Молекулярно-генетический анализ останков людей из погребального комплекса эпохи бронзы Бертек-56 (II тыс. до н.э., Республика Алтай, Россия)
")
Автор: Пилипенко А.С., Молодин В.И., Трапезов Р.О., Черданцев С.В., Журавлев А.А.
Журнал: Археология, этнография и антропология Евразии @journal-aeae-ru
Рубрика: Антропология и палеогенетика
Статья в выпуске: 4 т.44, 2016 года.
Бесплатный доступ
В статье представлены результаты палеогенетического исследования останков двух индивидов (взрослого и ребенка), представляющих население Горного Алтая эпохи развитой бронзы, из погребального памятника Бертек-56 (плато Укок, Республика Алтай, Россия) и их интерпретация с учетом археологического и палеоантропологического контекста. Были исследованы четыре системы генетических маркеров: митохондриальная ДНК, полиморфный фрагмент гена амелогенина, аутосомные STR-локусы и STR-локусы Y-хромосомы. Для взрослого индивида получен полный объем данных по этим системам, а для ребенка - по структуре мт ДНК, гену амелогенина и частичные профили аутосомных STR-локусов и STR-локусов Y-хромосомы. Генетические данные позволили определить мужской пол обоих погребенных. Установлено отсутствие прямого родства между индивидами. Филогенетический анализ показал принадлежность митохондриальной ДНК ребенка к западно-евразийской гаплогруппе K (подгруппа K1a24a), взрослого - к восточно-евразийской гапло-группе C. С помощью программы-предиктора было установлено, что исследованный гаплотип Y-хромосомы мужчины (17 STR-локусов Y-хромосомы) относится к гаплогруппе Q, которая является характерной для популяций Восточной Евразии. Филогенетический и филогеографический анализ полученных палеогенетических данных свидетельствует в пользу происхождения исследуемой популяции Горного Алтая эпохи развитой бронзы в результате смешения двух генетически контрастных групп (с восточно-евразийскими и западно-евразийскими генетическими характеристиками), что согласуется с выводами археологов и антропологов об участии в формировании группы, оставившей памятник Бертек-56, как автохтонного населения региона, так и пришлой популяции из Западной Евразии.
Палеогенетика, древняя днк, митохондриальная днк, маркеры половой принадлежности, str-маркеры, y-хромосома, горный алтай, плато укок, эпоха бронзы
Короткий адрес: https://sciup.org/145145780
IDR: 145145780 | УДК: 575.17 | DOI: 10.17746/1563-0102.2016.44.4.141-149
Текст научной статьи Молекулярно-генетический анализ останков людей из погребального комплекса эпохи бронзы Бертек-56 (II тыс. до н.э., Республика Алтай, Россия)
Погребально-поминальный комплекс Бертек-56, расположенный на плато Укок в Республике Алтай (рис. 1), был исследован западно-сибирским отрядом

Рис. 1. Плато Укок.
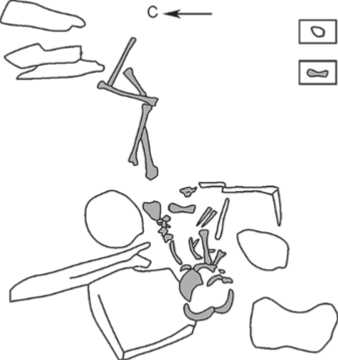
Северо-Азиатской комплексной экспедиции ИАЭТ СО РАН под руководством В.И. Молодина летом 1992 г. в рамках международной программы «Пазы-рык» [Молодин, 1993]. Памятник представляет собой каменное кольцо диаметром 14 м с каменной наброской в центре (рис. 2, 3), под которой обнаружены два погребения на уровне материка. Погребенные находились в каменных «цистах», которые состояли из длинных боковых стенок (торцевые отсутствовали), перекрытых массивными плитами [Молодин и др., 2004, с. 205–206]. Они были уложены в вытянутом положении на правом боку, головой на восток (рис. 4, 5). Отсутствие некоторых костей скелетов свидетельствует о возможном вторичном характере погребений. Согласно палеоантропологическим определениям, погр. 1 содержало останки ребенка 8–9 лет, погр. 2 – взрослого мужчины, умершего в возрасте 30–35 лет [Чикишева, 2012, с. 129–131].
Обнаруженные в комплексе Бертек-56 фрагменты трех керамических сосудов позволяют уверенно
Рис. 2. Курган Бертек-56 после расчистки конструкции.

0 20 cм
Рис. 4. Погребение 1.
– каменная конструкция; 2 – кости человека.
0 120 cм
Рис. 3. План кургана.
1 – кострище; 2 – погребение 1; 3 – погребение 2.
датировать его периодом развитой бронзы (II тыс. до н.э.). Ранее неоднократно отмечалось своеобразие культурного контекста этого комплекса [Молодин, 1993; Молодин и др., 2004, с. 206; Молодин, 2006], что дает основания для выведения памятника за рамки синхронной ему каракольской культуры Горного Алтая.
Исследование погребенных методами физической антропологии позволило установить наличие краниотипа, в целом тяготеющего к европеоидному. При этом как особенности краниотипа, так и комплекс одонтологических признаков свидетельствуют об их происхождении в результате смешения носителей двух разных морфологических комплексов [Чики-шева, 2012, с. 129–131] (см. раздел «Результаты и обсуждение»).
