Mesolithic human teeth from Zamostye-2, Moscow region

Author: Zubova A.V., Moiseev V.G., Kulkov A.M.
Journal: Archaeology, Ethnology & Anthropology of Eurasia @journal-aeae-en
Section: The metal ages and medieval period
Article in issue: 4 т.47, 2019.
Free access
Short address: https://sciup.org/145145454
IDR: 145145454 | DOI: 10.17746/1563-0110.2019.47.4.120-127
Text of the article Mesolithic human teeth from Zamostye-2, Moscow region
The Zamostye-2 site is located in the north of the Sergievo-Posadsky District of the Moscow Region, on the left bank and in the bed of the Dubna River. The site was first discovered in 1987; and since 1989, full-scale excavations have been carried out there by V.M. Lozovsky and O.V. Lozovskaya. With some short breaks, the excavations lasted until 2001, when they were temporarily stopped in order to study the excavated material, and were then resumed in 2009–2013 (Lozovsky, Lozovskaya, 2013: 6–8).
The cultural layers of the site lie in lake-bog deposits at a depth of 2–4 m from the ground’s surface, and can be described as a consequence of organogenic sapropelic deposits saturated by peat and macroremains. Several cultural layers belonging to the Late Mesolithic, Early and Middle Neolithic were detected at the site. The lower Late
Mesolithic layer is dated to between 7000 and 6600 BC, the upper layer ca 6400–6000 BC, and the final Mesolithic layer ca 6000–5800 BC. The Early Neolithic horizon consists of the remains of a dwelling area, dated to 5800– 5200 BC on the basis of the site’s materials, and belonging to the Upper Volga culture. The Middle Neolithic layer belongs to the Lyalovo culture and is dated to 4900– 4300 BC (Lozovsky et al., 2013: 18).
A small sample of human bone remains was collected at the site. The sample can be divided into two categories: deciduous teeth that were lost in a natural way and persisted in the layer of the dwelling area; and isolated reburied fragments of skulls and postcranial bones from adult. As no intentional burials were found at the site, it can be suggested that somewhere nearby there was a burial ground blurred by the river, and the bone fragments were brought by water during seasonal flooding of the camp.
While the Mesolithic remains do not constitute a representative sample, they are of great interest, since no Mesolithic skeletal remains have previously been found in this area. The main purposes of this study are a comprehensive morphological description of the sample, a preliminary reconstruction of their possible population affinities, and a reconstruction of the general diet of the Mesolithic population that inhabited the Zamostye-2 site.
Material and methods
The Mesolithic dental sample from Zamostye-2 includes a fragment of the mandible of an adult individual, a fragment of a maxilla of an adult individual (both with teeth preserved), a fragment of a subadult mandible with teeth in situ , and four isolated deciduous teeth. The number of individuals to whom the remains might have belonged is from 4 to 7. The bone specimens probably belonged to three individuals: a young female, an adult individual of unidentified sex, and a subadult 1.5–2 years old. The isolated deciduous teeth were lost naturally and at different ages; thus they could have belonged to one or several children.
All the teeth were studied using several protocols for describing crown non-metric traits (Zubov, 2006; Zubov, Khaldeeva, 1993; Edgar, 2017). Main diameters of the crowns and roots of the teeth were measured as well. The second deciduous molar of the subadult’s mandible was microCT scanned using Skyscan-1172. The scanning protocol was set as follows: tube voltage 100 kV, current strength 100 μA (no filter), rotation step 0.25°, averaging over three frames, at the resolution of 3.45 μm/pixel. Reconstruction of the raw images was carried out in the NRecon software (Bruker-microCT); a 3D model of the tooth and separation of the dentine from enamel were done using the CTAn software (Bruker-microCT). For visualization of the digital model we used the CTVox software (Bruker-microCT). Morphology of the dentineenamel junction was described using the 3D model, maximum thickness of the lateral enamel level of the metaconid was measured, as well as the volume of lateral dentine and the pulp chamber (LDPV). Enamel thickness was measured perpendicularly to the vertical axis of the lingual surface of the metaconid. For this, a buccolingual section was done along the line connecting apexes of the protoconid and the metaconid. The LDPV was measured following a standard protocol (Benazzi et al., 2011, Toussaint et al., 2010) in CTAn.
