Микросателлиты и их использование для оценки генетического разнообразия животных (обзор иностранной литературы)
")
Автор: Марзанов Н.С., Озеров М.Ю., Насибов М.Г., Марзанова Л.К.
Журнал: Сельскохозяйственная биология @agrobiology
Рубрика: Наука за рубежом
Статья в выпуске: 2 т.39, 2004 года.
Бесплатный доступ
Представлен обзор зарубежной литературы, посвященный проблеме применения микросателлитов для исследований эволюции, происхождения и дивергенции сельскохозяйственных животных. На обширном материале рассматривается роль микросателлитов в организме человека и животных разных видов. Обсуждается возможность использования микросателлитов в качестве маркеров для оценки аллелофонда, достоверности происхождения и филогенеза локальных и широкораспространенных пород животных.
Короткий адрес: https://sciup.org/142132958
IDR: 142132958 | УДК: 636:577.2:575
Microsatellites and their application in investigation of genetic diversity of animals (review)
Presented review of forein literature is devoted to application of microsatellites in investigation of evolution and divergence of agricultural animals. On the basis of extensive investigations the role of microsatellites in human and animal organisms was considered. The authors discuss the problems of application of microsatellites as marker system for evaluation of allelic pool, parentage control and phylogenesis of local and widespread breeds of animals.
Текст научной статьи Микросателлиты и их использование для оценки генетического разнообразия животных (обзор иностранной литературы)
Микросателлиты (microsatellites) — это наиболее распространенные тандемные непрерывные повторы участков ДНК, длина которых не превышает шести базовых пар, содержащих от одного до десяти нуклеотидов в одной единице (1-3). Исторически термин «микросателлиты» чаще всего использовали для обозначения динук-леотидных повторов типа СА (GT), а для других последовательностей применяли различные термины — от «простая последовательность» до STR (short tandem repeats), что привело к некоторой путанице. В настоящее время этим термином обозначают в основном короткие тандемные повторы, хотя было бы правильнее все короткие повторы, включая минисателлиты и простые кодирующие участки ДНК (cryptically simple regions), называть «простые последовательности» (4).
Микросателлиты подразделяют на пять основных групп по длине единицы повтора (5): моно-, ди-, три-, тетра-, пентануклеотидные и более (до десяти нуклеотидов включительно). Микросателлиты подразделяют также на основе их структуры на три класса (6): совершенные повторы мотивов (СА) n или (GT) n , которые не прерываются другими нуклеотидами и не расположены рядом с повторами других последовательностей; несовершенные (неполные) повторяющиеся последовательности с двумя или б о льшим числом блоков (CA) n , разделенных не более чем тремя следующими друг за другом неповторяющимися основаниями; сложные повторяющиеся последовательности, у которых блоки (CA) n отделены от других блоков более чем тремя неповторяющимися основаниями, представлены в виде одинаковых нуклеотидов в количестве 5-10 и более. Сложные повторы подразделяются в свою очередь на совершенные и несовершенные в зависимости от типа блоков (CA) n . Совершенные повторы составляют до 80 % микросателлитных последовательностей (табл. 1) (6-8).
Соотношение микросателлитов различных классов в геномах млекопитающих разных видов (%) (цит. по 6 и 7)
|
Класс микросателлитов |
Свинья |
Крупный рогатый скот |
Овца |
Куры |
Собака |
Человек |
|
Совершенные |
71 |
56 |
84 |
64 |
80 |
64 |
|
Несовершенные |
19 |
24 |
10 |
22 |
8 |
25 |
|
Сложные |
10 |
20 |
6 |
14 |
12 |
11 |
Р а с п р е д е л е н и е м и к р о с а т е л л и т о в п о г е н о-м у в наибольшей степени изучено у человека и мыши. Картирование на высоком разрешающем уровне предполагает более или менее равномерное распределение микросателлитов по всему геному обоих видов, однако даже такие карты содержат пробелы и участки с низкой плотностью, которые чаще встречаются вблизи теломер (9, 10).
