Модель распределения ассимилятов и оптимального роста растительного покрова

Автор: Воротынцев Александр Васильевич
Журнал: Вестник Бурятского государственного университета. Философия @vestnik-bsu
Рубрика: Математическое моделирование
Статья в выпуске: 9, 2015 года.
Бесплатный доступ
Формулируется модель роста биомасс четырех органов растительного покрова, сопряженная с моделью переноса тепла и влаги в покрове и почве с учетом баланса энергии. С помощью ростовых функций исследуется задача рационального распределения ассимилятов фотосинтеза и оптимального роста биомасс для максимизации конечного значения биомассы репродуктивного органа растения.
Моделирование, растительный покров, распределение ассимилятов
Короткий адрес: https://sciup.org/148183107
IDR: 148183107 | УДК: [574.4+631.43+631.6+626.8]:517.9 | DOI: 10.18097/1994-0866-2015-0-9-54-61
The model of assimilates allocation and optimal growth of plant cover
It was formulated the growth model of four plant organs biomass, conjugated with the model of heat and moisture transfer in plant cover and soil taking into account energy balance. Through the growth functions we studied the problem of photosynthesis assimilates rational distribution and optimal growth of biomass to maximize the final value of plant reproductive organ biomass.
Текст научной статьи Модель распределения ассимилятов и оптимального роста растительного покрова
Моделирование динамики роста биомассы органов растений совместно с переносом тепла и влаги в системе «почва – растительный покров» – одна из базовых задач описания функционирования агроценозов и их урожайности, [1; 5; 7]. Особый интерес представляют исследования механизмов адаптации растительных сообществ с помощью вариационных методов. В статье на основе модели роста биомасс растительного покрова в условиях дефицита влаги исследуется распределение ассимилятов фотосинтеза по органам растения, максимизирующее биомассу репродуктивных органов. Полученная стратегия распределения ассимилятов приводит к магистральному сбалансированному характеру роста биомасс, структурирующему и упрощающему модель.
1. Модель переноса влаги и тепла в растительном покрове
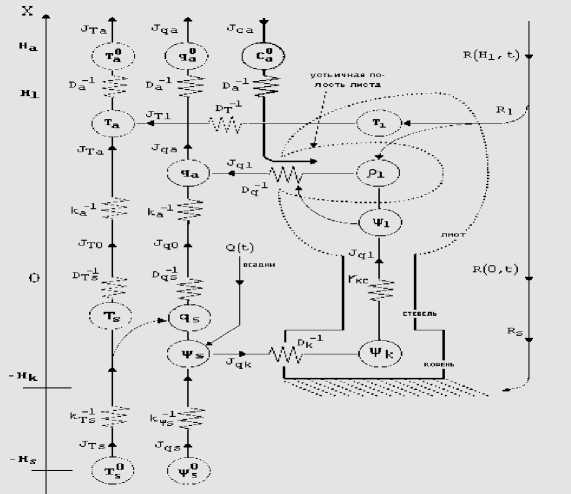
Рис. 1. Схема переноса тепла, влаги и CO 2 в РП
Приведем систему уравнений модели переноса, описанной в [1-5]. В слое 0 < x < Hl растительного покрова (РП) и в корнеобитаемом слое - Hs < x < 0 почвы рассматриваются две подсистемы уравнений (1)-(4) для температуры воздуха Ta , листьев Tl , концентрации водяного пара в межлистном воздухе qa , в устьичных полостях листьев ql :
JTa = - Cpka 3 Ta lxx > Jqa =- ka 3Ча I3x • 0 < x < Hl i
Cp STa/8t = -8JTa (8.x + /„ , 8Ча /51 = - 8J_/8x + f„ ,(2)
/„ = CpDTSl Tl- Ta )Pl , fql = DqSl (4i — Ча )Pl ,(3)
/ti + Xfqi =8R1I/8x ;(4)
и две подсистемы (5-7) для температуры Ts и водного потенциала v s < 0 почвы:
JTs =-cpkTsdTs/sx, J*, = -kVs a^s/5x, -Hs < x < 0(5)
Cs dTs/St = -dJTs /dx, S 9Vs/St = -dJvs /5x - V ,
V = JqiPk+ DkSk (Vs- Vs) Pk;
