Межпопуляционная изменчивость Orobanche cumana Wallr., поражающей подсолнечник в регионах юга России, выявляемая молекулярно-генетическими маркерами

Автор: Гучетль С.З., Челюстникова Т.А., Рамазанова С.А.
Статья в выпуске: 1 (146-147), 2011 года.
Бесплатный доступ
В целях изучения возможности молекулярно-генетической идентификации разных рас заразихи и характеристики её популяций нами были испытаны 48 праймеров для ПЦР. Использовали праймеры RAPD, SSR, ISSR, минисателлиты. Полиморфизм ДНК фрагментов, выявляемый данными праймерами, изучался на 38 индивидуальных образцах заразихи, представляющих следующие популяции: Выселковская, Привольненская, Светлоградская, Морозовская, Казанская, Копанская, Тацинская, Константиновская, Турецкая. Результаты исследования показали, что из 48 праймеров 16 не гибридизировались с ДНК заразихи. 24 праймера не выявили полиморфизма. Из оставшихся 8 праймеров 6 выявляли либо полиморфизм минорных фракций, либо не воспроизводимый. Лишь 2 RAPD праймера из 48 изученных выявили чёткий, воспроизводимый полиморфизм у разных образцов заразихи. Полученные результаты показывают, что молекулярно-генетический полиморфизм популяций заразихи на территории России можно оценить как слабый.
Заразиха, подсолнечник, пцр, днк-маркеры, полиморфизм
Короткий адрес: https://sciup.org/142150971
IDR: 142150971 | УДК: 633.854.78:582.952.6
Interpopulation variability of Orobanche cumana Wallr. affecting sunflower in south regions of Russia revealed by molekular-genetic markers
With a purpose to study a possibility of molekular-genetic identification of different broomrape races and to characterize its populations, we tested 48 primers for PCR. There were used primers RAPD, SSR, ISSR, minisatellites. Polymorphism of fragment's DNA revealed by data of the primers was studied on 38 individual broomrape samples, representing the following populations: Vyselkovsky, Privolnensky, Svetlogradsky, Morozovsky, Kazansky, Kopanskoy, Tatsinsky, Konstantinovsky, and Turkish. The results of a research showed that 16 primers from 48 ones were not hybridized with broomrape DNA. 24 primers haven't displayed polymorphism. Six primers from remained eight one revealed either polymorphism of minor fractions, or not reproduced one. Only two RAPD primers from 48 studied have revealed accurate and reproduced polymorphism in different broomrape samples. The obtained data showed that molecular-genetic polymorphism of broomrape populations on the territory of Russia can be estimated as weak.
Текст научной статьи Межпопуляционная изменчивость Orobanche cumana Wallr., поражающей подсолнечник в регионах юга России, выявляемая молекулярно-генетическими маркерами
Виды Orobanche – бесхлорофильные однолетние и многолетние растения, которые поражают корни различных видов растений. Паразитизм представителей рода Orobanche привел к упрощению их морфологии и, таким образом, к трудностям при идентификации некоторых видов [1]. В связи с этим исследователи все чаще используют методы молекулярной биологии, в частности полимеразную цепную реакцию с RAPD и другими праймерами как для более точного разделения видов рода Orobanche, так и для изучения внутривидового полиморфизма отдельных таксонов.
Roman с соавторами [2] с пользованием RAPD ПЦР была изучена вариабельность среди двадцати таксонов в роду Orobanche, собранных от пораженных заразихой растений в Испании. Авторы выявили потенциально видоспецифичные маркеры, которые могли быть использованы для обнаружения случаев естественной межвидовой генетической интро- грессии. Результатами кластерного анализа были четко разделены виды O. cumana Wallr. и O. cernua Loefl.
Эффективность использования RAPD маркеров для изучения пяти видов заразихи на сельскохозяйственных полях Израиля была показана Katzir c соавторами [3]. Были выявлены два маркера, которые можно использовать для идентификации различных секций ( Osproleon и Trionychon ) в роду Orobanche , и пять маркеров для идентификации разных видов, в том числе и O.cumana [3].
По шести RAPD праймерам были показаны различия между популяциями O. cumana и O . cernua , происходящими из разных районов Болгарии, России и Украины [4]. Benharrat с соавторами [5] исследовали геномный полиморфизм между отдельными видами, в том числе и близкородственными, O.cumana и O . cernua с помощью ПЦР с ISSR праймерами. Праймер GATA 4 обнаружил полиморфизм между пятью экземплярами каждого вида O. cumana и O . cernua из разных стран, позволяющий дифференцировать эти два вида.