Микросателлиты редко бывают локализованы в кодирующих участках генома и даже в непосредственной близости от них (11). Это частично относится к микросателлитам, повторяющиеся единицы которых содержат меньше трех оснований, например поли(СА). Тандемные повторы поли(A)/поли(T) наиболее часто встречаются в геноме человека. Однако не представляется возможным использовать микросателлиты для 104
генетического картирования или популяционного анализа в связи с их нестабильностью при проведении полимеразной цепной реакции (ПЦР). Показано, что наиболее распространенными являются динуклеотидные СА/ТG-повторы, которые встречаются в 2 и 3 раза чаще, чем соответственно АТ- и AG/TC-повторы (12). Среди тринуклеотидных повторов наибольшей частотой характеризуются CAG- и AAT-повторы (13). В геномах млекопитающих содержится такой же набор микросателлитов, как и в геноме человека, несмотря на небольшую разницу по частоте аллелей (12, 13). В то же время в геномах наиболее отдаленных видов (например человек и крыса) могут встречаться микросателлиты иной структуры (14).
В геноме дрожжей Saccharomyces cerevisiae выявлено б о льшее количество поли(AT)-повторов, чем в геноме человека (11). У растений GA- и AT-повторы встречаются чаще, чем CA-повторы, которые наиболее распространены в геноме Drosophila melanogaster (14, 15). В геноме бактерий число тандемно повторяющихся последовательностей меньше по сравнению с многоклеточными организмами, но и здесь наиболее распространены поли(A)/поли(T)-повторы (11, 16).
Происхождение этих различий до конца не выяснено. Hancock (1995) подсчитал, что большее количество вариаций по частоте простых последовательностей генома объясняется предполагаемыми механическими сдвигами при репликации ДНК (SSM — slipped strand mispairing), когда строящиеся цепи ДНК начинают соединяться в неправильном порядке. Различия между видами могут быть абсолютно случайными или отражать селективные процессы при отборе последовательностей, адаптированных к определенным функциям (4).
Ф у н к ц и о н а л ь н а я р о л ь м и к р о с а т е л л и т о в. В популяционной генетике микросателлиты используют как генетические маркеры с целью генетического картирования и оценки эволюционных связей. Однако в настоящее время накоплено много данных, свидетельствующих о регуляторной функциональной роли этих элементов в геноме (17). Микросателлиты были обнаружены повсеместно в участках, расположенных левее (upstream) промоторов. Во многих случаях у различных видов млекопитающих микросателлитные последовательности локализованы в одном и том же гене, в участках, близких к промоторам. Например, поли(TG)-повтор в геноме мыши, крысы и человека расположен слева от транскрипционной единицы рРНК, а у двух последних — слева от гена соматостатина (18-20). У кур и мыши цепь повторов (TCCC)2…(TCCC)2 обнаружена в участке, расположенном слева от промотора α 2(I)-гена, контролирующего образование коллагена (21). Последовательность CGG в 5'-области промотора гена FMR1 , связанного с синдромом хрупкости Х-хромосомы, была выявлена у 44 видов млекопитающих (от летучей мыши до дельфина); эта последовательность прошла 150 млн лет эволюции, что доказывает ее функциональную значимость (22). Данные многих исследований свидетельствуют о сохранении микросателлитов в геномах различных млекопитающих на протяжении десятков тысяч лет (13, 14, 23).
Некоторые микросателлиты, расположенные в участках промоторов могут влиять на функцию энхансера при экспрессии генов. Так, фрагмент ДНК, локализованный слева от гена Еβ иммуноглобулина и содержащий микросателлитные участки, служит в качестве позитивного энхансера (24). Однако имеются микросателлиты, которые ингибируют активность промотора, например расположенный слева от α 2(I)-гена коллагена кур длинный фрагмент ДНК, содержащий множество микроса-теллитных участков — (GGAA) 25 , (GGGA) 6 , (GGGAGGAA) 4 , (GGGGGGAA) 2 (25). Возможно, что в некоторых случаях неактивные участки микросателлитов все же несут какую-то функцию in vivo (4).
