популяций изменяется численность, происходит смена доминирующих видов, некоторые виды исчезают [6].
Проникновение химических веществ в организмы человека, птиц, млекопитающих и рыб происходит через органы дыхания, желудочно-кишечный тракт, кожные покровы и слизистые оболочки. В растения загрязняющие вещества попадают при корневом питании, путем газообмена и обменной адсорбции с поверхности листовой пластинки. По мере поступления в трофические цепи токсиканты аккумулируются в их конечных звеньях, оказывая негативное влияние на клеточном, гистологическом, организменном и популяционном уровнях. Уровень накопленных токсикантов в организме зависит от возраста, пола, путей поступления в организм [4]. Накопление токсикантов сопровождается изменением плодовитости и смертности характерными для каждого вида.
Математическая модель
Для описания динамики численности двух конкурирующих популяций используется локальная «модифицированная» математическая модель Вольтерры [7], учитывающая как межвидовую, так и внутривидовую конкуренцию:
du
— = A u 1 ( 1 - u 1 - Y 1 u 2 ) ,
“~2“ = ^2U1 (1 - u2 - YЩ ) , где u и u2 — численность популяций, ^ и ц2 — удельные локальные скорости роста численности популяций, y и Y — параметры, характеризующие интенсивность конкуренции. Емкости сред обеих популяций приняты в модели (1) равными единице [7].
Система уравнений (1) имеет четыре неподвижные точки
-
1. щ = 0, u 2 = 0.
-
2. u = 1, u 2 = 0.
-
3. u = 0, u 2 = 1.
-
4. u 1 = ( 1 - Y ) / ( 1 - / 1 / 2 ) , u 2 = ( 1 - Y 2 ) / ( 1 - Y 1 Y 2 ) ,
если y > 1 и y > 1, или Y < 1 и.
Первая точка является неустойчивой, вторая будет устойчивой, если y < 1 и Y > 1, а третья, если Y > 1 и у 2< 1. Четвертая стационарная точка реализуется и будет устойчивой, если одновременно выполняются неравенства y < 1 и у 2< 1. То есть, если внутривидовая конкуренция у популяций слабее межвидовой ( у2 <1 и y <1), то совместное существование популяций будет устойчивым [7].
Тяжелые металлы, выбрасываемые предприятиями в окружающую среду, образуют токсичные для живых организмов вещества. Токсиканты постукают в организмы либо непосредственно, либо по трофическим цепям. Накопление их в организме со временем приводит к внутренним изменениям. В модели предполагается, что изменения метаболизма вызывают понижение плодовитости особей [3]. Предполагается также, что рождаемость особей с увеличением количества токсикантов P уменьшается по гиперболической зависимости
R (P) =
1 + ap
1 + a2P,
где a^ и a2 — положительные параметры такие, что a j < a 2.
Часть среды обитания при антропогеном давлении может стать недоступной для особей популяции, или часть трофического ресурса может быть уничтожена. Этот фактор в модели учитывается через уменьшение емкости экологической ниши популяции. Уменьшение емкости ниши K происходит по гиперболической зависимости:
K (P) =
1 + bP
1 + b2P
где b и b2 — положительные параметры такие, что b < b 2.
С учетом этих предположений модель конкуренции двух популяции (1) в загрязненной зоне сводится к задаче Коши для системы двух дифференциальных уравнений
|
du |
' 1 + aP 1 + bP b |
|
|
—1 = m |
1 11 /‘ тР Y1 и 2 |
, |
|
dt \ |
1 + a^p 1 + byp |
|
|
du |
( 1 + a?p 1 + bp |
b |
|
= ^ u 2 |
---—---—— и - Y U ^ 1 + a2p 1 + b22P 2 2 |
где a:i и b ( i, j = 1,2) - параметры, удовлетворяющие неравенствам ij ij a11 — a12
a 21 — a 22 , b12 — b 11, b22 — b21 •
На параметры Y и Y накладываются ограничения: 0 — Y 1 < 1 , 0 — Y < 1 , тем самым предполагается, что в отсутствие токсикантов ни одна из популяций не гибнет.