Необходимо изучать популяционное разнообразие O. cumana , так как этот паразит поражает подсолнечник, являющийся важной масличной культурой во всем мире. За последние несколько лет наблюдается увеличение агрессивности заразихи подсолнечника, образование новых высоковирулентных рас и распространение на новые территории этого паразита [6; 7]. В литературе сообщается о существовании физиологических рас заразихи от А до Н [8]. Известно об обнаружении новых, не описанных ранее морфотипов [9].
Gagne с соавторами [10] определена генетическая изменчивость популяций O. cumana, происходящих из Болгарии, Испании, Румынии и Турции, где семенами заразихи засорены большие посевные площади. В качестве метода исследования использовали технику RAPD ПЦР с двадцатью тремя праймерами, показавшими воспроизводимость. Было показано, что у заразихи O. cumana процессы самоопыления преобладают над процессами переопыления. Популяции обладают малой внутрипопуляционной изменчивостью, и между индивидумами из различных географических областей происходит очень небольшой генный обмен. Молекулярное маркирование и последующий кластерный анализ показали, что изученные популяции заразихи из Восточной Европы и Испании отличаются друг от друга, хотя и минимально.
Cuica с соавторами [11] также использовали RAPD технологию для оценки генетической вариабельности десяти популяций заразихи O. cumana из Испании, Югославии и Румынии. ДНК-профили показали слабый полиморфизм между популяциями. Только румынские популяции из области Калараси отличались по двум праймерам, а из Барагани – по одному.
Генетическая изменчивость популяций заразихи, поражающей подсолнечник в России, практически не оценена по молекулярногенетическим маркерам. Поэтому целью нашей работы являлся поиск ДНК-маркеров для дифференциации известных рас заразихи и предварительная оценка молекулярно-генетического разнообразия популяций заразихи, собранных на разных генотипах подсолнечника в различных регионах юга России.
Материалы и методы. Материалом для исследования послужили тридцать восемь образцов из девяти популяций O. cumana . Семена заразихи были собраны в разные годы на полях подсолнечника Краснодарского, Ставропольского краёв и Ростовской области и хранились при –18 Со (табл. 1). Для анализа использовали клубеньки или верхушки соцветий заразихи, поразившей разные по устойчивости генотипы подсолнечника, выращенные в фитотронно-тепличном комплексе, в условиях, описанных ранее [12], или на искусственном инфекционном фоне из семян заразихи Светлоградской, Привольненской, Тацинской и Константиновской популяций [12].
ДНК выделяли модифицированным методом Zolan и Pukkila (1986) [13] из 4-5 индивидуальных образцов заразихи каждой популяции. Для ПЦР анализа использовали 48 праймеров: RAPD, SSR, ISSR, минисателлиты.
ПЦР выполняли в реакционной смеси (25 мкл) следующего состава: 67 мМ трис-HCl, рН 8,8; 16,6 мM сульфат аммония; 1,5-3,0 мM MgCl2; 0,01 % Tween 20; по 0,2 мM дезоксирибонуклеозидфосфатов; по 10 пМ праймеров; 10 нг матричной ДНК и 1 ед. рекомбинантной термостабильной ДНК-полимеразы («Госнииге-нетика», Россия). Амплификатор «Терцик» («АО ДНК-технология», Россия), режим ― начальная денатурация ДНК в течение 2 мин при 94 °С; 30-35 циклов с последовательной сменой температур: 60 с ― 94 °С (денатурация), 60 с – 3660 °С (в зависимости от праймера) (отжиг), 120 с ― 72 °С (элонгация); финальная элонгация – 4 мин при 72 ○С.
Таблица 1 – Популяции заразихи, использованные для анализа
|
Место сбора семян заразихи |
Год сбора |
Генотип подсолнечника |
|
Станица Выселки, Краснодарский край |
2001 |
Ригасол |
|
Станица Привольная, Краснодарский край |
2003 |
Сигнал |
|
г. Светлоград, Ставропольский край |
2005 |
ПР 64 А 44 |
|
Станица Копанская, Краснодарский край |
2006 |
Ригасол ОР |
|
г. Морозовск, Ростовская область |
2006 |
ПР 63 А 90 |
|
Станица Казанская |
2006 |
Румбасол |
|
Тацинский р-н, Ростовская область |
2008 |
неизвестный |
|
Константиновский р-н, Ростовская область |
2008 |
Арена |
|
Турция, смесь рас А-H |
2008 |
Неизвестный |
Электрофорез продуктов амплификации проводили в агарозном геле (1,5 % агароза, 1½ТАЕ-буфер) с использованием камеры для горизонтального электрофореза в течение 1,52,0 ч при I = 50 мА, U = 70-90 В. Реакционную смесь (10 мкл) наносили на гель вместе с красителем бромфеноловым синим. В качестве маркеров длин фрагментов ДНК использовали GeneRuler 1 kb DNA Ladder (UAB «Fermentas», Литва). Последующее окрашивание осуществляли бромистым этидием. Результаты электрофореза документировали при помощи гель-документирующей видеосистемы BIO-PRINT (Vilber Lourmat, Франция). Реакции повторяли 2-3 раза, принимая за достоверные только воспроизводимые фракции. Различия между образцами определялись по наличию или отсутствию полос на геле, соответствующих фрагментам ДНК определенной длины.