Показано, что короткие нуклеотидные последовательности, находящиеся вблизи генов, могут служить сайтами связывания различных белков, так же, как и промоторы, содержащие микросателлиты (26). Способность микросателлитов взаимодействовать с белками зависит от количества тандемных повторов на специфическом фрагменте последовательности, который может принимать участие в синтезе белков и особенно гомополимерных участков аминокислот (4).
Вариации повторяющихся повторов в микросателлитах оказывают влияние на фенотип, что наиболее наглядно можно проследить при генетических заболеваниях человека, вызванных увеличением тринуклеотидных повторов (27). Впервые взаимосвязь между заболеванием и увеличением числа повторов была отмечена в 1991 году при исследовании синдрома хрупкости Х-хромосомы (наиболее распространенная форма задержки умственного развития). К 1995 году было идентифицировано 11 локусов, в которых наблюдалось увеличение длины поли(CGG)-повторов (10-1000-кратное) и поли(CAG)-повторов (в 2-3 раза) по сравнению с длиной фрагментов у здоровых людей. Эти локусы обусловливают соответственно задержку умственного развития (пять сайтов) и нейродегенеративные заболевания (шесть сайтов). В настоящее время список заболеваний расширился и включает синполидакти-лию, атаксию Фридриха, аутосомальную доминантную спастическую параплегию и аутосомальную доминантную окулофарингеальную мышечную дистрофию, которые связаны с увеличением частоты следующих повторов: соответственно поли(CGG), по-ли(CAA), поли(CAG) и поли(GCG) (28-30).
М у т а ц и о н н ы е и з м е н е н и я м и к р о с а т е л л и - т о в. Частота мутаций микросателлитов сравнима с частотой точечных мутаций — 10–9-10–10. Предположительная частота мутаций на один локус in vivo составляет у Escherichia coli и дрожжей соответственно около 10–2 и 10–5 (31, 32). В работах по определению отцовства у людей установлена средняя частота мутаций на один локус (на одно поколение), которая равна 10–3 (33). Частота мутаций микросателлитов у мыши, овцы, свиньи и дрозофилы составляет соответственно 10–3-10–4, 1,3 ⋅ 10–4, 7 ⋅ 10–5 и 6 ⋅ 10–6 (34-38). Предполагают, что такая высокая частота мутаций микросателлитов возникает либо при ошибках репликации отдельной ДНК (31), либо в процессе рекомбинации между молекулами ДНК (39).
Ошибки во время репликации (SSM) появляются в то время, когда синтезирующаяся молекулярная цепь ДНК отделяется от матрицы. При репликации неповторяющихся последовательностей новая цепь присоединяется к матрице строго по последовательности нуклеотидов, а в случае повторяющихся фрагментов может произойти сбой при считывании, поэтому новая молекула ДНК будет короче или длиннее, чем матричная цепь; это зависит от того, где происходит сдвиг: если на матричной цепи, то продукт будет короче, если же на синтезирующейся цепи — длиннее.
Рекомбинация потенциально может повлиять на длину микросателлитов либо в результате неравного кроссинговера, либо конверсии генов. Неравный кроссин-говер обычно происходит между хромосомами, имеющими длинные тандемные повторы. При этом может возникнуть делеция в одной молекуле ДНК и вставка в другой, а также увеличиться или уменьшиться число вариантов в тандемно повторяющихся последовательностях (40). При конверсии генов происходит перенос последовательностей из одного аллеля в другой, то есть в локусах меняется порядок повторов в результате встраивания новых последовательностей. Большинство исследований все же свидетельствует о ведущей роли SSM в образовании мутаций в микросателлитах (32, 35).
