Многообразие симбиотических отношений и возможный подход к их классификации

Автор: Озерский Павел Викторович
Журнал: Общество. Среда. Развитие (Terra Humana) @terra-humana
Рубрика: Природная среда
Статья в выпуске: 4 (29), 2013 года.
Бесплатный доступ
С учетом консортивной природы симбиотических отношений разработана классификация симбиозов и близких явлений. Симбиозы подразделены на физиологические и экологические и противопоставлены динамическим консортивным связям. Обсуждается применимость к симбиотическим системам демоцентрической и ценоцентрической систем отсчета.
Консорция, метаболические связи, симбиоз, среда жизни
Короткий адрес: https://sciup.org/14031659
IDR: 14031659 | УДК: 574.2+574.3+574.4+574.5+591.557
Текст научной статьи Многообразие симбиотических отношений и возможный подход к их классификации
Введение. Описывая структуру теоретической экологии, мы [11] предложили распределить ее понятия по двум системам отсчета, демоцентрической (выстраиваемой вокруг популяции) и ценоцентрической (выстраиваемой вокруг экосистемы или сообщества). Некоторые затруднения вызывает рассмотрение с этой точки зрения симбиотических систем, так как они способны образовывать группировки, подобные популяциям (по А.Б. Савинову [16] – демоценозы), но при этом сами являются двух- или многовидовыми сообществами.
Для разрешения этих затруднений нужно определиться с трактовкой понятия симбиоза, различающейся у разных авторов. С учетом истории и синонимии термина мы [10] обосновали предпочтительность «широкой» трактовки симбиоза, не предполагающей обязательной взаи-мовыгодности отношений (одной из форм симбиоза оказывается паразитизм). Однако «широкая» трактовка позволяет считать симбиозами очень разные явления. Поэтому в настоящей работе мы попытались дать, в самом общем виде, классификацию симбиозов и смежных явлений.
Принципы предлагаемой классификации. Важным принципом нашей классификации является признание симбиотических систем вариантом индивидуальных консорций. Оно следует из работы В.Н. Беклемишева [1], где впервые был употреблен термин «консорций» в значении, близком к таковому современного термина «индивидуальная консорция» [5] (начиная со статьи Е. М. Лавренко [7], сложилась традиция использовать термин «консорция» в женском роде.). Как примеры «консорциев» в ней были приведены «сосна с ее микоризными грибками, с эпифитными мхами и лишайниками на стволе и ветвях, с паразитическими грибками в различных тканях, со всем множеством населяющих ее членистоногих» и «полевка с ее эктопаразитами, с гельминтами, простейшими и бактериями, населяющими ее внутренние органы» [1, с. 7]. Легко заметить, что в составе «консорциев» Беклемишевым упомянуты исключительно участники симбиозов: паразитизма, комменсализма и мутуализма.
Второй особенностью нашей классификации является частое обращение к понятию среды обитания. Основой для различения симбиоза и не-симбиоза в ней послужил восходящий к работам В.А. Догеля [3; 4] «критерий двух сред». Мы предлагаем следующую формулировку: при симбиозе по крайней мере у одного из участников отношений имеются две среды обитания: среда первого порядка, создаваемая другим участником этих отношений, и среда второго порядка, в которой существует вся симбиотическая система . Первоначально такой подход использовался лишь в отношении паразитизма, позднее С.С. Шульман и А.А. Добровольский [18] показали его применимость также и к других формам симбиоза.
Считая симбиотические системы консорциями, мы используем в их отношении принятую для консорций терминологию. «Макросимбионта» («хозяина») мы называем эдификатором [1; 14], а «микросимбионтов» (паразитов, комменсалов и т. п.) – консортами [8]. Cуть взаимоотношений между участниками симбиозов та же, что во всех консорциях: консорты так или иначе используют среды, созданные или трансформированные эдификаторами. Это не означает, что консорты сами не трансформируют и не порождают никаких сред, однако их роль в этих процессах обычно существенно меньше, чем у эдификаторов. Один организм одновременно может быть консортом для второго и эдификатором для третьего, как это имеет место, например, при гиперпаразитизме, когда парази-
Cреда обитания
тический организм оказывается хозяином для паразита следующего порядка.