а также подсистема для концентрации углекислого газа CO 2
Jca =-ka 9Ca /5x ,(8)
5Ca (бt = -dJca dx + fd ,(9)
fl = DcSl (Cl - Ca )Pl ,(10)
с краевыми условиями:
JTa = CpDa (Ta - T," ) , Jqa = Da (qa - Ч^ ), x = Hl ,(11)
J Ca = D a ( C l - C a ) i
JTa = CpDTs (Ts - Ta ) , Jqa = Dqs (4s - 4a ) , x = 0 ,(12)
J c, = 0 ;
JTa + XJqa - JTs = R (0 ,t) , - Jvs + Jqa = Q (t), x = 0,(13)
Ts = Ts0, Vs = Vs, x = -Hs.(14)
Транспорт воды в растениях и испарение ими влаги Jql (транспирация) регулируется вод- ным потенциалом листьев vl, их устьичным сопротивлением rst и описывается нелинейными выражениями:
н 1 _
J ql = J fqldx , J ql = D k V VS - V l ) , при V m < V l < V s < 0, (15)
где Vs = JVsPkdx, Mi = Vl/Vm , 1 Dq = Vdt + rst , VDk = rkc +1 (DkSk) , - Hk rst = rs о(1 - §iMi)(1 - Mi)"*, 0 < Mi < 1, §i > 0; Ms = Vs lvm ; (16)
J ql = 0, при V s ^ V m < 0-
Подсистема уравнений, описывающая распределение углеводов, преобразование поглощенного CO 2 в углеводы, затем в биомассу органов и рост биомассы листьев (i=l), корней (i=r), репродуктивных органов (i=R) и стеблей (i=s), имеет вид [6,7]:
A mi = р z. (t )ф,А t — RT mA t- Ri A mi, или
Ф‘ mii =рi(t)-—~--Rmi, ^рi(t)=1, рi(t)^0, i^{l,r,R,s},
-
1 + R i
где
Ф; = SiтФ0 , R; = R'(Ti)(1 + Ri), A ^T, Ф0 = rTTA---ZT,(19)
(a I Ф ) + r c c l
Г = SlD- + D- , r = A1r + rc .(20)
Здесь JTa ( x,t ) , Jqa ( x,t ) , Jca ( x,t ) - потоки тепла, водяного пара и углекислого газа CO 2 в межлистном воздухе, JTs ( x,t ) , J V s ( x,t ) - потоки тепла и воды в почве; V s ( t ) — усредненный водный потенциал корнеобитаемого слоя почвы, v l ( t ) - водный потенциал листьев.
Продукты Ф'А t фотосинтеза распределяются по органам растения долями р i ( t ) и используются для роста A m i органов; при этом часть углеводов расходуется на дыхание поддержания R,i(Tl ) m i A t и дыхание роста R i A m i . Температурный фактор ^ ( T ) и константы Б ф , a , rc определяют влияние потока фотосинтетически активной радиации (ФАР) и концентрации cl на скорость Ф 0 преобразования CO 2 в ассимиляты единицей поверхности листа.
Дифференциальные уравнения систем связаны алгебраическими выражениями (4), (10), описывающими балансы энергии в межлистном воздухе и на поверхности почвы.
Введем обозначения:
da (х, t) = р(Ta) - q, ds (t) - p(Ts (0, t)) - qs (t) , da = p(Ta0)- q°(t), p(T)=p(Ti)+5(t - i), k (х) = ka(х) , G = ----177-^, G = G (1 - b), G, = G (1 - b);
-
1+ 5x / Cp 1 + Cp/М ' >>
-
1 = H d^ 1 -^XL = 1 + 1 = Hi A. 1 = 5x 1 = 1
Dak 1 ka (X) ' D)k c.D. ' 4 Dak Dak { kft) ' DTa cpD, ' D) DD
-
1 5x 1 5x 1 _ 1 1 _ 1 1 1 _ 11
, — , — , - , - + , - +
-
d t s C p D Ts D t C p D t S i D q D q S i Di D q d t D s D qs d t s
-
b i - G "‘ D , b s - G -1 D , D z- D a + D . + D S , D e - D ) + D T + D s .
2. Приближенная модель переноса влаги и тепла
Для модели (1)-(11) получим приближенную модель при условиях
D' Id ; << 1, D' id' << 1, ADy Id' << 1. ^ T ^ is ^ Hk
(22)
Здесь запись a << 1
математически эквивалентна a ^ 0; она используется
из-за своей боль-
шей физической содержательности.
