Математическое моделирование экосистемы озера Байкал: вчера, сегодня, завтра

Автор: Зилов Е.А.
Журнал: Известия Самарского научного центра Российской академии наук @izvestiya-ssc
Рубрика: Водные экосистемы
Статья в выпуске: 1 т.8, 2006 года.
Бесплатный доступ
Кратко рассмотрены первые математические модели экосистемы оз. Байкал. Описана модель антропогенных возмущений экосистемы оз. Байкал, построенная на основе полевых натурных экспериментов. Результаты моделирования указывают на большую устойчивость летнего планктонного комплекса, чем подледного, качественное сходство реакции экосистемы планктона на атрофирование и токси- фикацию. способность сообщества справляться с разовыми повышениями концентраций загрязняющих веществ без необратимых последствий, но неизбежность перестройки сообщества при хроническом поступлении загрязняющих веществ. Экосистема оз. Байкал практически функционирует уже на другом уровне, отличном от невозмущенного состояния, хотя и характеризующемся пока орбитальной устойчивостью.
Короткий адрес: https://sciup.org/148197822
IDR: 148197822 | УДК: 574.5:682.5:285.2
Mathematical modelling of the lake Baikal ecosystem: yesterday, today, tomorrow
The first models of lake Baikal created in 1970th are briefly reviewed. The model of the lake Baikal ecosystem anthropogenic disturbances, developed on the basis of field in situ experiments is presented. Incomes of nontoxic mineral and organic compounds, nutrients (nitrate and phosphate), organic (phenol compounds, oil products) and non-organic (heavy metals) toxicants with tributaries, coasts, atmosphere served as inputs of the model. The results of model experiments demonstrate higher stability of summer plankton community in comparison with under-ice one. The reaction of plankton to toxification and eutrophication was qualitatively similar. Now the lake Baikal ecosystem functions at different from non-perturbed level, though it is still stable.
Текст научной статьи Математическое моделирование экосистемы озера Байкал: вчера, сегодня, завтра
,.
. .-
. ,-
.-
, ,-
,-
,-
..
, ,-
.
Экосистема озера Байкал стала одним из
-
-
.
-
.
.. , . . ..-
[1, 2].-
: « », «-
», « », «-
», « », « » «».
использовался диапазон изменений каждой 0 +1, 0« -
», +1 «-
», « »,,
0,5,
, «-
» 0, «-
»
-
1, «»
+1.
«»
6:
-
- , - , -, -
- - , - ,--
- .-
-
- ,-
- , ,-
- ,
,-
,
-
-, ,,
.- трация биогенных элементов определяется
,-
.- речисленных факторов происходят сезонные
,-
,, -
,
,
.
Остальные подмодели не имеют сезонной
.«»
«-
»-
,«-
», « », «»
«Омуль» среднегодовыми количествами рыб, поедающих рачка.
Для подмоделей рыб среднегодовое коли -чество макрогектопуса служит положитель-но действующим фактором, а для планктона - отрицательным. СС убмодели рыб имеют возрастные группы: по 3 - у длиннокрылки и желтокрылки, 6 с у голомянки и 10 су омуля. Каждая из них зависит от планктона и мак -рогектопуса, смуль - еще и от бентоса. Омуль и желтокрылка вылавливаются, причем вылов регулируется величиной промыслового усилия.
X ( t ) = Ax ( t ) + Bx ( t -1) + Cu ( t ), где
u ( t ) = ( u i( t ),..., u 2o( t )), u ( t ) e [0,1] x ( t ) = ( x 1( t ),..
Желтокрылка, ссиннокрылка и голомянка поставляют пищу нерпе, соличество которой сказывается на них и определяется промыс-лом, который зависит от промыслового усилия . Наконец бентос зависит от температурных и кормовых условий через планктонный блок и от выедания омулем.
Всего в модели 120 переменных состояния , можно варьировать 20 сходных переменных , 3 пессменных - полезный выход в виде вылова желтокрылки и омуля, добычи нерпы .