Для различения форм консортивных отношений мы используем нашу классификацию сред жизни [12], в которой введены такие понятия, как биогенные среды (организменные – внутренние среды живых организмов; периорганизменные – формирующиеся вблизи особи в ходе ее средообразовательной деятельности; пос- торганизменные – когда-то возникшие в результате средообразовательной деятельности особи и какое-то время сохраняющиеся после ее смерти или выселения) и биогенные трансформанты сред (среды, частично изменившие свои свойства под
Terra Humana
влиянием живых организмов, но сохранившие значительное сходство со своим исходным состоянием).
физиосимбиоз и экосимбиоз. Термин «симбиоз» был введен в науку в XIX в. А. де Бари для обозначения «явлений совместной жизни разноименных организмов» [19, с. 5]. Следующей вехой в развитии концепции симбиоза можно считать выдвижение Догелем «критерия двух сред» и расширение области его применения Шульманом и Добровольским. По мере развития методов биологических исследований возникали предпосылки для дальнейшего уточнения этой концепции. Накопилось много данных о способности симбионтов-консортов (прежде всего, паразитов) биохимически регулировать жизненные процессы эдификаторов (хозяев), влияя на их морфогенез, физиологические функции и поведение [9; 20; 22; 25]. Известны также и примеры биохимической зависимости симбионтов-консортов от эдификаторов: например, такова стимуляция роста микоризных грибов веществами, вырабатываемыми растениями [17]. Описано много примеров обмена между участниками симбиозов сигнальными молекулами, координирующими совместный метаболизм [24]. А.Э. Дуглас [23] отметила появление у организмов при переходе к симбиозу новых «метаболических способностей», например, вовлечение животных в процессы фото- и хемосинтеза.
Наконец, в 2006 г. Н.А. Проворов и Е.А. Долгих [13] предложили три критерия сим-биотичности отношений. Прежде всего, по их мнению, всем симбиотическим системам свойственен особый характер метаболических связей, «в основе которых при симбиозе лежит не механический перенос питательных веществ, а формирование общих биохимических путей» [13, с. 418]. При этом, однако, они отметили формаль- ное соответствие этому критерию явления синтрофии – последовательного осуществления разными видами микроорганизмов разных этапов сложных биохимических процессов – например, деструкции мертвой органики. Поскольку Проворов и Долгих не причисляли синтрофию к симбиозам, тем самым они признали недостаточность «биохимического» критерия. Поэтому они предложили два «генетико-эволюционных» критерия симбиоза: «во-первых, при симбиозах метаболические отношения основаны на взаимной регуляции генов путем сигнального взаимодействия, а нередко и на закономерных изменениях организации генов, что не характерно для биоценозов <...>. Во-вторых, эволюция симбиоза – это повышение интегрированности партнеров и усиление их взаимной зависимости, которые в итоге могут приводить к преобразованию надорганизменной системы в единый организм» [13, с. 418].
Однако оба «генетико-эволюционных» критерия также недостаточны для выявления симбиоза. «Взаимная регуляция генов путем сигнального взаимодействия» может носить как прямой, так и косвенный характер. Прямой мы считаем регуляцию, при которой вещества, производимые одним из симбионтов, непосредственно взаимодействуют с ДНК (или, по меньшей мере, с белками – регуляторами экспрессии генов) второго. Такая регуляция, действительно, известна [13], но многие примеры, использованные Проворовым и Долгих для иллюстрации «первого генетико-эволюционного» критерия, не укладываются в ее рамки, так как в них участвуют вещества-посредники. Однако если считать симбиотическими системы, основывающиеся на непрямой регуляции, то их круг неоправданно расширится. Так, симбиозами окажутся все отношения, в которых имеют место перестройки метаболизма организма (несомненно, имеющие в своей основе регуляцию экспрессии генов) в ответ на химические контакты с представителями других видов (например, реакции на кай-ромоны и алломоны).
Не является надежным признаком сим-биотичности также и «интегрированность партнеров», «их взаимная зависимость», так как они свойственны также и несимбиотическим межвидовым взаимодействиям, например, хищничеству. У жертвы, например, это может быть выработка ядов, защищающих от хищника, способность своевременно распознавать вещества-кай-ромоны, выделяемые хищниками, и т.п. У хищника это могут быть специфический ферментный состав пищеварительных соков, учитывающий химический состав жертв, способность нейтрализовать или использовать в своих целях яды, содержащиеся в жертвах, и т.п.