Результаты и обсуждение. Для осуществления программы создания устойчивых к заразихе сортов и гибридов подсолнечника необходимы знания о генетической вариабельности паразита. С этой целью нами для молекулярно-генетической характеристики были использованы девять популяций паразита, собранных в разные годы в разных районах юга России и Турции. Изучение этих популяций с помощью набора дифференциаторов позволило нам ранее сделать вывод, что они представлены биотипами с разной вирулентностью [7;12].
Для выявления генетической изменчивости заразихи нами было оценено 48 праймеров для ПЦР. Результаты исследований показали, что из 48 праймеров 16 не гибридизировались с матричной ДНК заразихи. Из оставшихся 32 праймеров 24 не генерировали полиморфизм. Шесть праймеров выявляли либо полиморфизм минорных фракций, либо невоспроизводимый полиморфизм. Лишь два праймера из сорока восьми выявили четкий, воспроизводимый полиморфизм у разных образцов заразихи – RAPD праймеры OPJ13 и OPG06 (табл. 2).
Таблица 2 – RAPD праймеры, выявившие полиморфизм у образцов заразихи
Праймер Нуклеотидная последовательность OPJ13 CCA CAC TAC C
OPG06 GTG CCT AAC C
По праймеру OPJ13 только два образца из тридцати восьми показали отличия от других – по одному образцу из Казанской и Высел-ковской популяций. У этих образцов имелся фрагмент длиной 575 пар нуклеотидов (п.н.), отсутствующий у других (рис. 1).
1 2 3 4 5 6 7 8 9 10 11 М
Рисунок 1 – Электрофоретические спектры продуктов амплификации ДНК с праймером ОРJ13 образцов заразихи. Дорожки: 1 – Копанская; 2, 3 – Казанская; 4, 5 – Тацинско-константиновская, многостеблевые формы заразихи; 6 – Тацинско-конс-тантиновская; 7, 8 – Турецкая; 9 – Выселковская; 10 – Светлоградская; 11 – Выселковская. М – маркер молекулярной массы ДНК 1 kb. Стрелками показан фрагмент ДНК длиной 575 п.н.
По праймеру OPG06 шесть образцов показали отличия от других – Морозовская; Привольненская; два образца Казанской; Турецкая; Выселковская. У этих образцов имелся фрагмент длинной 850 п.н., отсутствующий у других (рис. 2).
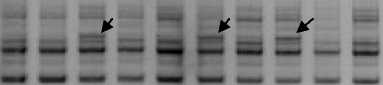
1 2 3 4 5 6 7 8 9 10 М
Рисунок 2 – Электрофоретические спектры продуктов амплификации ДНК с праймером ОРG06 образцов заразихи. Дорожки: 1, 2 – Та-цинско-константиновская, многостеблевые формы заразихи; 3, 4 – Турецкая; 5, 6 – Казанская; 7, 8 – Выселки; 9 – Светлоград; 10 – Турецкая; М – маркер молекулярной массы ДНК 1 kb. Стрелками показана фракция 850 п.н.
Молекулярно-генетическому маркированию подвергали и новые морфотипы заразихи – многостеблевые формы, описанные в работе Антоновой и др. (2010) [9]. Наши результаты показали, что ДНК профили и степень изменчивости данных образцов не отличаются от обычных.
Полученные результаты показали, что образцы заразихи обладают слабым межпопуляционным полиморфизмом.
Только две популяции – Казанская и Выселковская, показали изменчивость по двум праймерам. Популяции Морозов-ская, Привольненская и Турецкая – изменчивость по одному праймеру. Не было выявлено различий между образцами заразихи разных рас. Выявленный слабый полиморфизм показывает, что у популяций O. cumana процесс самоопыления выражен в большей степени, чем перекрестное опыление. Это подтверждается и работами зарубежных авторов [10]. Результаты наших исследований позволяют сделать вывод, что изученные популяции O. cumana произошли из небольшого генетического пула, что объясняет слабый молекулярно-генетический полиморфизм, выявляемый у популяций, разделенных территориально.
В дальнейшем нами планируется продолжение поиска молекулярно-генетических маркеров, способных идентифицировать физиологические расы заразихи.
