Моделирование динамики межфазного натяжения на границе углеводород - вода при образовании кластеров актинобактерий

Автор: Куюкина М.С., Феоктистова Е.В., Осипенко М.А., Ившина И.Б., Няшин Ю.И.
Журнал: Российский журнал биомеханики @journal-biomech
Статья в выпуске: 1 (83) т.23, 2019 года.
Бесплатный доступ
Одним из загрязняющих факторов окружающей среды являются нефть и нефтепродукты, ежегодные мировые выбросы которых составляют десятки тысяч тонн. Проблема нефтезагрязнения водных ресурсов важна вследствие ведущейся шельфовой добычи нефти и перевозок нефтепродуктов морским и речным транспортом. Эффективным способом ликвидации нефтяных загрязнений водной среды является биоремедиация с помощью углеводородокисляющих микроорганизмов, в частности актинобактерий рода Rhodococcus . В работе исследована динамика адсорбции клеток Rhodococcus ruber ИЭГМ 231 на границе, разделяющей бактериальную суспензию и жидкий углеводород ( н -гексадекан либо н -додекан), и выполнено математическое моделирование образования биопленки на основе описания процессов формирования и разрушения клеточных кластеров. Разработана феноменологическая биомеханическая модель изменения поверхностного натяжения; она описывает соответствующие экспериментальные зависимости, полученные с помощью высокоточной тензиометрии при разной оптической плотности бактериальной суспензии. Разработана также микроуровневая модель, основанная на теории насыщения слоя бактерий на поверхности раздела фаз и образования единой биопленки на этой поверхности. В этой модели предложено теоретическое объяснение скачкообразной динамики межфазного натяжения, обусловленной прохождением измерительного кольца через формирующуюся биопленку и случайным характером формирования-разрушения клеточных кластеров до достижения стабильного состояния системы.
Бактериальные клетки, биопленки, поверхностное (межфазное) натяжение, детерминистическая модель, вероятностная модель
Короткий адрес: https://sciup.org/146282117
IDR: 146282117 | УДК: 531/534: | DOI: 10.15593/RZhBiomeh/2019.1.05
Modelling of surface tension dynamics at the hydrocarbon - water interface during formation of actinobacterial clusters
One of the main factors of environmental pollution is oil and petroleum products, which annual global emissions resulting from multiple accidents and leaks amount tens of thousands of tons. The problem of oil pollution of water resources has acquired a threatening scope due to a growing offshore oil production and transportation of petroleum products by sea and river transport. An effective way to eliminate the oil pollution of water environments is bioremediation using hydrocarbon-oxidizing microorganisms, in particular actinobacteria of the genus Rhodococcus . We studied the adsorption dynamics of Rhodococcus ruber IEGM 231 cells at the interface between bacterial suspension and liquid hydrocarbon ( n -hexadecane or n -dodecane). As a result, mathematical modelling of the initial stage of biofilm formation was performed based on thermodynamic description of cell cluster formation and destruction. The first developed biomechanical model of the surface (interfacial) tension time dependence is phenomenological, it describes the experimental graphs of the corresponding dependence obtained by high-precision tensiometry at different optical densities of bacterial suspensions. The second microlevel model is based on the theory of saturation of the bacterial adsorption layer and the biofilm formation at the hydrocarbon-water interface. In this model, a theoretical explanation of the revealed oscillatory dynamics of interfacial tension is proposed, thus relating to the measuring ring passages through the formed biofilm and the random nature of cell cluster formation-destruction to achieve thermodynamically stable state of the system.
Текст научной статьи Моделирование динамики межфазного натяжения на границе углеводород - вода при образовании кластеров актинобактерий
Актинобактерии рода Rhodococcus (рис. 1) являются представителями важной эколого-трофической группы микроорганизмов, окисляющих природные и антропогенные углеводороды и, следовательно, участвующих в биогеохимических циклах углерода и восстановлении загрязненных углеводородными ксенобиотиками экосистем [2]. Родококки способны к окислительной деструкции практически всех углеводородных компонентов нефти: алканов, циклических и ароматических углеводородов и их производных, сложных поли- и гетероциклических соединений. Отдельные представители рода Rhodococcus используются в составе биопрепаратов, предназначенных для биоремедиации нефтезагрязненных сред [12].