Применение микросателлитов для иссле-дования э в о л ю ц и и с е л ь с к о х о з я й с т в е н н ы х ж и- в о т н ы х. Микросателлиты используют как для анализа эволюционной связи между различными породами сельскохозяйственных животных, так и для определения времени появления той или иной породы. Например, при исследовании шести пород овец (ромни-марш, авасси, бордер, суффолк, австралийские и новозеландские мериносы) при помощи восьми микросателлитов (MAF33, MAF48, MAF65, MAF209, OarFCB11, OarFCB128, OarFCB266 и OarFCB304) показано, что между австралийскими и новозеландскими мериносами генетическое расстояние было наименьшим (0,08), а между породами авасси и бордер — наибольшим (0,50) (41). Между остальными четырьмя породами и породой авасси этот показатель варьировал от 0,36 до 0,50, а между породами ромни-марш, суффолк и бордер — от 0,26 до 0,30.
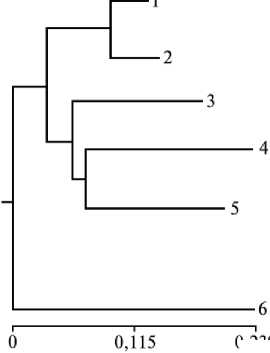
Дендрограмма взаимосвязи между шестью породами овец на основе анализа восьми микросателлитов: 1, 2, 3, 4, 5 и 6 — соответственно новозеландский меринос, австралийский меринос, ромни-марш, бордер, суффолк и авасси (цит. по 41).
0,230
Генетическое расстояние
На основе этих данных авторы построили дендрограмму, состоящую из трех ветвей, в одну из которых входили мериносы, в другую — породы ромни-марш, бордер и суффолк, а в третью — порода авасси, расположенная на большом расстоянии от остальных (рис.). Результаты этих исследований свидетельствуют о том, что расхождение между мериносами и британскими породами овец произошло не более чем 2000 лет назад (точнее 1094 года назад). На основании экспериментов с использованием 30 локусов белков крови эта дата определена как 69700 лет, а время расхождения между австралийскими и новозеландскими мериносами составляет 227 лет (42).
Микросателлиты (в отличие от белковых маркеров) позволяют более точно рассчитать время дивергенции популяций или пород, происходящих от общего предка. Так, в работе Diez-Tascon с соавт. были выявлены генетические связи между шестью популяциями мериносовых овец из Испании, Португалии и Новой Зеландии (испанский и новозеландский меринос, французский и немецкий мясной меринос, португальский черный и белый меринос) с помощью 20 микросателлитов: BM1824, BM4621, BM6444, BM6506, BM757,
ETH225, MAF209, MAF23, McM214, McM218, McM357, OarCP20, OarCP34,
OarCP49, OarFCB11, OarFCB128, OarFCB20, OarFCB304, OarFCB48, OarHH64 (43). При этом оказалось, что между испанскими и португальскими черными мериносами генетическое расстояние было наименьшим (0,086), между другими популяциями Пиренейского полуострова оно варьировало от 0,135 до 0,086. Генетическое расстояние между французскими и немецкими мериносами составляло 0,139, а между этими породами и новозеландскими мериносами — соответственно 0,356 и 0,328. Популяции, имеющие общую продуктивную направленность (мясная или шерстная), были объединены в общие кластеры.
Сравнительный анализ общности животных на основе оценки вероятности происхождения по микросателлитам позволяет определить родословную и принадлежность к той или иной породе. Например, анализ происхождения австрийских домашних овец ( Ovis aries ) проводили по шести микросателлитным локусам — McM42, McM527, OarFCB20, INRA49, TGLA53 и MAF65 (44). При этом рассматривалась возможность использования для микросателлитного анализа образцов различных тканей и жидкостей (кровь, ткани кожи, эпителий внутренней стороны щек, волосы и др.). Наиболее пригодным материалом оказались образцы волос, которые сохраняются при температуре окружающей среды без всякой предварительной консервации, что уменьшает расходы, а также они могут быть легко собраны.
