Механизм эндокринной регуляции полового цикла и нормализация репродуктивной функции у телок голштинской породы

Автор: Степашин С.Н.
Статья в выпуске: 1 т.245, 2021 года.
Бесплатный доступ
В крупном молочном предприятии Рязанской области проведены исследования механизма эндокринной регуляции эстрального цикла, а также изучены пути нормализации репродуктивной функции. Экспериментальные данные получены на телках голштинской породы случного возраста. В сравнительном аспекте оценена экономическая эффективность применяемых методик. Используемые препараты и методы качественно отличались друг от друга. Впервые доказано сочетанное применение препаратов «PGF Вейкс форте» и «Седимин», позволяющее в среднем на 30 % увеличить показатель вероятности наступления беременности у телок случного возраста.
Половой цикл, гормоны, нейромедиаторы, крупный рогатый скот, воспроизводство
Короткий адрес: https://sciup.org/142229511
IDR: 142229511 | УДК: 591.147.88 | DOI: 10.31588/2413-4201-1883-245-1-185-192
The mechanism of endocrine regulation of the sexual cycle and the normalization of reproductive function in the Holstein heifers
In a large dairy company in the Ryazan region, studies of the mechanism of endocrine regulation of the estrial cycle were carried out, as well as ways of normalization of reproductive function were studied. Experimental data were obtained on the heifers of the Holstein breed of the same age. In a comparative aspect, the economic efficiency of the methods used is assessed. The drugs and methods used differed qualitatively from each other. For the first time, the combined use ofPGF Wakes Forte and Sedimin drugs has been proven to increase the probability of pregnancy in heifers of the same age by an average of 30 %.
Текст научной статьи Механизм эндокринной регуляции полового цикла и нормализация репродуктивной функции у телок голштинской породы
Стратегия современного развития молочной отрасли все больше учитывает физиологический стереотип животного. Мировая практика показывает, что, несмотря на имеющийся научный опыт, молочное скотоводство несет значительные потери от ошибок в сфере воспроизводства, а решающая роль в интенсификации принадлежит динамичному совершенствованию репродуктивной функции до максимально возможных значений [12].
В своей работе Г.Н. Сердюк отмечает роль односторонней селекции в появлении низкорезистентного к заболеваниям, со сниженной воспроизводящей способностью поголовья крупного рогатого скота, где в погоне за высокими качественными и количественными показателями молочной продуктивности зоотехники-селекционеры отодвинули на второй план такой наследуемый показатель как уровень защитных механизмов, в результате чего мы имеем животных с валовым удоем 6000-7000 кг молока в год, при этом срок использования коров ограничивается 1-2 лактациями [13]. С ним согласны ученые Г.М. Туников, А.А. Коровушкин и др., которые отмечают влияние быстрой смены поколений животных ввиду снижения воспроизводительной способности, наступления яловости, при этом животные до выбытия из стада не успевают реализовать свой продуктивный потенциал [14].
Увеличение темпов производства молока и телят определяет важность детальных и комплексных исследований в области точечной регуляции репродуктивной функции у крупного рогатого скота (КРС) молочных пород, а методика все больше основывается на биологических законах целостного организма [3]. В новаторском течении научной мысли значимы методы коррекции и стимуляции репродуктивной функции телок случного возраста с применением фармсредств [2, 7].
В этой связи перспективным и своевременным направлением является изучение физиологических аспектов гормональной коррекции в целях повышения репродуктивных функций крупного рогатого скота и интенсификации воспроизводства стада. Данный вопрос активно изучается научным сообществом. Однако в доступной литературе редко встречается информация о работах в сфере комплексной оценки гормональных программ контроля над воспроизводством молочного стада, а также единое мнение о путях эффективного повышения их результативности, что и послужило основой для разработки актуальной проблемы требующей своего решения.
Функциональная способность яичников контролируется аденогипофизом. По сведениям И.Б. Манухина, Л.Г. Тумиловича, М.А. Геворкяна (2010), фоллитропин (ФСГ) регулирует рост фолликулов и синтез ароматаз, трансформирующих андрогены в эстрогены, рецепторов лютеинизирующего гормона (ЛГ) и ФСГ на клетках гранулёзы фолликула; запускает пролиферацию клеток гранулёзы в фолликулах; стимулирует инкрецию активина, ингибина, инсулиноподобного фактора роста (ИФР); продуцирует эстрадиол. Лютропин (ЛГ) регулирует синтез андрогенов в клетках фолликулов, эстрадиола в доминантном фолликуле, прогестерона в жёлтом теле яичника; способствует овуляции совместно с ФСГ и лютеинизации клеток гранулёзы [10].