При P = 0 (токсиканты отсутствуют) эта модель переходит в модель (1). Удельная скорость роста популяций в зависимости от значения P при выполнении условий (3) уменьшается, но не может уменьшиться более чем на величину ^a{ j / a12 для первой популяции, и на величину ^2a 2j / a22 для второй популяции. Этим учитывается факт ограниченного накопления токсикантов организмами особей популяций и неоднородность накопления внутри самих популяций [3, 4].
Загрязнители среды могут уничтожить часть трофического ресурса или сделать его недоступным для особей популяций. Емкости ниш популяций при выполнении неравенств b 11 > b 12 и b 2j > b22 в (2) с увеличением количества загрязнителей будут уменьшаться. Предполагается, что емкость ниши первой популяции не может стать меньше величины b 1 / b 2, а второй — b 2j / Ъ22 . Этим учитывается то обстоятельство, что небольшая часть популяций может выдержать антропогенную нагрузку.
Нетривиальная неподвижная точка системы уравнений (2) определяется через параметры модели:
и =
j_ ( i + bp i + ap D ^ 1 + b22P 1 + anP
- Y 1
i + aPл 1 + a 22 P J
1 (1 + bP 1 + a2,P 1 + aP У u7 = — ---—--21--Y----11—
-
2 D ^ 1 + bnP 1 + a22P 1 + anP J
где
D =
1 + bP 1 + b21P
1 + bnP 1 + b22P
Поскольку при P = 0 эта стационарная точка существует и является устойчивой (поскольку y <1 и Y i <1), то в силу непрерывности правых частей выражений (4) от P , она будет существовать и при малых значениях P .
Собственные значения матрицы Якоби
J =
- ш ---11 и
1 1 + b12P 1
-Y1PU1
-Y 2 ^ 2 U 2
- ш ----21 u2
2 1 + b22P 2
системы уравнений (2) в стационарной точке (4) будут иметь отрицательные вещественные части, если выполняется неравенство
D =
1 + bP 1 + b2iP
- Y 1 Y 2 > 0.
1 + b12P 1 + b22P
Поскольку рассматривается модель, для которой выполняются условия (3), то, соответственно неравенство D > 0 при 0 < y < 1 и 0 < Y2 < 1 будет выполняться. То есть, если стационарная точка (4) реализуется, то она будет устойчивой.
Распределение вероятностей
Реализация математических моделей популяционной биологии возможна во встроенном модуле SIMBIOLOGY среды программировании математического пакета Matlab [8]. Имитационная модель основывается на случайном переборе параметров модели (2) из заданного диапазона значений [9] с учетом соотношений (3). При заданном количестве вариантов выбора параметров строятся вероятности распределения отклонений стационарных значений численностей популяций при антропогенном давлении от естественных значений.
На Рисунке 1 приведены распределения стационарных значений численностей популяций при Y2 = 0.6, Yi = 0-1 и P = 1.5 для параметров выбранных случайным образом из диапазонов an е (1,2) , a2j g (1,2) , a12 g an + (0,1) , a12 е an + (0,1) , b22 e (0,1) , bn g (0,1) , bn e b12 + (0,1) , b21 e b22 + (0,1) . На Рисунке 2 приведено вероятностное распределение численности популяций для 2 000 вариантов выбора параметров системы уравнений (2). Такой вариант выбора параметров обеспечивал выполнение неравенств (3). Пунктирными линиями на Рисунке 1 и Рисунке 2 отмечены стационарные значения численностей популяций для чистой территории. Сплошной линией на Рисунке 1 отмечена граница области, в которой лежат стационарные значения численности популяци0439.
В рассмотренном варианте параметров большие потери несет та популяция, которая в отсутствие токсикантов имеет большую численность (на Рисунке 2 — u ). Знание реальных параметров, входящих в модель (2), дает возможность прогнозировать степень влияния антропогенной нагрузки на численность популяций для реальных экосистем [10, 11].