Введем обозначения
в - ( D a + D s )/ d ; , b - D s /( d) + D s ) ;
(23)
d t - ( z - 1 gr) + D a d a 0+ D s d s )/( D a + D s ) ;
(24)
J qa - bJ + a ,
J qa - X - GR a + D a d a ;
(25)
J Ta - bJ - a + ( 1 - Ь ) R a ,
J ia - ( 1 - G ) R a 0 - x d) d a 0;
(26)
d a + - d t ,
d a - 0 ;
(27)
q - - q a + J - a 1 D a ,
q a + - q a 0 + b - 1 ( q a - q a 0 ) ;
(28)
Ttt- T a" + J- a ^pD) ) ,
. T a +- T a 0 + b ( t, -- T a 0 ) + ( 1 - b ) r) /( c p D a ) .
(29)
DT
Отметим, что введенные величины положительны, кроме, возможно, G l и G s . Очевидно, d ;/ d t < 1, D s/ d t s < 1.
В [5] доказано, что в условиях (22) справедлива система
J ,s -- k ,s d^ s/д х , c v -^ < 0, 5V s
( D' ;+ D s ) r o 1 -5 i p J + a 1 1 + D k ( F kc S k )
P -c — + A kc „ . — ; A kc - „ ,
Si 1 -Ps Sk 1 -Ps -VmDk при 0 < ps < 1, в - да при 1 < ps .
Проинтегрируем (30) на отрезке [ - Hk, 0 ] с учетом Jq) - Jql + Jq 0:
W + J qa - Q ( t ) - J v sHk - 0, w - f 9 w dx - H k 9 w , J , sHk - J , s I
-
- H k lX - H k
3. Модель фотосинтеза растительного покрова
Чтобы сделать нижеследующие оценки примем два допущения. Во-первых, примем, что из определения cv -59w /5^s для графика 9w -9w (vs) основной гидрофизической характеристи- ки (ОГХ) с достаточной точностью следует cV =50w /суs для зависимости средней влажности Оw от среднего потенциала Vs по слою [- Hk,0] • Дополнительно примем зависимость Vs = Ae~aw0w • Тогда (34), (33) преобразуется к виду:
d" Йs =аwЙs [Jqa (Йs ) —Q(t) —JvsHk ] , Vs = ЙsVm , aw = aH— , (35)
J qa (Й s ) = J qa 11^ , при 0 < M s < 1, J qa ( ц ' ) = bT ^ , при 1 <Ц ' • (36)
Получим приближенную модель фотосинтеза растительного покрова, предположив, что в системе уравнений (8)-(10): а) концентрации ca , c l , фотосинтетически активная радиация I ф незначительно изменяются внутри растительного покрова; б) покров находится в состояние квазистационарного равновесия в отношении cl , т.е. 5 cl/дt ~ 0.
С учетом сделанных предположений проинтегрируем (9)-(10) по высоте покрова на отрезке 0 < x < H l :
—
[ j ca l x = H
— J ca |x = 0 ]+ J 0 ' D c S l ( c l — c a ) P l dx = 0 •
С учетом (10)-(12) получим — D a ( c a — c 0 ) + DS ( c l — c a ) = 0. Выразив отсюда ca — c l и подставив в (10), получим выражение для потока CO 2 , поглощаемого листьями в единице объема РП:
fcl = —
D a D c S l
D a + D c S l
( c 0 — c l ) P l •
|
Интегрируя это в поглощенного стол CO 2 , поглощенног c l = c 0 — Г 1 Ф 0 в (19), |
ыражение на 0 < x < Hl , т.е. по высоте РП, получим величину Sl Ф 0 CO 2, бом РП с единичным основанием за единицу времени. Отсюда величина ) единицей поверхности листа, будет равна Ф 0 = ( c 0 — cl )/ r 1 . Подставляя получим для Ф 0 уравнение
|
|
Легко видеть, Ф нейшем для получе вать следующие бол |
0 ~ A 1 c0 / r при v ^ да , например, при I ф ^ да или r ^ да . Однако в даль-ния явных оценок в условиях дефицита почвенной влаги будем использо-ее сильные условия. D — = . r' 1 ( 1 6 1 Й s ) + A kc „’"„ S l . (38) 1 — Ц ' L r ' 0 D a + D ' S r J |
|
В самом деле, (38 |
) прямо следует из (33), (23), (22): e = ( D a + D ' ) ( 1 D ’ + 1 D T M D a + D ' )/ ( D q S l ) . |
|
Пусть r s<< D — 1 mes q |
<< r cut , S l D a 1<< D q 1 . Из (16) следует D q 1 = r 't + D T 1 ~ r 't , D —1 = V ( r ' — 1 + r cut ) + r mes ~ D q' , r 1 = S l D a 1 + D — « D — « D q1 . (39) |
|
Таким образом, п |
ри выполнении приведенных выше приближенных равенств (39) Ф 0 « c " jrx « c " D q . (40) |
|
Введем обозначе |
ния |
K- m k- , S i -p m , S k =P A , ml
B 1 - 1 -8 i ц s , B 2 =
A kc J qa P l
r s о D ‘+ D ’e k
в l T c a ~ ~
, Ф 1 -^1+^ ’ R i = R ,
G (X , H s ,t ) = ^TT(1 — ^ s ) , F ( X , ^ s ,t ) = G Yr s ^Ф 1
B ^ K + B 2 K + 1
Уточним (40):
r ~ D - - r s о ( 1 -ц s ) - 1 ( B 1 K + B 2 ! л ,
Ф' S,TC° x h ~ - A ~ Y - mlФ^(K,цs,t).