В аркуатных ядрах медиобазального гипоталамуса синтезируется гонадотропин-рилизинг-гормон (ГнРГ), который через портальную кровеносную систему, поступает в аденогипофиз, обеспечивая синтез и выделение гонадотропинов. По данным И.И. Дедова, Г.А. Мельниченко (2007)
секреция гонадотропного релизинг-гормона моделируется нейропептидами экстрагипоталамических структур, а также эстрадиолом по принципу обратной связи. Нарастание овуляторного пика эстрадиола сопровождается усилением образования ГнРГ, что в свою очередь ведёт к потенцированию секреции ФСГ и ЛГ аденогипофизом в течение 12-24 часов [6]. И.Б. Манухин, М.А. Геворкян, Н.Б. Чагай (2006) указывают на то, что цирхоральная секреция ГнРГ запускает гипоталамо-гипофизарно-гонадальную систему, функцию которой, по мнению авторов, нельзя считать автономной по причине её моделирования как нейропептидами центральной нервной системы, так и яичниковыми стероидами по механизму обратной связи [9].
На стимулирующее действие нейрогормона аденогипофиз повышает секрецию ЛГ, результатом чего является терминация созревания фолликула и овуляция посредством активизации пролиферативных процессов в их строме и синтеза андрогенов и простагландинов (ПГФ2α и ПГЕ2) а также готовность яйцеклетки к оплодотворению. E. Ioannis и др. (2002) сообщает о том, что после овуляции клетки гранулёзы подвергаются дальнейшей лютеинизации с образованием желтого тела беременности, выделяющего прогестерон под влиянием ЛГ. Авторами установлено прогрессивное нарастание эстрадиола и прогестерона, однако коррелятивной зависимости между данными параметрами ими не было выявлено, о чем свидетельствует независимость путей биосинтеза этих гормонов в жёлтом теле. Схожие данные получены H. Patel [15].
По данным И.Б. Манухина, Л.Г. Тумиловича, М.А. Геворкяна (2010) действие ЛГ повышает концентрации эстрадиола и пролактина. Физиологическое снижение функции яичника вызывает уменьшение амплитуды и частоты секреторных импульсов, что обеспечивает экспансивное увеличение ФСГ по сравнению с ЛГ и запускает развитие очередного пула антральных фолликулов. Ученые сообщают, что ингибин снижает секрецию ФСГ; активины напротив потенцируют синтез ФСГ, пролиферацию клеток гранулёзы, трансгенез андрогенов в эстрогены; инсулиноподобные факторы роста I и II (ИФР-I, ИФР-II) вызывают активизацию ЛГ-индуцированного синтеза андрогенов в клетках теки, ароматизации андрогенов в эстрогены, ФСГ-индуцированного синтеза рецепторов к ЛГ на поверхности клеток гранулёзы. Сосудистый эндотелиальный фактор роста имеет важное значение в ангиогенезе растущих фолликулов, создаёт в доминантном фолликуле повышенные концентрации ФСГ. Эпидермальный фактор роста является наиболее важным стимулятором клеточной субстрации, трансформирующий фактор роста вызывает клеточную пролиферацию, оказывает миогенный и онкогенный эффекты [10].
В.Н. Бабичев (1998) указывает на то, что к центральной регуляции гипоталамо-гипофизарной системы имеют отношение нейропептиды. Особая роль отведена синаптическим нейропептидам: дофамин, норадреналин, серотонин, семейство опиоидных пептидов. Они регулируют гипоталамическую секрецию ГнРГ [1]. Дофамин поддерживает цирхоральную секрецию ГнРГ, серотонин оказывает опосредованное тормозящее влияние на циклический выброс ГнРГ. По данным В.А. Бурлева (1998), опиоидные пептиды увеличивают секрецию пролактина и гормона роста и напротив, блокируют секрецию ЛГ, ФСГ и тиреотропного гормона, опосредованно модулируют секрецию гонадотропинов через катехоламинергические нейромедиаторы гипоталамуса [5].
Центральная нервная система содержит большое количество рецепторов к эстрадиолу, что указывает на важную роль данного гормона в реализации механизмов обратной связи, а также в нейромедиаторном обмене.