Mesiodistal and buccolingual diameters of the upper first and lower second molars of the adult individuals were further compared to measurements of Mesolithic and Upper Paleolithic dental specimens (from an opensource repository) from Germany, France, Sweden, Denmark, Italy, Portugal, and Serbia (Voisin et al., 2012).
The specimens were also compared to the sample from the Mesolithic burial ground of the Onega culture at Yuzhny Oleny Island, and to some Upper Paleolithic finds from European Russia (unpublished data of A.V. Zubova). As it was not possible to identify sex from a mandible, comparative analyses were carried out on sex-combined samples.
Besides the morphological study, a paleopathological description of the teeth was performed, in order to reconstruct the diet of the population under study. This included scoring caries lesions, enamel hypoplasia (LEH), periodontal disease, ante-mortem tooth loss, dental calculus, and ante-mortem enamel chipping.
The frequency of carious lesions is traditionally considered the main indicator of diet composition. The lesions emerge as a result of enamel demineralization triggered by bacterial fermentation of carbohydrates from food, which is accelerated by consumption of soft and viscous food items (Lillie, 1996; Keenleyside, 2008; Larsen, Shavit, Griffin, 1991). Another important marker, dental calculus, is a result of mineralization of the bacterial plaques attached to enamel’s surface. There is no strict correlation found between diet composition and the development of dental calculus, but a number of studies have shown that a higher prevalence of calculus in combination with a low frequencies of caries is observed in populations consuming more protein and less carbohydrates. However, in agriculturalists, a high prevalence of both pathologies is found.
The markers of disturbance of periodontal tissue supply were employed as a secondary marker, which can be related to the severity of dental calculus or a vitamin deficiency (Nazir, 2017; Strohm, Alt, 1998; Putten et al., 2009). Enamel hypoplasia was used as yet another secondary variable, which is viewed in the majority of bioarchaeological studies as a marker of biological stresses in childhood, or a lack of food recourses (Mednikova, 2017: 80).
Morphological description of the finds
In the sample from Zamostye-2, specimens No. 8, 9, 14, 17, 18, 20, and 21* can be assigned to Mesolithic times.
Specimen No. 8 (layer 7, square B3; date according to the layer 6500–6000 BC, date of the bone 7663 ± 44 BP (KIA-51435)). This is the left half of the mandible of a young female (18–20 years). Alveoli of the first and second premolars, and third molar are preserved; the first and second molars were in situ (Fig. 1). The crown of the first molar is destroyed. The second tooth is four- cusped, crown pattern is “Y”, cervical enamel extension is grade 6, protostylid is grade 1. Tami, t6, distal and middle trigonid crests, deflecting wrinkle of the metaconid, posterior fovea, and central cusp are all absent. The odontoglyphic pattern displays intertubercular fissures I–VI and grooves 1 and 2med, 1 and 2prd, 1 and 2end. The hypoconid is heavily abraded, and its odontoglyphic pattern is not visible. An anterior fovea is observed, odontoglyphic variant is 2med(II).
The roots of the surviving teeth protrude from the alveoli for 3.5–4.0 mm on average, which is in general not typical for such a young age. But no traces of inflammation were observed on the fragments of the alveolar margin, excluding a small porotic area in the interdental space. Deposits of dental calculus were detected on the preserved part of the crown of the first molar. This preserved part is adjacent to the mesial plane of the second molar. Calculus is observed in all planes of the lower third of the crown. There is also a carious lesion in fissure III of this tooth.