( 1 + R i ) ( 1 + R i r
Для приближения (40) модели фотосинтеза выпишем модель роста четырех биомасс m i ( t ) растительного покрова и сопряженную с ней модель динамики усредненного водного потенциала ц s корнеобитаемого слоя почвы:
d-m, - рi(t)mlG(K,цs,t)Ф1 -Rmi , i e {l.k.R.s} ;(43)
dt d Цs -a w цs [Jqa (u s )-Q (t )-J„sHk ] , JqQ (цs )-J+ ’ ’ ^^ ;
t+ U
Spi (t)- 1 , Pi (t)-0 , P- (D д Ds -rs 0A , ^s -Цs ^ m , 0 < Ms < 1;
m i p i G (K , ц s i )
m i ( 0 )- m 0, ц s ( 0 )-ц 0 .
-
2. Рациональное распределение ассимилятов
Рост биомасс m i ( t ) в (43) зависит от значений ростовых функций р i ( t ) , моделирующих сложные механизмы распределения и транспорта ассимилятов в растении. Обычно функции р i ( t ) определяются натурными измерениями.
Варьируя р i ( t ) можно в значительной степени регулировать рост всех биомасс m i ( t ) и, следовательно, величину биомассы mR ( t ) репродуктивного органа в конце жизненного цикла растения [ 0 ,Т ] . Представляет интерес определить такие р i ( t ) , при которых mR ( t ) достигает максимума. Разумно пр едполагать, что такая структура распределения ассимилятов может возникать эволюционно у некоторых видов растений, например злаковых, подвергаемых отбору на максимальную урожайность репродуктивных органов.
Рассмотрим задачу выбора функций рi (t), максимизирующих для системы (43)-(45) на [0,Т] функционал mR (Т) ^ max (46)
при ограничениях (45). Для частного случая, когда параметры внешней среды постоянны, в частности постоянны водный потенциал ц s и температура листьев T l , в [6] доказана теорема о магистрали для m i ( t ) из (43). Опишем содержательную интерпретацию результатов формальной теоремы, представленных на рис. 2 и 3, где m 1 - m l , m 2 - mk , m 3 - mR . Заметим, что оптимальные р s ( t ) - 0, ибо из-за отсутствия в модели (43)-(45) учета архитектоники растения затраты ассимилятов на рост биомассы ms стеблей оказываются непродуктивными. Также нетрудно видеть, что затраты ассимилятов на рост mR продуктивны лишь на заключительном репродуктивном периоде [ т 2 ,Т ] . Для начального вегетационного периода T 1 < t <т 2 рассмотрим функцию F ( K , ц s,t ) удельной фотосинтетической мощности покрова, приходящейся на единицу суммарной биомассы
F ( K , ц s t ) --^^ G ( K , ц s ,t ) Ф 1 (47)
m l + m k
Функция F имеет единственный максимум при X * = ^B\ /В 1 - решении уравнения
_L F=Ф (1 _й )d X -в^2+в2
dX Л Цs’d (X +1)(B1X + B2) (X +1)2(B1X + B2)2
равный F* = Ф1 (1 - цs )(тв"+вг- )г2 . Оптимальные р* (t) и рк (t), отвечающие F*, есть реше- ния системы ml = рlmlGФ1 - Rml , mk = рkmkGФ1 - Rmk , mk = X*ml, рl + рk = 1 .