К.Ю. Боярским (1998) было установлено влияние факторов роста (инсулиноподобного, эпидермального, трансформирующего, сосудистого эндотелиального, ингибинов и активинов)
в механизмах ауто- и паракринной регуляции функции гонадальной системы [4].
По сведениям В.Н. Бабичева, обязательным условием формирования феноменов стадии возбуждения полового цикла являются определённые эстрогено-прогестероновое и андрогено-прогестероновое соотношения [1]. Учёный, проанализировав динамику половых стероидов в крови в течение полового цикла, показал, что повышение уровня в крови андрогенов и эстрогенов вызывает формирование стадии возбуждения и её феноменов только при второй волне роста фолликулов на фоне третьей стадии развития желтого тела и снижения уровня прогестерона. При этом пик концентрации эстрогенов вызывает ярко выраженную течку и половое возбуждение, тестостерона - половую охоту.
Также некоторые ученые отдельно рассматривают роль факторов внешней среды, на полноценное проявление всего комплекса описываемых признаков. В этой связи С.А. Нефедовой и соавторов, ведутся исследования регуляции компенсаторноприспособительных реакций сельскохозяйственных животных [8].
Целью работы стало изучение механизма эндокринной регуляции полового цикла и нормализация репродуктивной функции у телок голштинской породы. Исходя из этого, были поставлены задачи, в рамах которых проведены исследования по выявлению эффективных программ синхронизации эструса и овуляции у телок голштинской породы. В задачи исследований входило: изучить физиологические аспекты регуляции половой цикличности у крупного рогатого скота голштинской породы; проанализировать схему применения препарата «PGF Вейкс форте», используемого для стимуляции и синхронизации половой охоты; установить влияние железо-йод-селен содержащего препарата «Седимин» на репродуктивную функцию телок случного возраста; обосновать экономическую целесообразность применения препарата
«PGF Вейкс форте» на случных телках старше 16 месяцев.
Материал и методы исследований. Научно-исследовательскую работу проводили в 2019 году в ФГБОУ ВО РГА-ТУ, производственные испытания – в условиях молочно-товарной фермы ООО
«Вакинское Агро» Рыбновского района Рязанской области. Для проведения опыта, согласно схеме (Таблица 1), были отобраны телки голштинской породы случного возраста (16-18 месяцев), живой массой 380-430 кг оставшиеся яловыми. В каждой группе по 20 голов.
Таблица 1 – Схема исследований
|
Группа |
Количество голов |
Половозрастная группа |
Схема обработки |
|
Опытная I |
20 |
телки |
схема № 1 |
|
Опытная II |
20 |
телки |
схема № 2 |
|
Опытная III |
20 |
телки |
схема № 3 |
|
Контрольная IV |
20 |
телки |
без обработки |
Формирование опытных групп происходило с учетом принципа ограниченной рендомизации. При этом использовали метод «сбалансированных групп-аналогов» [16]. Кормление животных осуществляли по нормам Всероссийского НИИ животноводства.
Выявление телок «в охоте» проводили визуально – в случае осеменения в естественную охоту. Особое внимание в этих случаях обращали на «впрыгивания» животных, «рефлекс неподвижности», истечение слизи из половой щели, её цвет, консистенцию, прозрачность. При ректальной диагностике обращали внимание на состояние стенки матки, ригидность. Используя УЗИ-сканер, устанавливали наличие или отсутствие доминантного фолликула, способного овулировать. Искусственное осеменение животных проводили фронтально в установленное время, за одну охоту однократно. При этом использовали замороженно-оттаянную сперму в дозе 0,25 мл, содержащей 15 млн. спермиев с активным поступательным движением. Применяли ректо-цервикальный способ осеменения с использованием системы фиксации «Head Lock» (Россия). Для проведения УЗИ использовали ультразвуковой сканер EasiScan-3 («BCF Technology», Великобритания) с линейным зондом 7,5 МГц.
Первая группа телок была обработана по следующей схеме № 1:
1 сутки (8:00-11:00) – ректальное исследование на выявление врождённой патологии половых органов и введение внутримышечно (в/м) 2 мл «PGF Вейкс форте» и 10 мл «Седимин». 14 сутки (8:0011:00 ч) – 2 мл в/м «PGF Вейкс форте». 17 сутки (8:00-11:00 ч) – искусственное осеменение (ИО). 56 сутки – ректальное исследование на стельность при помощи УЗИ-сканера. За основу взята система двухразовых инъекций.