Specimen No. 9 (ditch; date according to the layer 7000–6000 BC). This is a fragment of the right half of the maxilla of an individual 20–30 years old. The alveolus of the first incisor is preserved; the second incisor, canine tooth, premolars, and first molar are in situ (Fig. 2). The lateral incisor displays an absence of both lingual and labial shoveling, a weak development of the lingual cusp, and the presence of finger-like projections. The canine exhibits very weakly pronounced marginal ridges on the lingual surface (shoveling, grade 0–1), as well as tuberculum dentale and distal accessory ridge. The labial cusp of the first molar is only slightly larger than the lingual one, while the two cusps of the second molar are of equal size. The hypocone of the first molar is not reduced, while its metacone shows an initial stage of reduction. The crista oblique is interrupted. A weakly pronounced Carabelli cusp (grade 2), distal and mesial accessory cusps are present. The odontoglyphic pattern of the crown is not traceable, owing to dental wear; the anterior and posterior fovea and cingular derivatives are absent. Cervical enamel extension is grade 5.
The roots of the present teeth protrude from the alveoli for 3–4 mm, on average. Some porosity of the alveolar margin is observed, which suggests a disturbed supply of the periodontal tissues. Enamel hypoplasia is present on the labial surface of the canine and incisor. This is also observed in the lower third of the crown of first molar, where it surrounds the whole crown. Dental calculus is present on labial surface of all the teeth and in the interdental space. A caries lesion was detected near the central fovea of the first molar. Ante-mortem vertically oriented enamel defects (chipping) are present on the incisor, the canine, and the first premolar.
Specimen No. 14 (layer 1, square B1). This is the mandible of a subadult of 1.5–2 years of age (Fig. 3).

Fig. 1. Mandible, specimen No. 8.

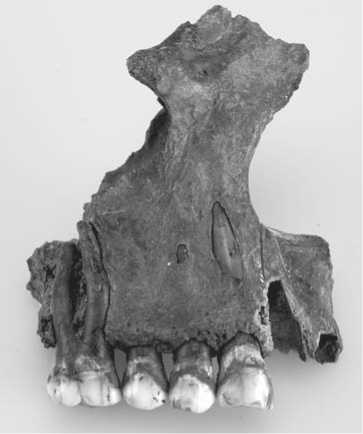
0 3 cm
Fig. 2. Maxilla, specimen No. 9.
Fig. 3. Subadult mandible, specimen No. 14.
Erupting deciduous teeth (both first molars and the left second molar) were found in situ . The crowns of the first molars are five-cusped, with a “Y” pattern. The epicristid connecting the cusps of the trigonid is interrupted by intertubercular fissure II. The odontoglyphic pattern includes deep intertubercular fissures I–V and the grooves delimiting axial ridges of all cusps except hypoconulid. Fissures I and V reach the labial surface.
The second left molar is five-cusped, but the odontoglyphic differentiation of its occlusal surface is significant. The crown exhibits an X5 type of contact. The odontoglyphic pattern includes intertubercular fissures I–V and grooves 1 and 2 of all the five cusps. Duplicate grooves 2’med and 2’prd are observed in the metaconid and protoconid, while 3end is present on the entoconid. The distal trigonid crest, epicristid, deflecting wrinkle of the metaconid, protostylid, cebtarl cusp, and posterior fovea are absent. A very weakly developed fovea of the protostylid, an anterior fovea, a 2med (fc) variant, and type 2 of superposition of contact points of the first grooves of the metaconid and protoconid with fissure II are present.
The morphology of the enamel dentine junction completely corresponds to the appearance of external enamel of the crown. The maximum thickness of lateral enamel of the metaconid is 1.037 mm, while the volume of lateral dentine and pulp chamber is 151 mm3.
Specimen No. 17 (layer 7, square A12; date according to the layer 6500–6000 BC). This is the crown of an upper right second deciduous molar of a subadult 8–10 years old. The strong enamel abrasion suggests that an age of 10 years is most plausible. The preserved fragment has three roots: two labial and one lingual. The hypocone of the tooth is markedly reduced, and the metacone does not display any sign of reduction. The odontoglyphic pattern is abraded. Dental calculus is observed on the interproximal surfaces, the hypocone displays antemortem enamel chipping.
Specimen No. 18 (layer 7, square B12; date according to the layer 6500–6000 BC). This is the crown of an upper right lateral deciduous incisor of a subadult individual of 6–7 years of age. The preserved part of the tooth is an adjacent fragment of the cervical portion of the root system, about 2.5 mm in length. The tooth is moderately worn; some dental calculus is observed on the interdental surfaces and on the labial side. Lingual or labial shoveling, lingual inclination of the crown, and accessory ridges are all absent.