Анализ происхождения пород крупного рогатого скота на основе микросател-литных маркеров представляет особенную ценность при формировании широкоразво-димых стад, в которых используют животных, полученных от нескольких производителей. Так, при определении родственных связей между несколькими группами крупного рогатого скота (по 50 гол. в каждой), включающими животных разных пород, оценивали эффективность 21 микросателлитного маркера (45). Для выявления отцовства анализировали 5960 телят (по 50-500 гол. в группе), полученных от производителей из 39 групп, включающих от 4 до 34 быков. В большинстве тестируемых групп количество телят, полученных от неизвестных быков, существенно различалось. При этом оказалось, что если доля телят, полученных от неизвестных производителей, составляет более 50,8 %, то точность определения отцовства возможна в 86 % случаев; в тех вариантах, где имелись сведения о быках-производителях, точность анализа составляла 100 %.
Polli с соавт. оценивали генетический полиморфизм семи аутохтонных (местных) итальянских пород крупного рогатого скота с целью определения их генетической структуры и филогенетического происхождения (46). Изучение генетического разнообразия аутохтонных пород имеет большое значение для программ сохранения биоразнообразия видов крупного рогатого скота. Породы Aosta Red Pied, Aosta Black Pied и Aosta Chestnut разводят в предгорьях Альп, а породы Oropa, серая альпийская, Rendena и Burlina — в других районах Италии. Для анализа использовали 15 высокополиморфных микросателлитов из набора, одобренного Международным обществом по генетике животных (МОГЖ), для изучения биоразнообразия крупного рогатого скота. ДНК выделяли из крови или спермы и амплифицировали посредством ПЦР. Аллели визуализировали автоматическим секвенатором в 4,2 % полиакриламидном геле. Оценивали гетерозиготность каждого локуса, частоту аллелей и PIC (polymorphism information content), рассчитывали генетические расстояния. При этом наименьшее генетическое расстояние выявлено между породами Aosta Red Pied и Aosta Black Pied, а наибольшее — Burlina и Aosta Black Pied.
Высокий уровень полиморфизма и случайное расположение в геноме позволяют использовать микросателлиты для идентификации отцовства, не генотипируя самок. Genzini с соавт. проанализировали 444 семьи итальянских буйволов (бык-производитель—дочь) с использованием шести микросателлитных локусов (DRB3, CSSM47, D5S2, MAF65, CSSM19, CYP21) и гена главного комплекса гистосовместимости класса II овец (47). По частоте аллелей общая вероятность исключения составила 0,98, то есть ошибка определения происхождения равна 2 %.
При исследовании происхождения финской айрширской и голштинской пород крупного рогатого скота были использованы девять микросателлитных локусов в различных комбинациях в трех реакциях мультиплекс-ПЦР, согласно стандартному протоколу: мультиплекс 1 — BM2133, BM1824, SPS115; мультиплекс 2 — ETH3, ETH225, ETH10; мультиплекс 3 — TGLA227, TGLA126, TGLA122 (48). В третью реакцию (мультиплекс 3) были включены дополнительные локусы — TGLA53 и INRA023 для анализа частоты аллелей и вероятности исключения. Высокий уровень полиморфности в популяции скота айрширской породы ( n = 50) выявлен по локусам BM2113, TGLA227, ETH10 и ETH225, голштинской породы ( n = 50) — по локусам TGLA227, TGLA122 и BM2113; вероятность исключения составляла соответственно 56 и 62 %. Общая вероятность исключения при использовании девяти локусов достигала 99,84 и 99,91 % соответственно у животных айрширской и голштинской пород; при использовании дополнительных локусов TGLA53 и INRA023 полученные ранее показатели возросли соответственно до 99,94 % и 99,98 %.