Вторую группу телок обработали по схеме № 2:
1 сутки (8:00-11:00) – ректальное исследование на выявление врождённой патологии половых органов и введение в/м 2 мл «PGF Вейкс форте». 14 сутки (8:0011:00 ч) – 2 мл в/м «PGF Вейкс форте». 17 сутки (8:00-11:00 ч) – ИО. 56 сутки – ректальное исследование на стельность при помощи УЗИ-сканера. За основу взята система двухразовых инъекций.
Третья группа телок была обработана по следующей схеме № 3:
-
1 сутки – ректальное исследование на выявление врождённой патологии половых органов и введение в/м 10 мл «Се-димин». 1-30 сутки – выявление животных в охоте и проведение искусственного осеменения.
Обработку животных по предложенным схемам проводили в каждой группе однократно. Для контроля в IV группу были включены животные, которым стимуляцию половой охоты не проводили и не применяли препарат «Седимин». В данной группе осеменение проводили однократно при выявлении феноменов стадии возбуждения. Телок первой и второй опытных групп осеменяли в установленное время, не обращая внимания на признаки охоты.
Статистическую обработку результатов проводили с помощью программы Microsoft Excel.
Результат исследований. После синхронизации половой охоты, проведения искусственного осеменения получены результаты (Рисунок 1, Таблица 2), из которых следует, что с первого раза стельными стали не все животные, подвергнутые обработкам.
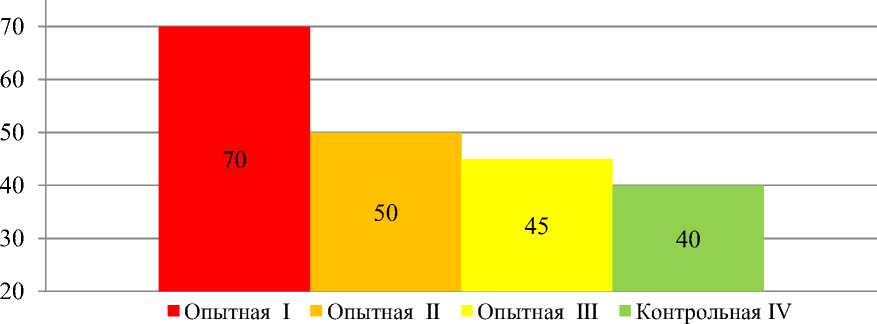
Рисунок 1 – Средний процент стельных животных в исследуемых группах
Таблица 2 – Итоги синхронизации эструса у телок
|
Показатель |
Группа |
||||
|
I |
II |
III |
IV |
||
|
Количество, голов |
20 |
20 |
20 |
20 |
|
|
Обработано, голов |
20 |
20 |
20 |
20 |
|
|
Осеменено, голов |
20 |
20 |
20 |
20 |
|
|
Оплодотворилось, голов |
14 |
10 |
9 |
8 |
|
|
Осеменено повторно, голов |
6 |
10 |
11 |
12 |
|
|
Таблица 3 – Рыночная стоимость препа |
ратов |
||||
|
Наименование препарата |
Количество |
Цена, р. |
Стоимость, р. |
||
|
PGF Вейкс форте 50 мл (флакон) |
4 |
2128 |
8512 |
||
|
Седимин 100 мл (флакон) |
4 |
120 |
480 |
||
Оптимальные результаты были получены после обработки по схеме № 1. В этой группе нестельными оказались шесть голов (30 %) от обработанных, что на 30 % эффективнее, чем в группе контроля, на 20 % по сравнению со второй группой и на 25 % – с третьей. Во второй группе нестельными остались десять голов (50 %), что на 10 % результативнее, чем в группе контроля, и на 5 % выше, чем в третьей группе, в которой оплодотворилось девять голов (45 %), что на 5 % больше, чем в группе контроля.
В четвертой группе получено стельных – восемь голов (40 %). Все нестельные животные были осеменены по- вторно. После проведения двух этапов стимуляции, безрезультативно осеменённые животные выбраковывались.
Из-за разных методических подходов к организации случного периода, неравномерного прихода животных в охоту, неодинаковой оплодотворяемости телок из опытных групп, различного количества затраченных спермодоз появилась необходимость установить дифференцируемые экономические показатели эффективности воспроизводства стада ремонтных телок, что и было сделано (Таблица 3, 4).