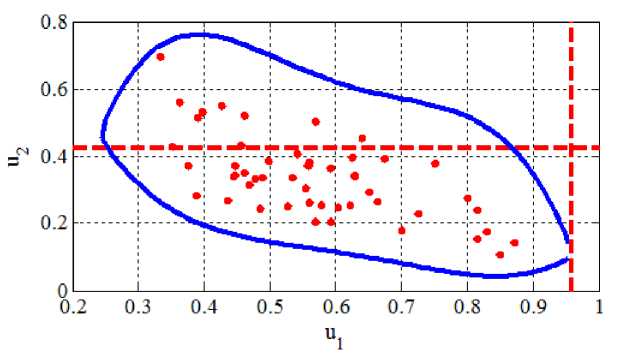
Рисунок 1. Распределения стационарных значений численностей популяций
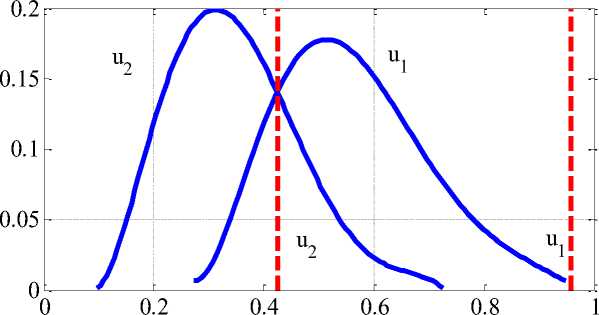
u1, u2
Рисунок 2. Распределение вероятностей численности популяций при антропогенном давлении
Заключение
Таким образом, разработанная модель прогнозирует сдвиг распределений вероятностей в сторону уменьшения численности обеих популяций при наличии токсикантов, по сравнению с их численностью в «чистой» среде. Наибольшие потери численности будут у популяций, которые до антропогенного давления имели большую численность. Численность малочисленных популяций может вырасти за счет уменьшения давления конкурирующих популяций.
Список литературы Математическая модель конкуренции популяций на загрязненной территории
- Пегов С. А. Антропогенное воздействие на биосферу // Труды Института системного анализа Российской академии наук. 2009. Т. 42. С. 5-32.
- Шевцова О. В., Добротина Е. Д., Гончарова А. Б., Недашковский А. П. Химические характеристики снежного покрова в высокоширотной арктике (мыс Баранова, остров Большевик, архипелаг Северная Земля) // Лёд и снег. 2022. Т. 62. №4. С. 564-578. https://doi.org/10.31857/S2076673422040152
- Моисеенко Т. И. Биодоступность и экотоксичность металлов в водных системах: критические уровни загрязнения // Геохимия. 2019. Т. 64. №7. С. 675-688. https://doi.org/10.31857/S0016-7525647675-688
- Ивантер Э. В., Медведев Н. В. Экологическая токсикология природных популяций птиц и млекопитающих Севера. М.: Наука, 2007. 229 с.
- Катаев Г. Д. Воздействие выбросов медно-никелевого предприятия на состояние популяций и сообществ мелких млекопитающих Кольского полуострова // Nature Conservation Research. Заповедная наука. 2017. Т. 2. №Приложение 2. С. 19-27. https://doi.org/10.24189/ncr.2017.033
- Касиков А. Г. Пылевые выбросы медно-никелевого производства и последствия их воздействия на организм человека в условиях Крайнего Севера // Вестник Кольского научного центра РАН. 2017. №4. С. 58-63.
- Горыня Е. В., Колпак Е. П., Гасратова Н. А., Гончарова А. Б. Математическая модель иерархической конкуренции // Перспективы науки. 2023. № 8 (167). С. 103-108.
- Гончарова А. Б., Виль М. Ю. Имитационное моделирование лечения онкологического заболевания с использованием приложения Matlab Simbiology // Моделирование систем и процессов. 2021. Т. 14. №3. С. 90-96. https://doi.org/10.12737/2219-0767-2021-14-3-90-96
- Гончарова А. Б. Постановка предварительного медицинского диагноза на основе теории нечетких множеств с использованием меры Сугено // Вестник Санкт-Петербургского университета. Прикладная математика. Информатика. Процессы управления. 2019. №4. С. 529-543. https://doi.org/10.21638/11702/spbu10.2019.409
- Даувальтер В. А., Кашулин Н. А. Прогнозирование долговременных изменений пресноводных региональных систем рыбного хозяйства Арктики // Вестник Мурманского государственного технического университета. 2012. Т. 15. №1. С. 171-180.
- Кривополенова С. Д., Гончарова А. Б. Первичный анализ данных для построения системы поддержки принятия решений // Процессы управления и устойчивость. 2019. Т. 6. №1. С. 250-254.