Specimen No. 20 (layer 7, square A11; date according to the layer 6500–6000 BC). This is the upper left first deciduous incisor of a subadult ca 5–6 years of age. A marked lingual cusp is present, while finger-like projections, as well as lingual or labial shoveling, are absent. Several very small enamel defects are observed on the cutting edge of the tooth. Focal hypoplasia in the form of a vertical strip of malformed enamel is present (3.1 mm long and 1.6 mm wide).
Specimen No. 21 (layer 7, square A11; date according to the layer 6500–6000 BC). This is the lower right second deciduous incisor of a subadult ca 5–6 years of age. Dental calculus is found on the mesial and distal interproximal surfaces. Several small enamel chippings are observed on the labial surface of the cutting edge.
Discussion and conclusions
Reconstruction of diet. Both individuals represented by permanent teeth display carious lesions in the fissures. A high prevalence of this pathology is typically considered a marker of an agricultural subsistence strategy. It is believed that caries is very rare or absent in populations of hunters and gatherers (Murphy et al., 2013: 2554; Turner, 1979: 623, tabl. 2). This view is largely supported by world-wide data, but there are a number of exceptions. For instance, increased prevalence of caries was described for the Early Bronze Age hunter-gatherers of Western Siberia. According to isotopic data, these people consistently consumed local wild plants with the C3 type of photosynthesis (Marchenko et al., 2015: Tabl. 4; Marchenko et al., 2016: 173; Zubova, Marchenko, Grishin, 2016: Tabl. 1). To date, there has been no archaeological evidence of any type of agricultural activities for the Mesolithic population of the Upper Volga lowlands. This suggests that the high prevalence of caries in the ancient population from Zamostye-2, like in Western Siberia, is related to a general diversity of diet, which included not only fish or meat, but also local plants. This is confirmed by the results of archaeobotanical studies showing the long-standing tradition of consumption of local fruit and berries by the inhabitants of the camp (Beriuete, 2018: 47), as well as by the results of the isotopic analysis (Meadows, Lozovskaya, Moiseev, 2018). The vertical enamel chippings found in the incisors, canines, and premolars might be due to gnawing of small bones or nuts (Lee et al., 2011: 971).
Population history of the group. The set of nonmetric dental traits observed in the permanent dentition is generally not diagnostic. The only conclusion that can be drawn is the absence of “Eastern” markers in the upper lateral incisor and lower second molar. The fingerlike projections detected in the upper incisor and canine, and the distal accessory ridge observed in the canine, suggest somehow “archaic” (on the modern human scale) morphology of the teeth.
The lower second deciduous molar is the key tooth for both deciduous and permanent molar row (Farmer, Townsend, 1993; Bockmann, Hughes, Townsend, 2010). In our case (specimen No. 14), it shows a combination of taxonomically neutral (five cusps, absence of the
“Eastern” markers) and “archaic” (complexity of the odontoglyphic pattern, presence of the anterior fovea) features. On the other hand, according to the micro-CT results, the proportions of the internal tissues of the crown are fairly progressive, and match up completely to the features of modern European populations (Benazzi et al., 2011: Tabl. 3). The tooth displays the maximum possible value of the ratio between the largest thickness of lateral enamel of the metaconid and the volume of lateral dentine and the pulp chamber. This ratio tends to increase in the course of human evolution, as enamel thickness increases while dentine volume decreases in more progressive taxa as compared to more archaic ones (Ibid.: 325). These features sharply distinguish this molar from the Upper Paleolithic specimens studied previously. The tooth displays, rather, similarity to the Mesolithic specimen
Table 1. Proportions of inner tissues of the crowns of the deciduous second molars of the mandible from Zamostye-2 and reference samples
|
Site |
LDPV |
MaxETH/ LDPV |
|
Kostenki-14 (Paleolithic) |
192 |
0.47 |
|
Yudinovo (Paleolithic) |
171.05 |
0.60 |
|
Yuzhny Oleny Island (Mesolithic) |
147.5 |
0.63 |
|
Zamostye-2 (Mesolithic) |
151 |
0.68 |
Note . LDPV – lateral dentine plus pulp volume; MaxETH – maximum thickness of metaconid enamel.
from the Onega culture burial ground of Yuzhny Oleny Island (Table 1).