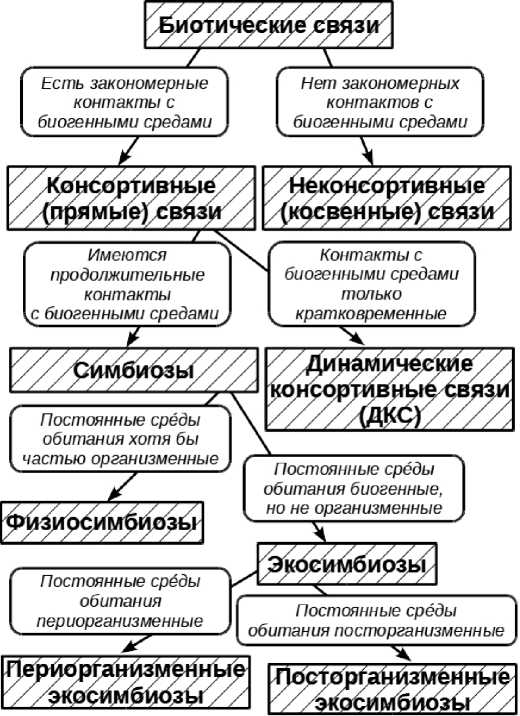
Таким образом, все три критерия Про-ворова и Долгих недостаточны для разграничения симбиозов и не-симбиозов и не могут заменить «критерий двух сред». Ими можно лишь дополнить этот критерий, но с важным уточнением. Концепция Проворова и Долгих предполагает наличие тесных прямых взаимодействий между участниками симбиоза при отсутствии каких-либо неживых посредников, то есть хотя бы у консорта среда обитания (первого порядка) должна быть организменной. Симбиоз такого типа выходит за рамки предмета исключительно экологии и может также изучаться с точки зрения структуры морфологией, а с точки зрения функций – физиологией. Поэтому мы предлагаем для обозначения симбиоза, соответствующего, помимо критерия «двух сред», также и критериям Проворова и Долгих, название « физиологический симбиоз» (или «физиосимбиоз»).
Однако всё разнообразие симбиотических отношений к физиосимбиозу свести невозможно. Среда обитания первого порядка в симбиотических системах, соответствующих определению де Бари [19] и современной трактовке «симбиоза в широком смысле» [15], – это не обязательно организменная среда. Это может быть любая среда, свойства которой определяются жизнедеятельностью одного или нескольких участников экологических отношений (то есть биогенная среда [12]). Мы предлагаем считать биогенность среды первого порядка, сформированной эдификатором и населенной консортом (консортами), еще одним формальным критерием симбиоза.
Биосистемы, образованные представителями двух и более видов и характеризующиеся тем, что в них одни из участников (эдификаторы) формируют для других (консортов) не организменную, но биогенную среду (или биогенную трансформанту абиогенной), могут считаться симбиотическими, так как удовлетворяют «критерию двух сред». Однако прямых взаимодействий между организмами (в том числе и метаболических) в таких системах нет, они опосредованы неживой средой. Мы обозначаем варианты симбиоза, в которых среда первого порядка, используемая участниками-консортами, не является организменной, термином « экологический симбиоз» («экосимбиоз»).
Все экосимбиозы можно подразделить на две группы. Экосимбиозы, в которых среда-посредник является периорганиз-менной, мы обозначаем как периорга-низменные . Их консорты обитают в специфических средах, сформированных и постоянно или регулярно поддерживаемых эдификаторами. При этом они практически не контактируют с организменной средой своих эдификаторов-хозяев. Примерами таких экосимбиозов являются, в частности, явления обрастания раковин моллюсков водорослями, мшанками и другими сидячими организмами.
Экосимбиозы, в которых консорты населяют посторганизменные среды, мы называем посторганизменными . Их особенностью является разделенность во времени средообразовательной деятельности эдификатора и заселения сформированной им среды консортами. Принадлежность отношений такого типа к симбиозам не всегда очевидна. Если они носят эксплуатационный характер, то их рассмотрение как особых форм паразитизма вполне традиционно (достаточно вспомнить насекомых – инквилинов пчел, сначала уничтожающих в ячейке яйцо хозяина, а потом длительное время питающихся имеющимися в ней запасами).