Для выявления полиморфизма у коз пяти китайских аборигенных популяций (льаонин кашмир, n = 33; внутренний монгольский кашмир, n = 35; тибетский кашмир, n = 37; вуан кашмир, n = 49 и мясные козы мату, n = 39) использовали микросателлиты крупного рогатого скота (BM1842, BM4621, BM6444, BM6506, BM757) и овец (OarCP20, OarCP34, OarCP49, OarFCB11, OarFCB11, OarFCB128, OarFCB20, OarFCB266, OarFCB304, OarFCB48, MAF33, McM218) (49). Более четырех аллелей были обнаружены в шести локусах (BM4621, OarCP34, OarFCB11, OarFCB20, OarFCB304, OarFCB48), другие девять микросателлитов оказались мономорфными. Определяли внутрипородное разнообразие, гетерозиготность и PIC. При этом рассчитывали общее генетическое разнообразие, среднюю гетерозиготность в пределах каждой популяции и коэффициенты дифференциации гена между породами; средние значения этих параметров составляли соответственно 0,831; 0,800; 0,038. По данным этих исследований, межпородное различие китайских аборигенных коз находится на низком уровне, тогда как внутрипородное разнообразие оказалось достаточно высоким.
Таким образом, представленный обзор свидетельствует о том, что микросателлиты обладают рядом свойств (высокий уровень полиморфизма и изменчивости, кодоминантное наследование, равномерное распределение по геному и др.), которые позволяют использовать их в качестве маркеров при оценке происхождения и эволю-108
ции различных видов и пород животных (50). С помощью микросателлитов можно идентифицировать все кодоминантно наследуемые аллели закрытого локуса, что не всегда возможно при анализе генетических маркеров I порядка, оценивать генетическое разнообразие популяций; они также подходят для теста на достоверность отцовства, оценки дивергенции пород, теста на диагностику явления «бутылочного горлышка» (bottleneck) в популяции. Недостатком этого метода является дороговизна, необходимость солидной материальной базы и специалистов, которых, к сожалению, пока в нашей стране мало.
Л И Т Е Р А Т У Р А
-
1. T a u t z D. Hypervariability of simple sequences as a general source for polymorphic DNA markers. Nucleic Acids Research, 1989, 17: 6463-6471.
-
2. T a u t z D., R e n z M. Simple sequences are ubiquitous repetitive components of eukariotic genomes. Nucleic Acids Research, 1984, 12: 4127-4138.
-
3. W r i g h t J.M. Mutation at VNTRs: are minisatellites the evolutionary progeny of microsatellites? Genome, 1994, 37: 345-347.
-
4. G o l d s t e i n D.B., S c h l o t t e r e r C. Microsatellites. Evolution and Applications. Oxford, 1999.
-
5. V o g t P. Potential genetic functions of tandem repeated DNA sequence blocks in human genome are based on
highly conserved chromatine folding code. Human Genetics, 1990, 84: 301-336.
-
6. W e b e r J.L. Informativeness of human (dC-dA) n ⋅ (dG-dT) n polymorphisms. Genomics, 1990, 7: 524-530.
-
7. W i n t e r o A.K., F r e d h o l m M., T h o m s e n P.D. Variable (dG-dT) n (dC-dA) n sequences in the
porcine genome. Genomics, 1992, 12: 281-288.
-
8. V a i m a n D., I m a m - C h a l i M., M o a z a m i - G o u d a r z i K. e.a. Conservation of a synthetic group of microsatellite loci between cattle and sheep. Mammalian Genome, 1996: 676-683.
-
9. D i b C., F a u r e S., F i z a m e s C. e.a. A comprehensive genetic map of the human genome based on 5264 microsatellites. Nature, 1996, 380: 149-152.
-
10. M o r a n C. Microsatellite repeats in pig ( Sus domestica ) and chicken ( Gallus domesticus ) genomes. Zoo Biology, 1993, 10: 341-352.
-
11. H a n c o c k J.M. The contribution of slippage-like processes to genome evolution. J. Molecular
Evolution, 1995, 41: 1038-1047.
-
12. B e c k m a n J.S., W e b e r J.L. Survey of human and rat microsatellites. Genomics, 1992, 12: 627-631.
-
13. S t a l l i n g s R.L. Distribution of trinucleotide microsatellites in different categories of mammalian genomic se
quence: implications of human genetic diseases. Genomics, 1994, 21: 116-121.