The metric variables of the permanent teeth (Table 2) appeared to be more informative in terms of exploring the population history of the studied Zamostye group. The bivariant plots for mesiodistal and buccolingual diameters of the first upper molar and second lower molar clearly demonstrate that the closest analogs to the teeth from Zamostye-2 are found in the sample from the Yuzhny Oleny Island burial ground. This latter sample shows much smaller sizes of molars than the majority of other European finds (Fig. 4, 5).
The summary difference between the Yuzhny Oleny Island sample and the Mesolithic and Paleolithic European population is statistically significant for three out of four tooth dimensions used in our intergroup analysis (Table 3). This observation is in perfect agreement with genetic studies suggesting different origins of Eastern vs. Western European hunter-gatherers, where the former include the group from Yuzhny Oleny Island and the Mesolithic and Neolithic population from Samara region of the Volga (Mathieson et al., 2015).
In order to obtain an integral picture of the differences between the Mesolithic and Upper Paleolithic European samples, a canonical discriminate analysis employing mesiodistal and buccolingual diameters of the upper first and the lower second molars was carried out. Since both permanent molars from Zamostye-2 show a similar range of analogs, their characteristics were summarized. As is seen in Fig. 6, three clusters are clearly differentiated in the space of first two canonical vectors (CV). The first
Table 2. Dimensions of the dental specimens from the Mesolithic layers of the Zamostye-2 site
|
Number of the specimen |
Tooth |
Mesiodistal diameter |
Buccolingual diameter |
Height of the crown (min) |
Length of the root system (min) |
|
Maxilla |
|||||
|
9 |
I2 |
7 |
6.7 |
9.3 |
|
|
C |
7.9 |
8.9 |
21 |
||
|
Р1 |
6.6 |
8.7 |
7.9 |
||
|
Р2 |
6.6 |
9.3 |
7.5 |
||
|
M1 |
11 |
11.3 |
7.2 |
18.8 |
|
|
17 |
m2 |
9.6 |
|||
|
18 |
i 2 |
5.4 |
4.7 |
6 |
|
|
Mandible |
|||||
|
8 |
М 2 |
11.2 |
10 |
6.7 |
|
|
14 |
m1 right |
9.2 |
8 |
7.2 |
|
|
m1 left |
9.4 |
8.1 |
7.2 |
||
|
m2 right |
10.2 |
8.6 |
... |
||
|
21 |
i2 right |
5.4 |
5.04 |
||
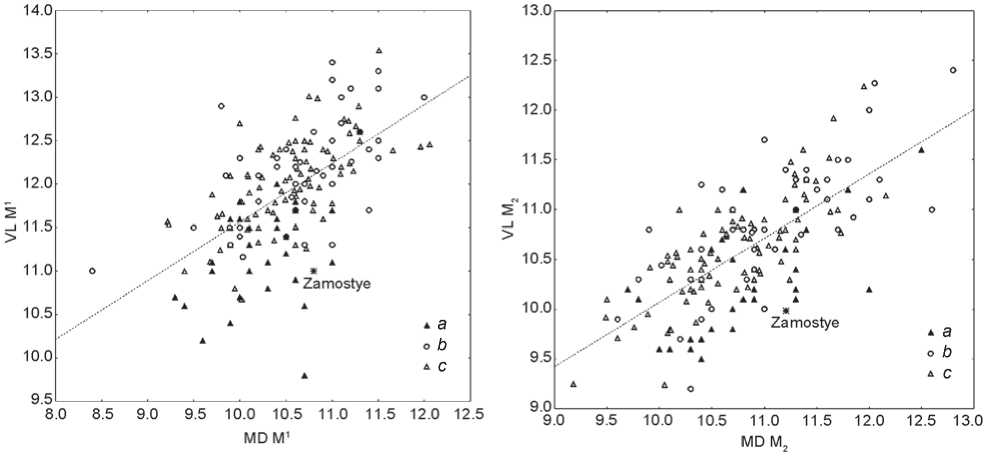
Fig. 4. Results of the comparison of metrics of the upper first molar from Zamostye-2 with European Upper Paleolithic and Mesolithic dental samples.
a – sample from Yuzhny Oleny Island; b – Upper Paleolithic specimens; c – Mesolithic specimens.