Модель представлена системой линейных уравнений:
- вектор входных переменных для года t ,
, x 120( t )), x, ( t ) e [0,1] - вектор фазовых переменных в тот же год,
A - матрица коэффициентов влияния внутри года,
B - матрица межгодовых связей,
C - матрица зависимости переменных модели от внешних условий.
При этом:
120 20
£( « ДМ)+ Е С «1 = 1, ‘ =1,2,...,120.
j =1 к =1
У стандартной точки системы все коорди-наты равны 0,5.
Эта модель самими авторами расценива-лась как промежуточный этап в работе и по -зволяла получать только качественные резуль -таты. Тсм не менее ее исследование позволило найти ряд зависимостей между абиоти-ческими факторами, обнаружить, сто биотические компоненты мало зависят от темпе -ратуры воздуха, но сильно зависят от инсоляции и ветровых условий, сыделить наиболее чувствительные к солнечной радиации (фитопланктон, бактерии, коловратки, циклоп ) и ветровой активности (фитопланктон) элементы сообщества. Ряд положений исходно полученных в экспериментах с моделью сейчас подтвержден анализом данных поле -вых наблюдений [9].
Следующей стала энергетическая модель пелагического сообщества оз. Байкал Л.Я. Ащепковой, В.И. Гурмана и О.М. Кожовой [1]. Модель, как и предыдущая, точечная. В модели рассматривались потоки энергии в слое
0-250 м.
В модель включены следующие компонен-сы: фитопланктон ( x 1), спишура ( x 2), циклоп ( x 3), макрогектопус ( x 4), смуль ( x 5), пелагические бычки ( x 6), соломянка ( x 7), серпа ( x 8), бактерии ( x 9), детрит ( x 10). В модели приняты следующие обозначения:
b . - биомасса z'-со компонента, г. - поток энергии от z-ro компонента к j-му, q . - энергетический обмен компонента i, m - потери энергии со смертью i-со компонента , u . - неусвоенные остатки пищи компонен-с a i,
К. - напряженность трофических отноше-ний компонента i, j - для z-ro компонент а в питании j-го, cjmax - удельный максимальный рацион, A - годовой приток аллохтонной органики ,
S 1 - седиментация органики,
S 2- сток органики через Ангару.
Потоки энергии , функции биомасс вычислялись по формуле Меншуткина,- лева и Вольтера не дали желательной устой-:
j1e ii ri j(bi,bj) cmjaxbjА ij i , j max j ij, кi cmj axbjxij cmax (bi ) = а je jbj.
Средняя поправка Меншуткина ( формуле для r ij ( b i , b j ))
0,8. v V- ловия, bj->0,- ный рацион увеличивается в 2- нению с максимальным рационом для сред-него значения биомассы .
энергии со смертью линейно зависели от био-масс компонентов,- оны,- циона.
:
db
-
1 r 01 q 1 m 1 "Е r 1 j , dt j
-
d dbt i -=Е r ij q i m i u i -Е r ij , i 2,3,...,9,
-
d dbt 10 -=Е m i +Е ui + r 109 + A S 1 S 2 , dt i i
где S1 = kb10, S2 = mb10. k значение 6,7, m - 20.
омуля и нерпы были введены потоки r50 r80, означающие их добычу и связанные с соот-ветствующими биомассами линейно . - рименты с полученной моделью показали связь первичной продукции экосистемы с притоком аллохтонной органики .
была подчеркнута гипотетичность применен-ных в модели коэффициентов и необходи-мость их уточнения в полевых эксперимен-тах на озере.
Сохранив те же функции для имитации биологических процессов В .В.
соавторами [15]- дель озера,- та на 65 484 2,- ная толща была разбита на трофогенную (05 0 ).
модель неплохо отражала реальную динами-ку экосистемы, распространения попавшего в озеро веще-ства, экосистемы в одной точке на другие районы.
Затем вот уже 20 ни одной попытки построения имитацион-ной модели естественной динамики экосис-темы озера. , - рование естественной динамики экосистем промежуточного ( океаном) , [3, 14]:
-
- существуют факторы, -
- ном на динамику не влияют, -
- ,
-
- для каждой фазы динамики определяю-щими являются свои собственные факторы,
-
- характер и уровень связей для различ-ных фаз существенно различаются,
-
- многие закономерности со временем раз-рушаются.