Во всех группах было затрачено одинаковое количество спермодоз – 20.
При этом результативность осеменения была ниже в необработанной группе. Соответственно возрос индекс осеменения в контрольной группе. Лучшей по этому показателю оказалась группа телок, обработанная по схеме № 1. Здесь индекс осеменения составил 1,42 ед., что на 1,08 ед. (на 43,2 %) меньше, чем в контрольной группе. Во второй группе индекс осеменения составил 2,0 ед., что на 0,5 ед. (на 20,0 %) меньше, чем в контрольной группе телок. При обработке по схеме № 3 индекс осеменения составил 2,22 ед., что на 0,28 ед. (на 11,2 %) меньше, чем в группе необработанных животных.
При обработке по схемам № 1 и №
-
2 продолжительность случного периода до искусственного осеменения составила 17 дней, что на 13 дней меньше, чем в третьей и контрольной группах, в которых аналогичный период продлился 30 дней.
Показатель «прогнозируемый выход живых телят» рассчитывали с учётом внутрихозяйственных данных. Так, выход живых телят в ООО «Вакинское Агро» на 100 нетелей с учётом двоен 90±5 %.
Среди обработанных телок затраты на одного телёнка были меньшими при реализации схемы № 1. Реализация схем стимуляции и синхронизации позволяет получить дополнительных телят.
Таблица 4 – Экономические показатели эффективности воспроизводства
|
Показатель |
Группа |
|||
|
I |
II |
III |
IV |
|
|
Затрачено доз семени |
20 |
20 |
20 |
20 |
|
Стоимость 1 дозы семени, р. |
550 |
550 |
550 |
550 |
|
Затраты на семя, тыс. р. |
11,0 |
11,0 |
11,0 |
11,0 |
|
Количество стельных нетелей |
14 |
10 |
9 |
8 |
|
Прогнозируемый выход живых телят |
13 |
9 |
8 |
7 |
|
Стоимость одного телёнка, тыс. р. |
7,2 |
7,2 |
7,2 |
7,2 |
|
Стоимость всех телят, тыс. р. |
93,6 |
64,8 |
57,6 |
50,4 |
|
Стоимость обработки 1 головы, р. |
97,12 |
85,12 |
12,0 |
0 |
|
Количество обработок |
20 |
20 |
20 |
0 |
|
Затраты на 1 обработку, р. |
1942,4 |
1702,4 |
240 |
0 |
|
Затраты на семя и обработку, тыс. р. |
12,942 |
12,702 |
11,24 |
11,0 |
|
Затраты на 1 телёнка, р. |
995,5 |
1411,4 |
1405,0 |
1571,4 |
|
Индекс осеменения, ед. |
1,42 |
2,0 |
2,22 |
2,5 |
|
Дополнительно будет получено телят |
6 |
2 |
1 |
0 |
|
Стоимость дополнительно полученных телят, тыс. р. |
43,2 |
14,4 |
7,2 |
- |
|
Затраты на дополнительно полученных телят, р. |
5973 |
2822,8 |
1405 |
- |
|
Дополнительно будет получено за телят, тыс. р. |
37,227 |
11,577 |
5,795 |
- |
|
Продолжительность реализации схемы до ИО, дней |
17 |
17 |
30 |
30 |
Заключение. Таким образом, наиболее эффективным способом стимуляции и синхронизации эструса у телок голштинской породы оказалась модифицированная система двухразовых инъекций с интервалом между инъекциями 14 дней, дополненная железо-йод-селен содержащим препаратом «Седимин». Так при реализации данной схемы количество стельных животных составило 70 %. Препарат «PGF Вейкс форте» гарантировано стимулирует и синхронизирует половую охоту, позволяя осеменять в установленное время. Препарат «Седимин» положительно влияет на репродуктивную функцию телок. При его применении в первой группе телок дополнительно получено 20 % стельных животных по сравнению со второй группой, где данный препарат не применяли. Среди опытных групп телок при реализации схемы № 1 затраты на одного телёнка были меньшими – 995,5 р., дополнительно будет получено за реализацию телят – 37227 р. В этой группе был ниже индекс осеменения – 1,42 по сравнению с другими группами. Необходимо профилактировать недостаток в организме микроэлементов, определяющих состояние репродуктивной функции путём применения комплексного препарата «Седимин».