Fig. 5. Results of the comparison of metrics of the lower second molar from Zamostye-2 with European Upper Paleolithic and Mesolithic dental samples.
Legend same as on Fig. 4.
1.2
1.0
0.8
0.6
Mesolithic of Germany О
Table 3. Values of the t-test for comparison of dimensions of the upper first and lower second molars from Yuzhny Oleny Island (YOI) and the Paleolithic (PE) and Mesolithic (ME) European population
|
Parameter |
t-test results |
|
|
YOI and ME |
YOI and PE |
|
|
MD M1 |
2.48 * |
3.12 * |
|
VL M1 |
7.15 * |
6.95 * |
|
MD M 2 |
–1.18 |
1.42 |
|
VL M 2 |
2.81 * |
4.39 * |
Note . YOI – Yuzhny Oleny Island sample ( N = 29); МЕ – Mesolithic population of Northern, Western, and Southern Europe ( N = 84); PE – Paleolithic population of Western and Southern Europe ( N = 47).
* р < 0.05.
0.4
<3
0.2
Zamostye О
Yuzhny Oleny Island О
Mesolithic of France О
Mesolithic of Scandinavia О Upper Paleolithic of Western Europe
О
Upper Paleolithic of Eastern Europe
-3.0 -2.5 -2.0 -1.5 -1.0 -3.5 0 0.5 1.0
I Q
Fig. 6. Plot of the first two canonical vectors of the analysis of the Paleolithic and Mesolithic dental samples.
cluster includes the teeth from Zamostye-2 and Yuzhny Oleny Island. These specimens display negative values of CV I, which clearly differentiate them from other samples that all occupy the other pole of the vector axis. CV II separates these latter into two subgroups. The first includes both Upper Paleolithic samples and a combined Mesolithic sample from Denmark and Sweden, the second Mesolithic samples from France and Germany. Thus, the results of the multivariate analysis confirm the observation regarding a possible affinity between the populations from Zamostye-2 and Yuzhny Oleny Island made when considering the single dimensions of the teeth.
In our opinion, the similarity between the specimens from these two sites is of interest from the point of view of detailing the picture of affinities of the population of forest zone of Eastern Europe, drawn by the results of paleogenetic studies. As mentioned above, a considerable genetic similarity is observed between ancient populations of such remote areas as Karelia and the Samara region of the Volga. Our results based on the study of dental remains from Zamostye-2 make it possible to hypothesize tentatively that this group might also be related to the Eastern hunter-gatherers.
Conclusions
While the Mesolithic dental remains from Zamostye-2 cannot be treated as a proper population sample, their study revealed results important for reconstruction of the subsistence strategy and affinities of the population of the forest zone of Eastern Europe. The pathological changes of the dentition observed in the studied specimens suggest that these people had a diverse diet that included both animal and vegetable food. Non-metric variables of the deciduous and permanent teeth from Zamostye-2 display a number of “archaic” features, but their pattern in general is insufficient to trace population affinities. The results of the analysis of the metric characteristics of the permanent molars from Zamostye-2 suggest that the inhabitants of the site might have had common origins with the Onega culture population from Yuzhny Oleny Island. More generally, the studied dental sample shows similarities to a wide range of Eastern European populations collectively referred to in paleogenetics as Eastern hunter-gatherers.
Acknowledgements
This study was supported by the Program for Basic Research of the Russian Academy of Sciences “Monuments of Material and Spiritual Culture in the Modern Information Environment”, Project “Morphometric Analysis of the Upper Paleolithic Paleoanthropological Collections of the MAE RAS with the Use of 3D Technologies and Databases”.
The authors express their gratitude to O.V. Lozovskaya for the opportunity to work with her data, and to Prof. E. Murphy for her consultations in the process of preparing this paper.