-
14. S t a l l i n g s R.L. CpG supression in vertebrale genomes does not account for the rarity of (CpG) n microsatellite repeats. Genomics, 1992, 17: 890-891.
-
15. S c h u g M.D., W e t t e r s t r a n d K.A., G a u d e t t e M.S. e.a. The distribution and frequency of microsatellite loci in Drosophila melanogaster . Molecular Ecology, 1998, 7: 57-69.
-
16. H a n c o c k J.M. Simple sequences in a «minimal» genome. Nature Genetics, 1996, 13: 14-15.
-
17. K a s h i Y., K i n g D., S o l l e r M. Simple sequence repeats as a source of quantitative genetic variation.
-
18. B r a a t e n D.C., T h o m a s J.R., L i t t l e R.D. e.a. Locations and contexts or sequences that hybridize to poly(dG-dT) n ⋅ (dC-dA) n in mammalian ribosomal DNAs and two X-linked genes. Nucleic Acid Research, 1988, 18: 865-881.
-
19. S h e n L.-P., R u t t e r W.J. Sequence of the human somatostatine gene. Science, 1984, 224: 168-170.
-
20. H a y e s T.E., D i x o n J.E. Z-DNA in the rat somatostatin gene. J. Biological Chemistry, 1985, 260:
-
21. M c K e o n C., S c h m i d t A., d e C r o m b r u g g h e B. A sequence conserved in both the chicken
and mouse α 2(I) collage promoter contains sites sensitive to S1 nuclease. J. Biological Chemistry, 1984,
259: 6636-6640.
-
22. E i c h l e r E.E., K u n s t C.B., L u g e n b e e l K.A. e.a. Evolution of the cryptic FMR1 CGG repeat.
-
23. M o o r e S.S., S a g e a n t L.L., K i n g T.J. e.a. The conservation of dinucleotide microsatellites among
mammalian genomes allows the use of heterologous PCR primer pairs in closely related species. Genomics, 1991, 10: 654-660.
-
24. G i l l i e s S.D., F o l s o m B., T o n e g a w a S. Cell-type specific enhancer element associated with a mouse MHC gene. Nature, 1984, 310: 594-597.
-
25. K o l l e r E., H a y m a n A.R., T r u e b B. The promoter of the chicken a2(VI) collagen gene has features characteristic of house-keeping genes and of proto-oncogenes. Nucleic Acids Research, 1991, 19: 485491.
-
26. L u e N.L., B u c h m a n A.R., K o r n b e r g R.D. Activation of yeast RNA polymerase II transcription by a thymidine-rich upstream element in vitro. Proceedings of the National Academy of Sciences of the USA, 1989, 86: 486-490.
-
27. S u t h e r l a n d G.R., R i c h a r d s R.I. Simple tandem DNA repeats and human genetic disease. Proceedings of the National Academy of Sciences of the USA, 1995, 92: 3636-3641.
-
28. C a m p u z a n o V., M o n t e r m i n i L., M o l t o M.E. e.a. Friedreich’s Ataxia: autosomal recessive disease caused by an intronic GAA triplet repeat expansion. Science, 1996, 271: 1423-1427.
-
29. N i e l s e n R. A likelihood approach to population samples of microsatellite alleles. Genetics, 1997, 146: 711-716.
-
30. B r a i s B., B o u c h a r d J.-P., X i e Y.-G. e.a. Short GCG expansions in the PABP2 gene cause oculo-pharingeal muscular distrophy. Nature Genetics, 1998, 18: 164-167.
-
31. L e v i n s o n G., G u t m a n G.A. High frequencies of short frameshifts in poly-CA/TG tandem repeats borne by bacteriophage M13 in Escherichia coli K-12. Nucleic Acid Research, 1987, 15: 5323-5328.