Сложнее обстоит дело с посторганизменными симбиозами типа комменсализма (это, например, обитание сапротрофных организмов в трупах животных, в мертвой древесине и т.п.). Помимо разделенности во времени деятельности эдификатора и консортов, здесь нередко имеет место относительная кратковременность отношений из-за неспособности посторганизменных сред к самоподдержанию. Вместе с тем, посторганизменные экосимбиозы, как и все симбиозы, всецело зависят от средообразовательной деятельности их эдификаторов, а их консорты используют специфические биогенные среды первого порядка.
Кроме того, с прочими симбиозами этот вариант отношений связывают, во-первых, многочисленные случаи специализации консортов к использованию определенных видов эдификаторов и, во-вторых, существование переходных форм между посторганизменными экосимбиозами и другими формами симбиозов. Например, использование усыхающих деревьев насекомыми-ксилофагами не только часто сопровождается узкой пищевой специализацией последних, но и дает целый спектр перехода от физиосимбиоза к посторганизменному экосимбиозу [2].
Cреда обитания
Динамические консортив-ные связи. В симбиотических отношениях (как в физио-, так и в экосимбиозах) всегда устанавливаются более или менее тесные и продолжительные отношения между индивидуумами – эдификаторами и консортами. Даже если существование организма-эдифика-тора и организмов-консортов разнесено во времени (в постбиотических экосимбиозах), то консорты все равно приурочены к среде, трансформированной определенной особью эдификатора. Что же касается отношений, в которых преобразованная эдификаторами среда посещается консортами лишь на краткие промежутки времени (как это свойственно, например, гетеротопным (постоянно перемещающимся между сообществами) животным [21]), то их придется признать несимбиотическими в силу несоответствия «критерию двух сред». Подобные биотические связи мы предлагаем называть динамическими консортивны-ми связями (ДКС).
Как и симбиозы, ДКС могут быть подразделены на группы по приуроченности консортов к определенным биогенным средам. Например, к организменным средам приурочены хищники и пастбищники, к пе-
Постоянные среды обитания хотя бы частью организменные
Постоянные среды обитания биогенные, но не организменные
Постоянные среды
J обитания посторганизменные
Постоянные среды обитания ь периорганизменные
Контакты с биогенными средами только кратковременные
Имеются ' продолжительные контакты с биогенными средами
Нет закономерных контактов с биогенными средами ^
Есть закономерные контакты с t биогенными средами
Рис. 1. Схема классификации биотических связей, основанной на специфике сред обитания и взаимодействий организмов со средами. В заштрихованных прямоугольниках приведены названия, в незаштрихованных – основные признаки выделенных типов связей.
Terra Humana
риорганизменным – например, птицы, охотящиеся на насекомых-кровососов вблизи крупных млекопитающих, к посторганизменным – временные посетители падали (например, грифы). Это подразделение имеет, однако, более условный характер, чем в случае симбиозов, что связано с меньшей, в целом, специализацией и с бульшим разнообразием ДКС по сравнению с симбиозами.
Общая схема классификации, отражающая взаимоотношения между разными формами симбиозов и смежных с ними явлений и содержащая информацию об отличительных признаках этих форм, представлена на рис. 1.
Заключение. Консортивная сущность симбиотических систем позволяет использовать в их отношении не только разработанную для консорций терминологию, но и ценоцентрическую систему отсчета.
В самом деле, симбиотическая система, как и всякая консорция, обладает всеми признаками биоценоза (сообщества), так как представляет собой совокупность живых организмов, связанных между собой биотическими связями, и образует вместе с окружающей ее неживой средой особую экосистему (по терминологии А.Ф. Зубкова [6], геоценоконсорцию). Ценоцентрическая система, однако, уместна только при описании внутренней структуры симбиотической системы. В отношении группировок, составленных однотипными симбиотическими системами, более уместна демоцентрическая система, однако отсчетным объектом в ней должна считаться популяция вида-эдификатора, а не демоценоз. Последнее особенно важно для случаев, когда консорты, входящие в одну и ту же популяцию, связаны с эдификаторами разной видовой принадлежности (как это бывает, например, у малоспециализированных насекомых-па-разитоидов).
Автор выражает искреннюю благодарность А.Б. Савинову (ННГУ) за чрезвычайно плодотворную дискуссию о симбиозе, способствовавшую написанию данной работы.
Список литературы Многообразие симбиотических отношений и возможный подход к их классификации
- Беклемишев В.Н. О классификации биоценологических (симфизиологических) связей//Бюлл. МОИП. Отд. биол. Т. 56. -1951, № 5. -С. 3-30.