Отсюда следует
1 X *
X = V B 2/ в 1 , р / =Г^ , р к =r"v , P R = 0 ’ P s = 0, t G [ T ’ T 2 ) • (48)
1+ X 1+ X
Итак, период роста [ 0 ,T ] разбивается на три фазы: начальную [ 0 , т 1 ) , вегетативную [ т 1 , т 2 ) и репродуктивную [ т 2 ,Т ] . На начальной фазе система стремится достичь сбалансированного отношения биомасс корней и листьев X * , направляя все ассимиляты на рост органа с недостаточной биомассой. Например, начальное значение X 0 > X * на рис. 3 характеризуется недостаточной биомассой листьев ml , поэтому все ассимиляты направляются на рост листьев, т.е,
р l = 1, р к = 0, р R = 0, р s = 0, 0 ^ t ^т1
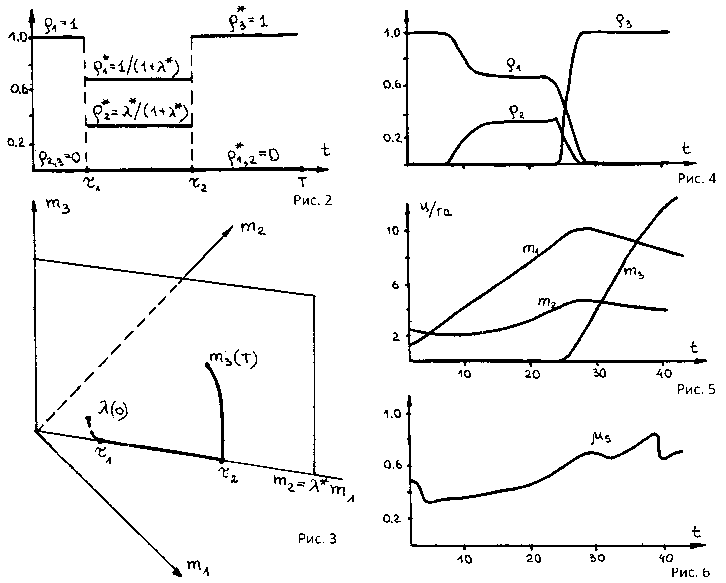
Рис. 2–6. Оптимальное решение модели (43)–(46)
В момент т1 отношение X(t) становится равным X*, система достигает магистрали m2 =X*m1 и переходит в состояние сбалансированного роста. Это - вегетативная фаза, в течение которой выполняется (48). Скорость роста зависит от условий среды, в частности от усред- ненного водного потенциала цs.
С момента т2 начинается третья репродуктивная фаза, при которой все ассимиляты направляются на рост репродуктивной биомассы mR р1 = 0, рк = 0, рR = 1, рs = 0, т2 < t < T .
В момент времени T система заканчивает рост с максимальным значением mR ( Т ) . На рис. 4–6 показаны численные решения задачи (43)–(46).
Таким образом, оптимальное сбалансированное распределение ассимилятов соответствует максимизации удельной мощности фотосинтеза F(X,цs,t). Траектория магистрали обладает эффектом «притяжения» оптимальных траекторий с другими значениями начальных и внешних условий. Можно ожидать, что использование магистрали позволит лучше структурировать модель растительного покрова.
Список литературы Модель распределения ассимилятов и оптимального роста растительного покрова
- Полуэктов Р.А. Динамические модели агроэкосистем. -Л.: Гидрометеоиздат, 1991. -312 с.
- Воротынцев А.В. Модель переноса тепла и влаги в системе «почва -растение» с учетом баланса энергии//Вестник Бурятского государственного университета. -2012. -№ 1. -С. 107 -111.
- Воротынцев А.В. Исследование модели водно-теплового режима системы почва -растение-приземный слой воздуха//Сообщения по прикладной математике. -М.: Вычислительный центр РАН, 1991. -53 с.
- Воротынцев A.B. Исследование модели переноса тепла и влаги в системе почва-растение//Математическая биология и биоинформатика. Спецвыпуск по итогам конференции ЭкоМатМод-2011. -2012. -Т. 7, № 1. -С. 45-53.
- Воротынцев A.B. Приближенная модель переноса тепла и влаги в системе почва-растение с учетом баланса энергии.//Вестник Бурятского государственного университета. -Вып. 9(1) Математика, информатика. -2014. -С. 22-26.
- Две задачи оптимального управления для модели системы «почва -растение»/А.В. Воротынцев, Н.Л. Рогожина, И.А. Соколова, С.О. Степченкова. -М.: Вычислительный центр АН СССР. Сообщения по прикладной математике, 1988. -38 с.
- Бихеле З.H., Молдау X.А., Росс Ю.К. Математическое моделирование транспирации и фотосинтеза растений при недостатке почвенной влаги. -Л.: Гидрометеоиздат, 1980. -223 с.