Резюме
В крупном молочном предприятии Рязанской области проведены исследования механизма эндокринной регуляции эстрального цикла, а также изучены пути нормализации репродуктивной функции. Экспериментальные данные получены на телках голштинской породы случного возраста. В сравнительном аспекте оценена экономическая эффективность применяемых методик. Используемые препараты и методы качественно отличались друг от друга. Впервые доказано сочетанное применение препаратов «PGF Вейкс форте» и «Седимин», позволяющее в среднем на 30 % увеличить показатель вероятности наступления беременности у телок случного возраста.
Список литературы Механизм эндокринной регуляции полового цикла и нормализация репродуктивной функции у телок голштинской породы
- Бабичев, В.Н. Нейроэндокрино-логия репродуктивной системы [Текст] / B.Н. Бабичев // Проблемы эндокринологии. - 1998. - № 1. - С. 3-12.
- Белобороденко, А.М. Воспроизводство и профилактика бесплодия коров в условиях Северного Зауралья [Текст] / А.М. Белобороденко, М.А. Белобороденко, Т.А. Белобороденко // Вестник государственного аграрного университета Северного Зауралья. - 2013. - № 3. - С. 58-61.
- Белобороденко, М.А. Профилактика скрытых абортов у коров [Текст] / М.А. Белобороденко [и др.] // Генетика и разведение животных. - 2016. - № 4. - C. 52-56.
- Боярский, К.Ю. Функциональные тесты, определяющие овариальный резерв, и вспомогательные репродуктивные технологии [Текст] / К.Ю. Боярский, [и др.] // Проблемы репродукции - 1998. - № 3. - С. 26-31.
- Бурлев, В.А. Факторы роста и их роль в регуляции репродуктивной функции у больных с синдромом поликистоз-ных яичников [Текст] / В.А. Бурлев, [и др.] // Проблемы репродукции. - 1998. - № 3. -С. 17-25.
- Дедов, И.И. Синдром полики-стозных яичников [Текст] / И.И. Дедов, Г.А. Мельниченко. - М.: Изд. «МИА», 2007. - 361 с.
- Дюльгер, Г.П. Дифференциальная гормонотерапия коров с фолликулярными и лютеиновыми кистами яичников под ультразвуковым контролем [Текст] / Г.П. Дюльгер // Ветеринария сельскохозяйственных животных. - 2010. - № 6. -С. 32-42.
- Коровушкин, А.А. Регуляция потребления корма, роста, развития, продуктивности коров с гипофункцией щитовидной железы в зависимости от индукции компенсаторно-приспособительных реакций Са2+-антагонистом / А. А. Коровушкин, В. С. Грачев, С. А. Нефедова // Известия Санкт-Петербургского государственного аграрного университета. - 2011 - № 22. - С.130-135.
- Манухин, И.Б. Ановуляция и ин-сулинорезистентность / И.Б. Манухин, М.А. Геворкян, Н.Б. Чагай // М.: Изд. «Геотар-Медиа». - 2006. - 415 с.
- Манухин, И.Б. Гинекологическая эндокринология. Клинические лекции [Текст] / И.Б. Манухин, Л.Г. Тумилович, М.А. Геворкян. - М.: Изд. «Гэотар-Медиа», 2010. - 275 с.
- Овсянников, А.И. Основы опытного дела в животноводстве [Текст] / А.И. Овсяников. - М., Колос, 1976. -304 с.
- Племяшов, К.В. Воспроизводительная функция у высокопродуктивных коров при нарушении обмена веществ и её коррекция / К.В. Племяшков // Автореф. дис. док. вет. наук: 06.02.06 - ветеринарное акушерство и биотехника репродукции животных. - СПб, 2010. - 41 с.
- Сердюк, Г.Н. Проблема продуктивного долголетия при голштинизации отечественных пород крупного рогатого скота и пути её решения / Г. Н. Сердюк // Молочное и мясное скотоводство. - 2015. - № 6. - С. 7-9.
- Туников, Г.М. Современные тенденции производства молока в условиях интенсивной технологии [Текст] / Г.М. Туников, А.А. Коровушкин [и др.] // Вестник Рязанского государственного аг-ротехнологического университета им. П. А. Костычева. - 2019. - № 4 (44). - С. 70-75.
- Patel, H. Follicle stimulating hormone modulates ovarian stem cells through alternately spliced receptor variant FSH-R3 / H. Patel, D. Bhartiya, S. Parte [et al.] // Ovarian Res. - 2013. - Vol. 6. - P. 52.