Если мы рассмотрим естественную дина-мику компонентов экосистемы пелагиали оз. Байкал , , всех компонентов непрерывно колеблются [17]. -видимому, колебаний зависят не только от внутренних механизмов взаимодействия компонентов, и от внешних факторов, (на- пример, ), - генных [5].
В связи с необходимостью оценки послед-
-тему было решено ограничиться построени-
-системы озера., что естественная динамика рассматривается как некая прямая («нулевое»), -
(увеличение) или отрицательным (уменьшение) [18-20, 22].
В первую очередь надо было выбрать ком-поненты,- дель .- ных моделей из биологических компонентов были взяты фито-, зоо- и бактериопланктон. Гидрохимические компоненты были пред -ставлены в модели суммарной минерализа-цией вод, концентрациями биогенных элементов (нитратный азот и фосфатный фосфор ), растворенное органическое вещество (измеряемое как ХПК). Кроме того, учтены главные токсичные вещества, попадающие в озеро, главным образом в результате хозяйственной деятельности: нефтепродукты, фенольные соединения, ионы тяжелых металлов .
В качестве исходной модели рассматрива-лась следующая:
dR / dt = Q ( R-R *) + u + r , (1)
где R - вектор состояния экосистемы (биомассы и концентрации компонентов),
R* - лектор невозмущенного состояния экосистемы (сллднее за много лет значение этого компонента для данного времени года), и - вектор внешних воздействий (поступление загрязнений ), r - вектор переноса компонентов (миграции , перенос течениями и диффузия между ячейками ),
Q - матрица взаимовлияний компонентов экосистемы (каждый элемент qj показывает изменения компонента i в единицу времени при отклонении компонентаj ла единицу, диагональные элементы qu отражают динамику возвращения i-го компонента в исходное состояние после отклонения, это скорости самовосстановления биологических компо- dZ't /d \»Z +(1/Vt)£{P.Z; j=1 l^L
где L - агрегат соседних камер, P , D л матрицы турбулентности и диффузии, рассчитанные Е.А. Цветовой, V лл вектор объемов камер .
При моделировании наиболее сложной и трудоемкой является задача информационно -го обеспечения. Основываясь на опыте построения моделей О.М. Кожовой, ле было решено провести на основе натурных экспе-риментов, лаправленных на определение неизвестных параметров. Для этого была применена следующая технология.
нентов и константы скоростей распада заг-рязнителей ).
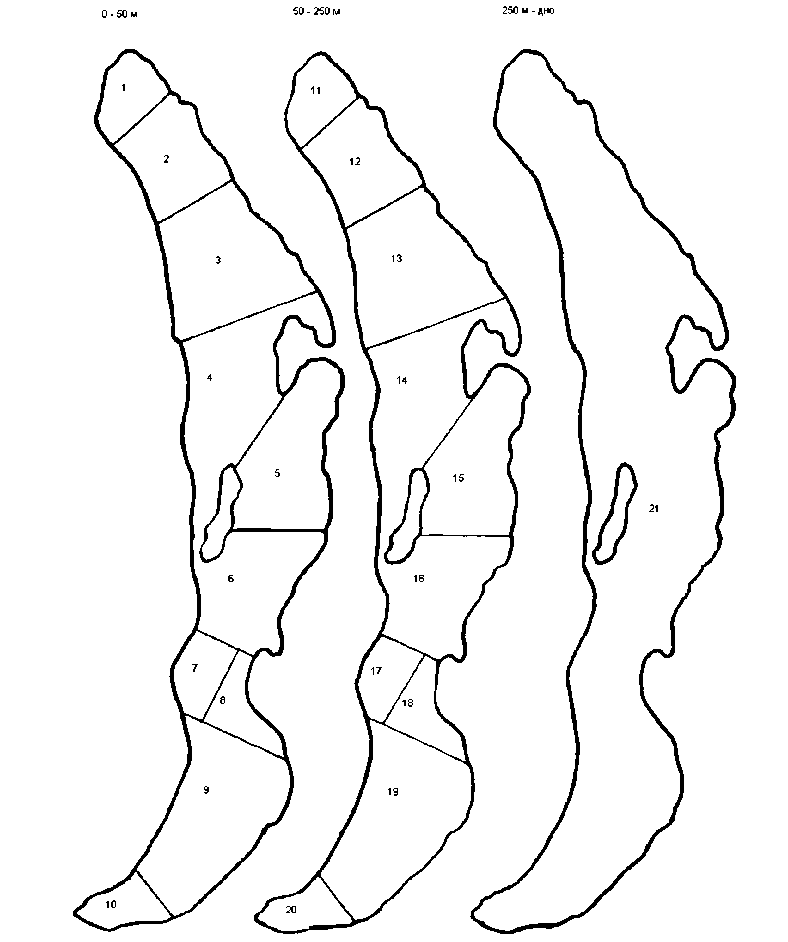
Была принята следующая схема разбиения водного тела на камеры (см. рис. 1). По глубине было выделено уже не два, а с учетом опыта работы с моделью В .В. Меншуткина и др. [15], три слоя, различающихся по физическим и биологическим показателям:
-
• 0-50 м (трофогенный, лорошо освещенный слой со значительными годовыми перепа-дами температур, фито-, зоо-, бактериопланктон ),
-
• 50-250 м (очень слабо освещенный слой, перемешивание, течения, меньшие перепады температур, зоо- и бактериопланктон),
-
• 250 м - дно (постоянные температурные условия, полная темнота, бактериопланктон ). По горизонтали 2 верхних слоя были разделены на 10 камер каждый, нижний слой рассматривался как одна камера.
Использование вектора некоего невозму-щенного состояния экосистемы предполага-ет, что мы знаем это невозмущенное состояние . Поскольку это не так, модель (1) была заменена моделью, выраженной в терминах отклонений [22]:
N dZj/ dt^QyZ^Ui, (2)
j =1
где Z л вектор отклонений показателей состояния , N лл число показателей, U лл вектор управления .
С учетом гидрологического блока мы по -лучаем:
-
-P k Z' .+ D ki ( Z l- Z k )}] U k , (3)
Пусть объект описывается следующей ма-тематической моделью:
dZ / dt = f ( t , z , a ) (4)
При наличии m реализаций и наблюдаемости части компонент:
Z j ( t ) = ( Z lj ( t ),..., Zmjt t )), j = 1 m
. 1. . ,
При задании m начальных условий:
Z i tt 0) = Z 0, i = 1, m .
надо минимизировать функционал t fm m
-
I= ВЁ 'LL Z Z j) ) 2 dt+L^jt^ t k )- Z j ( t k »2 > (5) t 0 i =1 ) j i =1 ) mMj
где i - номер компонента вектора Z, j - номер испытания,
M j - множество номеров компонент вектора Z , наблюдаемых в ) -м испытании,
Рi., у*. - aaniaua eiyooeoeafou.
На основе экспериментального материа-ла [6-8] определялись величины отклонений z в на каждый учитываемый момент времени и коэффициенты системы:
z ‘^^ч , ' , к =ш (6)
где первоначальные коэффициенты q , определялись экспертной оценкой, a, b g g принимались равными 1. После минимизации функционала и определения новых q & система вновь интегрировалась, полученные значения z сравнивались с экспериментальными . Пр и наличии существенных расхождений процесс повторялся с новыми b и д .
Таким образом, для определения коэффициентов взаимовлияния для модели из N компонентов было необходимо поставить N серий опытов, в каждой из которых отклоняя на заданную величину один из компонентов, а затем определяя концентрации всех компо-нентов до конца экспозиции. В нашем случае вносились следующие отклонения:
-
• добавки сульфата натрия (повышение минерализации ),
-
• добавки нитратного азота и фосфатного фосфора,
-
• добавки аллохтонной органики (пептон)
-
• добавки фенольных соединений (пирокатехина ),
-
• добавки нефтепродуктов (водная эмульсия
дизельного топлива),
-
• добавки ионов тяжелых металлов (хлорид кадмия ),
-
• бактериопланктона,
-
• фитопланктона и
-
• зоопланктона.