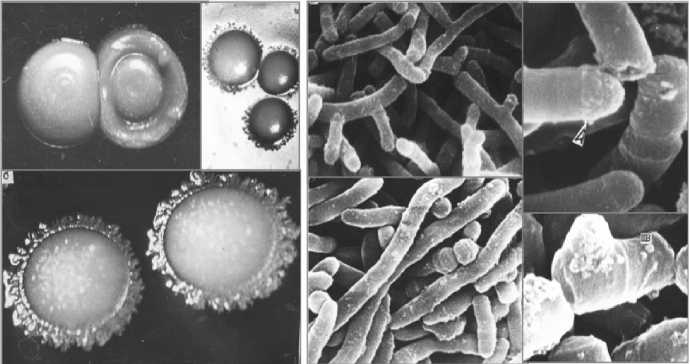
а б
Рис. 1. Колонии ( а ) и клетки ( б ) родококков [2]
Начальным этапом процесса биодеструкции углеводородных соединений является бактериальная адгезия, интенсивность которой определяется гидрофобностью клеток и их адгезивной активностью. Бактериальная адгезия к жидким углеводородам рассматривается отдельными авторами [13, 16] как механизм образования биопленок в водно-углеводородных средах. Например, в работе [13] исследована динамика комплексного процесса пленкообразования, сопряженного с биодеструкцией полихлорированных бифенилов смешанной культурой микроорганизмов. В ряде исследований описаны математические модели бактериальной адгезии к твердым субстратам [9, 17], формирования биопленок на границе твердый субстрат – жидкость [7, 8] и жидкая среда – воздух [6], а также процессов биофильтрации газообразных выбросов и очистки сточных вод от углеводородных загрязнителей с применением биопленок [1, 3]. Однако в литературе отсутствуют модели процесса биопленкообразования в двухфазной системе несмешивающихся жидкостей.
С использованием метода высокоточной межфазной тензиометрии исследована термодинамика адгезионного процесса и экспериментально подтвержден сорбционный механизм формирования биопленок родококками в водно-углеводородной системе [4, 5]. Цель настоящей работы – математическое моделирование процессов формирования и разрушения клеточных кластеров Rhodococcus ruber ИЭГМ 231 на основе экспериментальных данных по динамике межфазного натяжения на границе, разделяющей бактериальную суспензию и жидкий углеводород ( н -гексадекан либо н -додекан).
Экспериментальное исследование динамики межфазного натяжения в системе углеводород – бактериальная суспензия
Определение поверхностного (межфазного) натяжения в системе углеводород– вода проводили методом отрыва кольца на высокоточном автоматическом тензиометре Sigma 701 ( KSV Instruments , Финляндия) при температуре 25 ± 0,2 оС в течение 15–24 ч. Регистрацию результатов измерения осуществляли с использованием программного обеспечения « WinSigma » ( KSV Instruments , Финляндия). В работе использовали штамм Rhodococcus ruber ИЭГМ 231 из Региональной профилированной коллекции алканотрофных микроорганизмов (акроним ИЭГМ, WDCM # 768; www.iegm.ru/iegmcol/strains.html ). Готовили водные суспензии отмытых от питательной среды клеток родококков в различных концентрациях, соответствующих показателям оптической плотности (ОП 600нм ) 0,25–2,0 (ОП – безразмерная величина, характеризующая степень погашения света, прошедшего через слой материала), измеряемым с помощью спектрофотометра Lambda EZ 201 ( Perkin-Elmer , США) при длине волны 600 нм. В качестве контроля использовали систему вода – н -гексадекан ( н -додекан) без добавления клеток.