- Власов Д.В. Фауна усачей Ярославской области: степень изученности и перспективы изучения/Биологические ресурсы, их состояние и использование в бассейне Верхней Волги. Сб. научн. тр. -Ярославль, 1999. -С. 220-229.
- Догель В.А. Курс общей паразитологии. Изд-е 2-е, дополненное. -Л.: Учпедгиз, 1947. -372 с.
- Догель В.А. Общая паразитология. -Л.: изд-во ЛГУ, 1962. -464 с.
- Емельянов А.Ф. О существенных различиях консорций доминантов и ассектаторов, проявляющихся в распределении цикадок-олигофагов по растениям//Ботан. журн. Т. 50. -1965, № 2. -С. 221-223.
- Зубков А.Ф. Биогеоценотические объект-элементы и подходы к их изучению//Экология. -1996, № 2. -С. 89-95.
- Лавренко Е.М. Основные закономерности растительных сообществ и пути их изучения//Полевая геоботаника. Т. 1. -М.-Л.: изд-во АН СССР, 1959.-С. 13-75.
- Мазинг В.В. Консорции как элементы функциональной структуры биоценозов//Тр. МОИП. -1966, т. 27. -С. 117-127.
- Матренина Р.М., Гродницкая И.Д. Роль экзометаболитов эпифитной микрофлоры лиственничной почковой галлицы в инициации аномального роста лиственницы сибирской//Лиственница. -1991. -С. 23-28.
- Озерский П.В. Что такое симбиоз?//Биология для школьников. -2008, № 3. -С. 23-34.
- Озерский П.В. О структуре теоретической экологии и месте в ней для аутэкологии//Функциональная морфология, экология и жизненные циклы животных. Вып. 9. -СПб: ТЕССА, 2009. -С. 11-21.
- Озерский П.В. Опыт классификации сред жизни как основы стаций и адаптивных зон//Функциональная морфология, экология и жизненные циклы животных. Вып. 10. -СПб.: ТЕССА, 2010. -С. 30-59.
- Проворов Н.А., Долгих Е.А. Метаболическая интеграция организмов в системах симбиоза//Журн. общ. биол. Т. 67. -2006, № 6. -С. 403-422.
- Раменский Л.Г. О некоторых принципиальных положениях современной геоботаники//Бот. журн. Т. 37. -1952, № 2. -С. 181-202.
- Ройтман В.А., Беэр С.А. Паразитизм как форма симбиотических отношений. -М.: КМК, 2008. -310 с.
- Савинов А.Б. Новая популяционная парадигма: популяция как симбиотическая самоуправляемая система//Вестник Нижегор. ун-та им. Н. И. Лобачевского. Сер. Биол. -2005, вып. 1. -С. 181-196.
- Харли Дж.Л. Биология микоризы // Микоризы растений // ред. Н.В. Лобанов. – М.: изд-во с.-х. лит-ры, журналов и плакатов, 1963. – С. 15–244.
- Шульман С.С., Добровольский А.А. Паразитизм и смежные с ним явления//Паразитол. сборник. -1977, т. 27. -С. 230-249.
- Bary A., de. Die Erscheinung der Symbiose. -Strassburg: Karl J. Trьbner, 1879. -30 s.
- Chilton M.-D., Drummond M.H., Merio D.J., Sciaky D., Montoya A.L., Gordon M.P., Nester E.W. Stable incorporation of plasmid DNA into higher plant cells: the molecular basis of crown gall tumorigenesis//Cell. Vol. 11. -1977, № 2. -P. 263-271.
- Dahl F. Grundsдtze und Grundbegriffe der biocцnotischen Forschung//Zool. Anz. Bd. 33. -1908, № 11. -S. 349-353.
- Dawkins R. Extended phenotype. -Oxford-NY: Oxford University Press, 1999. -317 p.
- Douglas A.E. Symbiotic interactions. -Oxford-NY-Toronto: Oxford Univ. Press, 1994. -148 p.
- Hirsch A. M., McFall-Ngai M. J. Fundamental concepts in symbiotic interactions: light and dark, day and night, squid and legume//J. Plant Growth Regulation. Vol. 19. -2000, № 2. -P. 113-130.
- Phares K. An unusual host-parasite relationship: the growth hormone-like factor from plerocercoids of spirometrid tapeworms//Int. J. Parasitol. Vol. 26. -1996, № 6. -P. 575-588.