Два последних концентрировались с по -мощью соответствующих планктонных сетей и вносились в экспериментальные мезокос-мы (объемом 2 м3) непосредственно из озера .
Использование алгоритмов, разработанных и реализованных в виде программного комплекса в ИрВЦ ВСФ СО РАН, позволило решить поставленные задачи. На основе данных , волученных в натурных экспериментах на оз. Байкал, Ввши рассчитаны матрицы взаимовлияния компонентов для подледного и летне - осеннего сезонов.
После определения компонентов матрицы Q, в первую очередь был проведен анализ устойчивости модели. Она оказалась внутренне устойчива.
Сценарии модельных экспериментов вы -бирались на основе доступных опубликован -ных данных. Для современного уровня хозяйственной активности были приняты следу -ющие величины поступления химических веществ в озеро (см. табл. 1).
Сначала были проведены эксперименты
1. ( -1)
хозяйственной деятельности
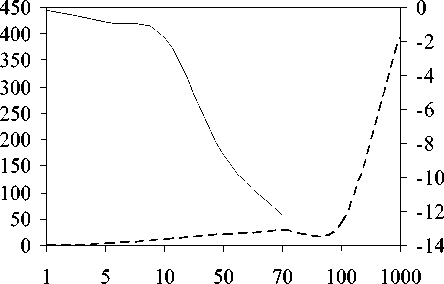
Были проанализированы ответы модели на токсификацию и эвтрофирование. Сравнивались воздействие разового стартового внесения токсикантов и биогенных веществ и их хронического действия. Подледное сообщество оказалось значительно чувствитель-нее летнего и к токсификации, и к эвтрофи-р вванию (рис. 2) и медленнее возвращалось к исходному состоянию, чем летнее (табл. 2
-5
-10
-15
-20
-25
1 10 100 900 1000 3000 3800
Рис. 2. Действие (А) смеси нитрата и фосфата натрия ([4N+1P] мкг Л" )и(Б) фенольных соединений на общую биомассу планктона (мкг л-1) подо льдом (правая шкала) и летом (левая шкала, пунктир)
Таблица 2. Максимальное отклонение компонентов (мкг Л"1) в ответ на разовое поступление фенольных соединений (100 мкг Л"1)
|
Компоненты |
Подледное сообщество |
Летнее сообщество |
||
|
от |
до |
от |
до |
|
|
Биогены |
-1.52 |
0.46 |
-1.80 |
0.25 |
|
Фитопланктон |
-4.12 |
1.21 |
-1.19 |
2.68 |
|
Зоопланктон |
-1.11 |
1.85 |
-0.51 |
0.48 |
|
Возвращение к исходному состоянию через (сут .) |
34 |
23 |
||
Таблица 3. Максимальное отклонение компонентов (мкг Л"1) в ответ на разовое поступление биогенных соединений (10 мкгР Л"1и 40 мкгN Л"1)
|
Компоненты |
Подледное сообщество |
Летнее сообщество |
||
|