Результаты долгосрочных экспериментов по определению динамики межфазного натяжения в системе вода – н -гексадекан при различных концентрациях клеток в суспензии представлены на рис. 2. Как видно из рис. 2, равномерное снижение показателя межфазного натяжения до стабильного минимального значения наблюдалось при низкой (ОП 600нм = 0,5) концентрации бактериальной суспензии. В то же время при средних (0,75–1,0) значениях ОП 600нм был зарегистрирован скачкообразный характер кривой межфазного натяжения после 4,5–6 ч взаимодействия клеток с н -гексадеканом, что, по-видимому, свидетельствует о постепенной стабилизации процесса адсорбции родококков в межфазном слое. Такое же скачкообразное колебание межфазного натяжения клеточной суспензии при
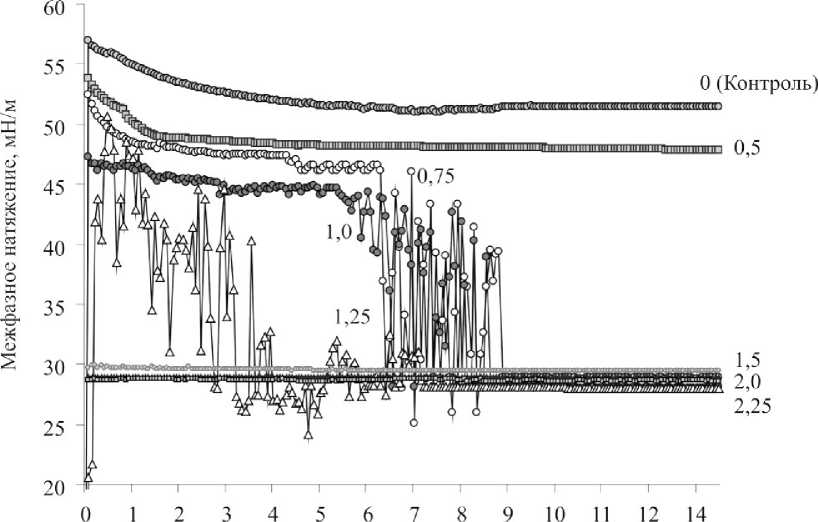
Время, ч
Рис. 2. Динамика межфазного натяжения суспензии клеток R. ruber ИЭГМ 231 в системе вода – н -гексадекан. Значения ОП600нм клеточных суспензий: 0; 0,5;
0,75; 1,0; 1,25; 1,5; 2,0; 2,25
ОП 600нм = 1,25 наблюдалось уже с первых минут в течение 8 ч эксперимента. При этом после 9 ч взаимодействия бактериальных клеток с н -гексадеканом показатели межфазного натяжения приобретали стабильный характер (28–30 мН/м), что указывает на предельное насыщение межфазного слоя клетками родококков и формирование межклеточных контактов. Стабильно низкие значения межфазного натяжения регистрировались на протяжении всего эксперимента при использовании высококонцентрированных (ОП 600нм = 1,5–2,25) клеточных суспензий.
Для исследования влияния природы углеводородного субстрата на динамику процесса адгезии клеток в двухфазной системе была проведена серия аналогичных экспериментов с н -додеканом (рис. 3), характеризующимся меньшей гидрофобностью, чем н -гексадекан, и, соответственно, большей растворимостью в воде. Как видно из рис. 3, в системе с н -додеканом наблюдалась сходная динамика межфазного натяжения в зависимости от концентрации клеток родококков. Скачкообразный участок кривой межфазного натяжения с последующим достижением минимального значения (27 мН/м) регистрировался при наиболее низкой (ОП 600нм = 0,25) исследуемой концентрации бактериальной суспензии. При использовании суспензии клеток в концентрации, соответствующей ОП 600нм = 0,5, не удалось зарегистрировать стабильных показателей межфазного натяжения на протяжении всего эксперимента. Стабилизация значений межфазного натяжения на минимальном уровне в системе с н -додеканом происходила при меньшей концентрации клеток (ОП 600нм = 1,25) по сравнению с таковой в опытах с н -гексадеканом.