-
32. H e n d e r s o n S.T., P e t e s T.D. Instability of simple sequence DNA in Saccharomyces cerevisiae . Molecular and Cell Biology, 1992, 12: 2749-2757.
-
33. W e b e r J.L., W o n g C. Mutation of human short tandem repeats. Human Molecular Genetics, 1993, 2: 1123-1128.
-
34. D a l l a s J.F. Estimation of the microsatellite mutation rates in recombinant inbred strains of mouse. Mammalian Genome, 1992, 3: 452-456.
-
35. C r a w f o r d A.M., C u t h b e r t s o n R.P. Mutations in sheep microsatellites. Genome Research, 1996, 6: 876-879.
-
36. S c h u g M.D., M a c k a y T.F.C., A q u a d r o C.F. Low mutation rates of microsatellite loci in Drosophila melanogaster . Nature Genetics, 1997, 15: 99-102.
-
37. K w i a t k o w s k i D.J., H e n s k e E.P., W e i m e r K. e.a. Construction of GT polymorphism map of human 9q. Genomics, 1992, 12: 229-240.
-
38. E l l e g r e n H. Mutation rates at porcine microsatellite loci. Mammalian Genome, 1995, 6: 376-377.
-
39. J e f f r e y s A.J., T a m a k i K., M a c L e o d K. e.a. Complex gene conversion events in germline muta
tion at human minisatellites. Nature Genetics, 1994, 6: 136-145.
-
40. D o v e r G.A. Molecular drive: a cohesive mode of species evolution. Nature, 1982, 299: 111-117.
-
41. B u c h a n a n F.C., A d a m s L.J., L i t t l e j o h n R.P. e.a. Determination of evolutionary relationships
among sheep breeds using microsatellites. Genomics, 1994, 22: 347-403.
-
42. M a n w e l l C., B a c k e r C.M.A. Genetic distance between the Australian Merino and the Poll Dorset sheep. Genetics Research Cambridge, 1977, 29: 239-253.
-
43. D i e z - T a s c o n C., L i t t l e j o h n R.P., A m e i d a P.A.R. e.a. Genetic variation within the Merino sheep breed: analysis of closely related populations using microsatellites. Animal Genetics, 2000, 31: 243251.
-
44. A c h m a n n R., B r e m G. Parentage control in Austrian domestic mountain sheep ( Ovis aries ) using DNA microsatellite analysis. Animal Genetics, 1998, 29: 12-13.
-
45. V a n k a n D.M., H e f f o r d C.G., F a d d y M.J. Efficacy and reliability of paternity testing in cattle using DNA microsatellites. Animal Genetics, 1998, 29: 9.
-
46. P o l l i M., D e l B o L., C e r i o t t i G. e.a. Genetic distances of seven cattle breeds of the Italian Alpine area by microsatellites. Animal Genetics, 1998, 29: 9-10.
-
47. G e n z i n i E., B l a s i M., C a p u a n o M. e.a. Amplification of seven microsatellites for parentage identification in Italian buffalo. Animal Genetics, 1998, 29: 10.
-
48. B r e d b a c k a P., K o s k i n e n M.T. Polymorphism of microsatellite loci used in parentage testing in Finnish Ayrshire and Holstein populations. Animal Genetics, 1998, 29: 10-11.
-
49. Z h a o S., Y a n g L., L i K. e.a. Analysis of genetic variation among five Chinese indigenous goat breeds by using bovine and ovine microsatellites. In: Animal Genomics: Synthesis of Past, Present, and Future Directions. 27th Intern. Conf. on Animal Genetics. USA, 2000.
-
50. M a l e v i C i U t ё J., B a 11 r ё n a i t ё L., M i c e i k i e n ё I. Domestic cattle breed diversity in Lithuania. Veterinary Medicine and Zootechnics, 2002, 20: 87-91.
Trends in Genetics, 1997, 13: 74-78.
8145-8156.
Nature Genetics, 1995, 11: 301-307.
Всероссийский государственный НИИ животноводства,