от |
до |
от |
до |
|
|
Фенолы |
0.00 |
1.39 |
0.00 |
2.95 |
|
Фитопланктон |
-23.63 |
8.61 |
-6.69 |
24.63 |
|
Зоопланктон |
-9.95 |
21.69 |
-1.55 |
3.68 |
|
Возвращение к исходному состоянию через (сут .) |
36 |
24 |
||
Таблица 4. Отклонение компонентов (мкг Л"1) в ответ на хроническое поступление фенольных соединений (100 мкг Л"1 сут. 1)
Таблица 5. Отклонение компонентов (мкг Л"1) в ответ на хроническое поступление биогенных соединений (10 мкгР Л"1и 40 мкгN Л"1 сут. 1)
|
Компоненты |
Подледное сообщество |
Летнее сообщество |
|
Биогены |
38.07 |
42.10 |
|
Фенолы |
0.00 |
1.81 |
|
Фитопланктон |
-20.30 |
17.34 |
|
Зоопланктон |
11.57 |
2.26 |
|
Переходит в новое стационарное состояние через (сут.) |
43 |
29 |
Б
А
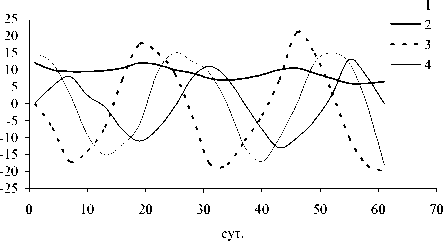
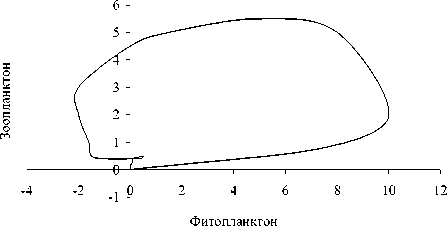
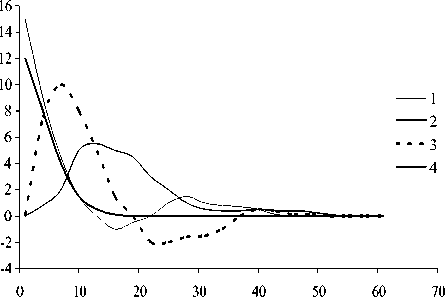
Рис. 3. Динамика отклонений (мкг л -1) ряда показателей состояния экосистемы в летне-осенний период (А) и фазовый портрет системы (Б): 1 - суммарная концентрация нитратного азота и фосфатного фосфора , 2 - концентрация фенольных соединений, 3 - концентрация фитопланктона, 4 - концентрация зоопланктона
Б
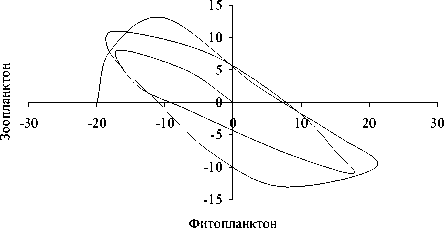
Рис. 4. Динамика отклонений (мкг л-1) ряда показателей состояния экосистемы в зимне-весенний период (А) и фазовый портрет системы (Б): 1 - суммарная концентрация нитратного азота и фосфатного фосфора, 2 - концентрация фенольных соединений, 3 - концентрация фитопланктона, 4 - концентрация зоопланктона
обращении к фазовым портретам (рис . 3Б и 4Б). Видно, что система стремится к асимп-тотической устойчивости в летний период и демонстрирует орбитальную неустойчивость в подледный период. Это еще раз подтверж-дает высказанное выше предположение о большей чувствительности подледного план-ктонного комплекса к возмущающим воздей- ствиям.
Надо отметить, что полученные на мате-матических моделях результаты реалистично отражают процессы, наблюдавшиеся в мезо-космах, как в ходе наших полевых экспери-ментов, так и отмечавшиеся для оз. Байкал другой группой исследователей [10]. В натур-ных экспериментах с мезокосмами на озере
A
-10
-20
0 5 10 15 20 25 30 35 40
.