Полученные результаты свидетельствуют о том, что родококки на границе раздела фаз вода – углеводород проявляют свойства поверхностно-активных частиц, сорбируясь на межфазной поверхности и понижая межфазное натяжение системы в концентрационно-зависимой манере до достижения минимального значения, соответствующего предельной сорбции клеток. При этом в насыщенном сорбционном слое клеток происходят образование межклеточных контактов и инициация формирования бактериальной биопленки. Инициация формирования биопленки
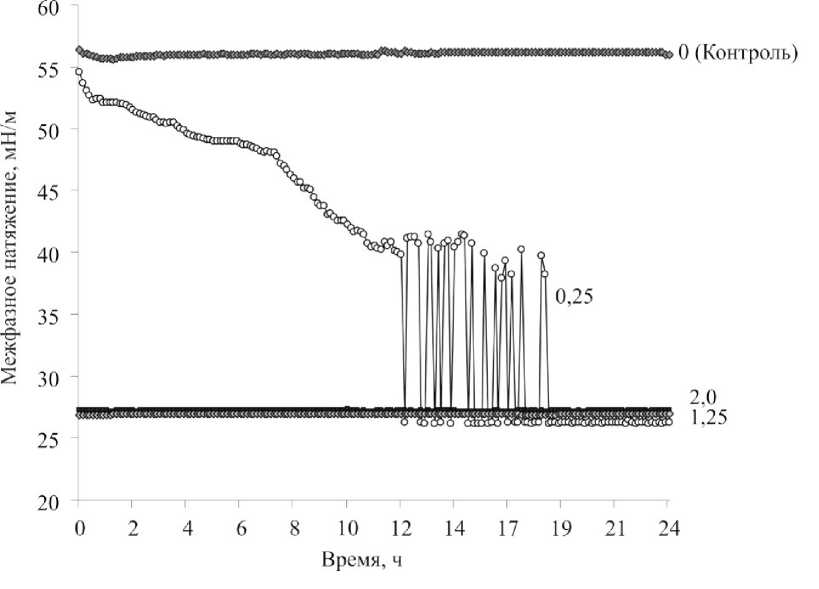
Рис. 3. Динамика межфазного натяжения суспензии R. ruber ИЭГМ 231 в системе вода – н -додекан. Значения ОП600нм клеточных суспензий: 0; 0,25; 1,25; 2,0
в системе вода – н -додекан начинается при меньших концентрациях клеток в суспензии, что, по-видимому, обусловлено термодинамическими свойствами данной системы. Вероятно, площадь раздела фаз возрастает с увеличением дисперсности системы, поэтому площадь контакта клеток с н -додеканом выше по сравнению с таковой в системе с н -гексадеканом, и клетки родококков за более короткое время адсорбируются на доступной поверхности. Замечено, что стабильные показатели межфазного натяжения клеточной суспензии были примерно одинаковы (27–30 мН/м) независимо от природы углеводородного субстрата. Таким образом, снижение показателей межфазного натяжения в двухфазной системе обусловлено участием гидрофобных взаимодействий между бактериальными клетками и углеводородом. Полученные экспериментальные данные могут быть использованы для регуляции роста бактериальных биопленок в жидких гетерофазных системах.
Детерминистическая биомеханическая модель зависимости ПОВЕРХНОСТНОГО НАТЯЖЕНИЯ ОТ ВРЕМЕНИ
Обозначим через n(t) временную зависимость концентрации клеток на межфазной границе. Будем считать, что эта зависимость имеет стандартный релаксационный вид n (t) = П1 (1 - exp (-t^i)), где n1 – максимально достижимая концентрация, t1 – некоторое характерное время. Зависимость поверхностного натяжения а от концентрации предполагаем имеющей вид
a(n) = а0 -(а0 - а1(s(n)) ) —, n1 где a1(s) = a10 -(а10 -а11)s , a0, aw, ап - некоторые характерные значения a, а величину 0 < s < 1 можно назвать степенью образования единой пленки [13, 16, 17] на межфазной границе. Для зависимости s(n) примем следующую феноменологическую формулу:
, х 11 . n - — s (n) = — + — arctg ----2 ,
2 n ^ n 3 v где n2 и n3 – некоторые характерные значения n.
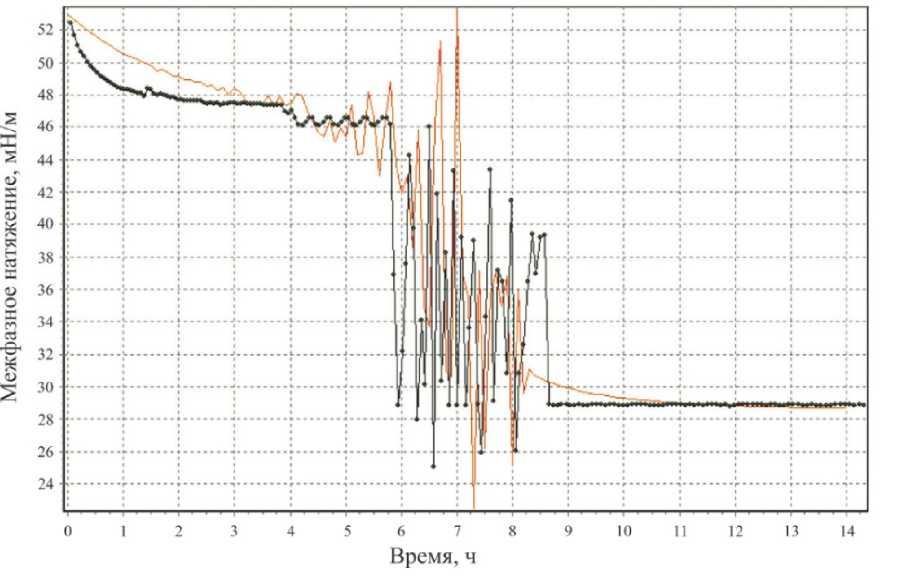
Сравнивая получившуюся в итоге теоретическую зависимость a ( t ) (без учета случайных вариаций) с экспериментальной зависимостью при ОП 600 нм = 0,75, находим методом наименьших квадратов параметры t 1 , a 0 , a 10 , an, n 2 , n 3 . Теоретическая и экспериментальная зависимости сопоставлены на рис. 4, а .