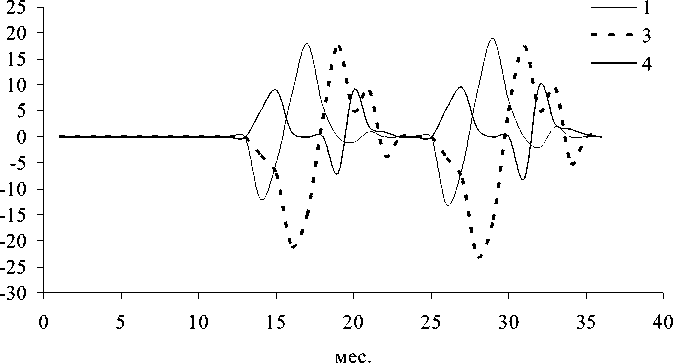
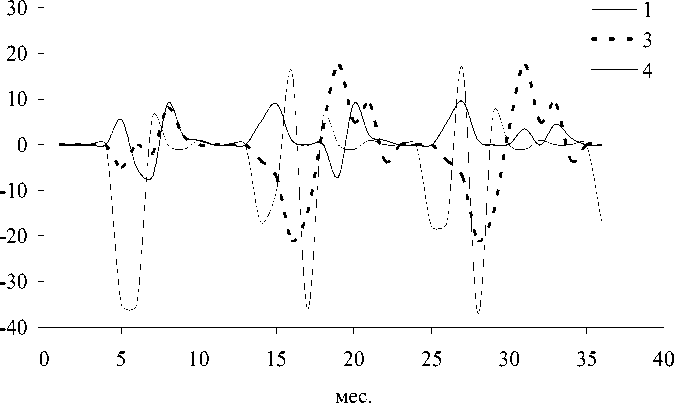
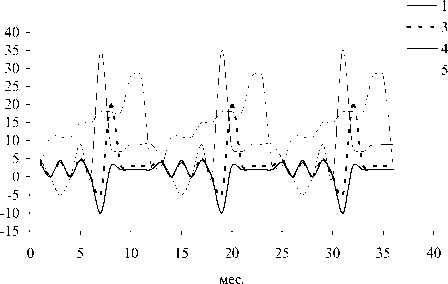
Рис. 5. Динамика отклонений (мкг л-1) ряд а показателей состояния в поверхностном слое при поступлении загрязнителей с атмосферными осадками (А); пр идонном слое при поступлении загрязнителей с атмосферными осадками (Б); поверхностном слое (Северный Байкал) пр и поступлении загрязнителей с водами р. Селенги (В): 1 -суммарная концентрация нитратного азота и фосфатного фосфора, 3 - концентрация фитопланктона, 4 - концентрация зоопланктона, 5 - концентрация бактериопланктона
Байкал показана большая чувствительность
,-
,
,-
(
), .,
,-
(
-
),-
.
,-
-
,-
.
-
-
-
[ 23]. ,-
-
- лучаем следующую картину в поверхностном
-
( . 5 ).,
-
-
-
) ( . 5 ).-
-
- .-
- вания начинают отмечаться отклонения от
-
-
( . 5 ). -
-
- ,-
- ,-
-
- ( . . 6 ). ,-
- .-
- ,-
- ,
( . 6 ).
,,
, подтверждение при анализе трендов ряда гидрохимических показателей по материалам наблюдений Государственного комитета по
, -
. , -
-
[12, 13].
Модель возмущений явилась логичной наследницей первых моделей экосистемы
. -
, -
, - ет разбиение водной толщи на камеры по
-
. ,-
,«-
». -
.-
Б
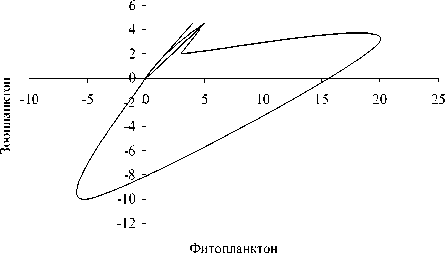
Рис. 6. Динамика отклонений (мкг л-1) ряд а показателей состояния в поверхностном слое при поступлении загрязнителей с водами притоков, атмосферными осадками, при таянии льда (А) и фазовый портрет системы (Б): 1 - суммарная концентрация нитратного азота и фосфатного фосфора, 3 - концентрация фитопланктона, 4 - концентрация зоопланктона, 5 - концентрация бактериопланктона тивно начинают использоваться модели структурной динамики, в частности на осно-ве целевых функций, таких как эксэргия [4, 20], ведется работа по разработке самоорга-
Список литературы Математическое моделирование экосистемы озера Байкал: вчера, сегодня, завтра
- Ащепкова Л.Я., Гурман В.И., Кожова О.М. // Энергетическая модель пелагического сообщества оз. Байкал // Модели природных систем. Новосибирск, 1978.
- Ащепкова Л.Я., О. М. Кожова, В. В. Мен-шуткин. Модель сезонной динамики пелагического сообщества оз. Байкал//Модели природных систем. Новосибирск, 1978.
- Брусиловский П.М. Коллективы предикторов в экологическом прогнозировании. Саратов: Изд-во Сарат. ун-та, 1987.