Вероятностная биомеханическая модель зависимости ПОВЕРХНОСТНОГО НАТЯЖЕНИЯ ОТ ВРЕМЕНИ
Вероятностная модель учитывает случайный характер процесса образования единой пленки бактерий. Пусть степень образования этой пленки есть случайная величина, равномерно распределенная на отрезке [ s ( n ) - Д ( n ), s ( n ) + Д ( n ) ] , где
Д ( n ) = k ( n )min( s ( n ), 1 — s ( n )) , k ( n ) = <
'1,7 0
( n < 0,985),
( n > 0,985)..
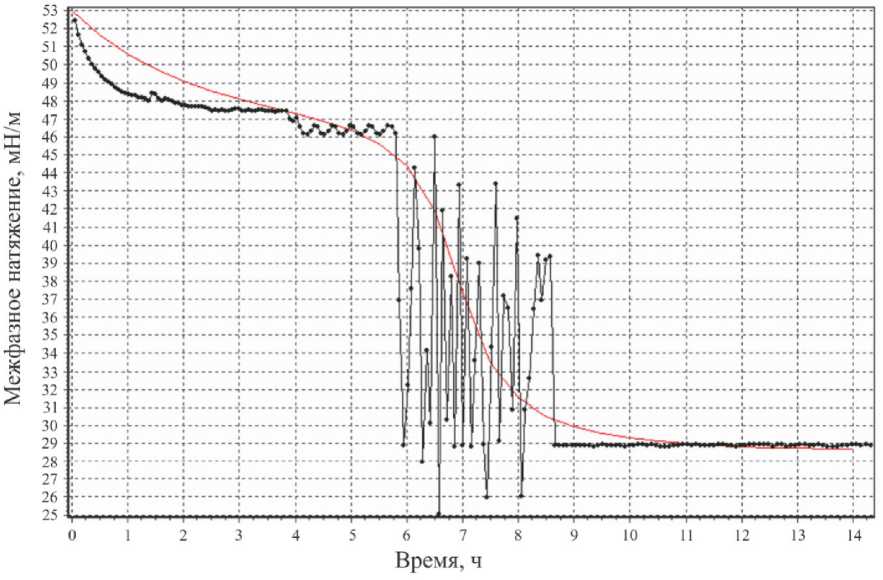
а
б
Рис. 4. Экспериментальная зависимость σ( t ) (ОП600нм = 0,75) и соответствующая теоретическая зависимость, не учитывающая ( а ) и учитывающая ( б ) случайные вариации
Получившаяся теоретическая зависимость (одна из реализаций вышеупомянутой случайной величины) и экспериментальная зависимость при ОП 600 нм = 0,75 сопоставлены на рис. 4, б .

Рис. 5. Пример кластера: а – скопление бактерий; б – наибольший кластер
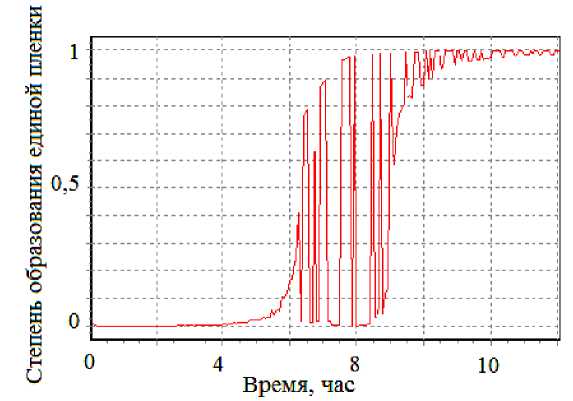
Рис. 6. Зависимость s ( t ) в модели формирования и разрушения кластеров
Далее вероятностная модель была дополнительно усложнена. Феноменологические формулы для s(n) и Δ(n) получили объяснение на уровне следующей микромодели. Предполагалось, что по мере накопления клеток на межфазной границе они объединяются в кластеры [10, 11, 15]. Моделирование кластерообразования проводилось по схеме клеточных автоматов. Если одна из соседних клеток данной клетки входит в уже имеющийся кластер, то в него включается и данная клетка. Кластеров может быть много, и среди них можно выделить наибольший (рис. 5). Доля клеток, входящих в наибольший кластер, растет со временем, при этом зависимость этой доли от концентрации клеток качественно соответствует приведенной выше феноменологической зависимости s(n). Поэтому можно считать, что доля клеток в наибольшем кластере и есть степень s образования единой пленки. Тогда отпадает необходимость в использовании искусственной феноменологической зависимости s(n). Далее случайные вариации величины s могут быть объяснены следующим образом. Измерительное кольцо, проходя межфазную границу, разрушает встретившийся кластер с некоторой вероятностью [7, 14]. Если наибольший кластер еще не очень велик, то к следующему проходу измерительного кольца он не успевает восстановиться, и величина s остается малой, а поверхностное натяжение – большим. С течением времени наибольший кластер растет и поэтому успевает восстановиться к следующему проходу кольца; тогда величина s устанавливается близкой к единице и поверхностное натяжение становится малым. В промежуточном случае наблюдаются случайные колебания значений s. В итоге отпадает необходимость в использовании искусственной феноменологической зависимости Δ(n), а зависимость s(t) имеет вид, показанный на рис. 6, т.е. совпадает с использованной при получении теоретической зависимости на рис. 4, б. Тем самым эта зависимость сохраняется, но объяснена теперь посредством микромодели формирования и разрушения кластеров бактерий.
Заключение
Проведено экспериментальное и теоретическое исследование динамики снижения межфазного натяжения в результате адсорбции клеток R. ruber ИЭГМ 231 на границе, разделяющей бактериальную суспензию и жидкий углеводород ( н -гексадекан либо н -додекан). Выполнено математическое моделирование начального этапа образования биопленки на основе математического описания процессов формирования и разрушения клеточных кластеров. Установлено, что экспериментально наблюдаемая зависимость межфазного натяжения в системе бактерии – вода – углеводород от времени носит нерегулярный характер, поэтому детерминистическая модель может лишь качественно описать эту зависимость. Более точное соответствие между теорией и экспериментом может быть достигнуто с помощью вероятностной модели. Разработанная микроуровневая модель основана на теории насыщения адсорбционного слоя бактерий на поверхности раздела фаз и образования единой биопленки на межфазной границе углеводород – вода. В этой модели предложено теоретическое объяснение выявленной скачкообразной динамики межфазного натяжения, обусловленной прохождением измерительного кольца через формируюшуюся биопленку и случайным характером формирования – разрушения клеточных кластеров до достижения термодинамически стабильного состояния системы. Эта модель качественно и количественно описывает основные черты рассматриваемого явления. Модель допускает дальнейшее развитие, которое должно состоять в более подробном анализе механизмов поведения бактериальных клеток на границе раздела фаз.
Благодарности
Исследования выполнены в рамках госзаданий 6.3330.2017.4.6 (ПГНИУ), 01201353246 (ИЭГМ УрО РАН) и поддержаны грантом РФФИ 18-29-05006.
Список литературы Моделирование динамики межфазного натяжения на границе углеводород - вода при образовании кластеров актинобактерий
- Дик И.Г., Пыльник С.В., Миньков Л.Л. Моделирование эволюции водоочищающей биопленки с учетом ее эрозии // Биофизика. - 2005. - Т. 50, № 3. - С. 505-514.
- Ившина И.Б., Пшеничнов Р.А., Оборин А.А. Пропанокисляющие родококки. - Свердловск: УНЦ АН СССР. - 1987. - 125 с.
- Митин А.К., Николайкина Н.Е., Загустина Н.А. Моделирование процесса биологической очистки газа // Вестник Тамбовского государственного технического университета. - 2016. - Т. 22, № 1. - С. 84-93.
- Николаев Ю.А., Плакунов В.К. Биопленка - «город микробов» или аналог многоклеточного организма? // Микробиология. - 2007. - Т. 76, № 2. - С. 149-163.
- Рубцова Е.В. Адгезия клеток родококков к жидким углеводородам и их производным: дис. … канд. биол. наук. - Пермь, 2011. - 188 с.