- Зилов Е.А. Использование эксэргии для оценки здоровья водных экологических систем//Экосистемы и природные ресурсы горных стран. Новосибирск: Наука, 2004.
- Зилов Е.А., Сериков П.Е. Использование гиперциклов для описания автоколебательных процессов в моделях экосистем//Математические проблемы экологии. Новосибирск, 1992.
- Зилов Е.А., Стом Д.И., Башарова Н.И.и др. Использование мезокосмов для информационного обеспечения математических моделей водных экосистем//Экологические исследования Байкала и байкальского региона. Иркутск: Изд-во Иркут. ун-та, 1992. Т. 2.
- Зилов Е.А., Стом Д.И. Использование физического, математического и комплексного моделирования в прогнозировании поведения водных экосистем под действием антропогенных факторов//Мониторинг состояния озера Байкал. Л.: Гидрометеоиздат, 1991.
- Зилов Е.А., Рудых А.Р., Стом Д.И. Подледный экотоксикологический эксперимент на Байкале//Гидробиол. журн. 1989. Т. 25, № 4.
- Изместъева Л.Р. Температурный режим и развитие фитопланктона в пелагиали Байкала//Экосистемы и природные ресурсы горных стран. Новосибирск: Наука, 2004.
- Изместъева Л.Р., Кузнецов И.Ю. Опыт пространственного зондирования фитопланкфитопланктона с применением погружного импульсного флуориметра//Оценка продуктивности фитопланктона. Новосибирск, 1993.
- Изместъева Л.Р, Кожова О.М. Опыт организации гиробиологического мониторинга и создания ГИС//Методология оценки состояния экосистем. Ростов-на-Дону: ЦВВР, 2000.
- Изместъева Л.Р., Шимараева С.В. Многолетние изменения содержания хлорофилла а в пелагиали Южного Байкала в период прямой температурной стратификации//Экосистемы и природные ресурсы горных стран. Новосибирск: Наука, 2004.
- Кожова О.М. Гидробиологическому мониторингу Байкала -50 лет//Проблемы сохранения биоразнообразия. Новосибирск, 1998.
- Кожова О.М., Павлов Б.К Экологическое прогнозирование и состояние планктона Байкала//Изменчивость природных явлений во времени. Новосибирск: Наука, 1982.
- Меншуткин В.В., Кожова О.М., Ащепкова Л.Я., Кротова В.А. Камерная модель динамики экосистемы озера Байкал с учетом трехмерной циркуляции вод//Математическое моделирование водных экосистем. Л., 1981.
- Сериков П.Е., Зилов Е.А. Аналитическое моделирование водной экосистемы на основе методов неравновесной термодинамики//Оценка состояния водных и наземных экологических систем. Новосибирск: Наука, 1994.
- Kozhova O.M., Izmest'eva L.R. (eds.) Lake Baikal: Evolution and Biodiversity. Leiden: Backhuys Publishers, 1998.
- Gurman V.J., Rosenraukh D.M., Stom D.J., Silow E.A. Mathematical modelling of perturbations of lake Baikal ecosystem and identification of it on the basis of experiments//Computer science for environmental protection. New York: Springer-Verlag, 1991.
- Silow E.A. Experimental approach to the development of a management and conservation strategy for the Lake Baikal ecosystem//Abstr. of Intern. Conf. on Ancient Lakes. Shiga, Japan, 1997.
- Silow E.A. The use of two lumped models for the analysis of consequences of external influences on the lake Baikal ecosystem//Ecological Modelling. 1999. Vol. 121.
- Silow E.A. Mathematical model of ecosystem perturbation//Handbook of environmental and ecological models. Baton Rouge: Lewis Publishers, 1995.
- Silow E.A., Gurman V.J., Stom D.J., Rosenraukh D.M., Baturin V.I. Mathematical models of lake Baikal ecosystem//Ecological modelling. 1995. Vol. 82.
- Silow E.A., Baturin V.A., Stom D.J. Prediction of Lake Baikal ecosystem behaviour using an ecosystem disturbance model//Lakes & Reservoirs: Research and Management. 2001. Vol.6, №1.