Современный уровень развития методов палеогенетики потенциально дает возможность получать достоверную информацию о генетических корнях и связях представителей древнего населения, половой принадлежности останков, степени родства индивидов в парных и коллективных погребениях. Высокая степень сохранности скелетных останков из памятника Бертек-56 позволила выполнить
Рис. 5. Погребение 2.
1 – каменная конструкция; 2 – кости человека.
разносторонний молекулярно-генетический анализ погребенных (маркеры половой принадлежности, филогенетически и филогеографически информативные однородительские маркеры (мтДНК и Y-хромосома), маркеры степени генетического родства). В данной ра-
боте мы приводим результаты этого анализа и интерпретацию полученных палеогенетических данных в археологическом и палеоантропологическом контексте.
Материалы и методы
Палеоантропологические образцы. Для молекулярногенетического исследования были взяты кости посткраниального скелета, характеризующиеся наибольшей макроскопической сохранностью: для индивида 1 (ребенок) – бедренная, для индивида 2 (взрослый) – большая берцовая. Следует отметить, что костный материал взрослого мужчины отличался высокой степенью сохранности, а ребенка, напротив, демонстрировал признаки, косвенно указывающие на более слабую сохранность ДНК в нем (низкая плотность и пористость компактного костного вещества, изменение его цвета).
Для исключения перекрестной контаминации между образцами индивидов 1 и 2 экспериментальные работы с материалами были разделены хронологически.
Предварительная обработка палеоантропологического материала и экстракция ДНК. Использовались методы, описанные в наших работах [Pilipenko, Romaschenko, Molodin et al., 2010; Pilipenko, Trapezov, Zhuravlev et al., 2015]. Поверхность костей обрабатывали 5%-м раствором гипохлорита натрия для разрушения возможных загрязнений современной ДНК, облучали ультрафиолетом не менее 1 ч, затем удаляли механически поверхностный слой ~1–2 мм и повторно облучали образец ультрафиолетом не менее 1 ч. Из компактного костного вещества высверливали мелкодисперсный порошок.
Для выделения ДНК костный порошок инкубировали в 5М гуанидинизотиоционатном буфере при температуре 65 ºС и постоянном перемешивании. ДНК выделяли методом фенол-хлороформной экстракции с последующим осаждением изопропанолом.
Анализ генетических маркеров. Анализировались четыре системы молекулярно-генетических маркеров: митохондриальная ДНК (последовательность ГВС I и статус соответствующих информативных позиций в кодирующей части мтДНК), фрагмент гена амело-генина (маркер половой принадлежности останков), высоковариабельные аутосомные STR-локусы (универсальные маркеры степени родства индивидов), STR-маркеры Y-хромосомы (филогенетически и филогеографически информативные маркеры, маркеры родства индивидов по мужской линии) (см.: [Пилипенко, Трапезов, Полосьмак, 2015]. МтДНК и STR-локусы Y-хромосомы позволяют реконструировать генетическую историю популяций человека по женской и мужской линиям соответственно. Методы генотипирования каждой из перечисленных систем маркеров приведены ниже.
Амплификацию ГВС I мтДНК проводили двумя разными методами: четырех коротких перекрывающихся фрагментов посредством однораундовой ПЦР [Haak et al., 2005] и одного длинного фрагмента с помощью «вложенной» ПЦР (включала два раунда реакции) [Пилипенко и др., 2008]. Амплификацию информативных фрагментов кодирующей части мтДНК осуществляли с помощью праймеров из работы [Wilde et al., 2014].
По следовательно сти нуклеотидов определяли с использованием набора реактивов ABI Prism BigDye Terminator Cycle Sequencing Ready Reaction Kit (Applied Biosystems, USA). Продукты секвенирующей реакции анализировали на автоматическом капиллярном секвенаторе ABI Prism 3130XL Genetic Analyser (Applied Biosistems, США) в центре коллективного пользования «Геномика» СО РАН (http://sequest. . Филогенетическую и филогеографиче-скую интерпретацию результатов осуществляли методами, описанными нами ранее [Пилипенко, Трапезов, Полосьмак, 2015].
Определение профилей девяти аутосомных STR-локусов и анализ полиморфизма участка гена амело-генина (маркер половой принадлежно сти останков) проводили с использованием коммерческого набора реактивов AmpFlSTR® Profiler® Plus PCR Amplification Kit (Applied Biosystems, США) согласно инструкции производителя. Профили 17 STR-маркеров Y-хромосомы определяли с помощью коммерческого набора реактивов AmpFlSTR® Y-filer® PCR Amplification Kit (Applied Biosystems, США) согласно инструкции производителя. Принадлежность исследованных STR-гаплотипов Y-хромосомы к гапло-группам устанавливали с использованием программы Haplogroup predictor, находящейся в свободном доступе .
Меры против контаминации и верификация результатов. Все работы с древним материалом выполнены в специально оборудованной для палеогенети-ческих исследований лаборатории межинститутского сектора молекулярной палеогенетики (ИЦИГ СО РАН, ИАЭТ СО РАН, г. Новосибирск, Россия). Меры против контаминации и процедуры верификации результатов описаны в нашей статье [Там же]. Их реализация, а также особенности полученных нами результатов свидетельствуют о достоверности экспериментальных данных.
Результаты и обсуждение
Степень сохранности ДНК в останках. Проведенный анализ свидетельствует о различной степени сохранности ДНК в останках взрослого и ребенка из погребального комплекса Бертек-56. Для обоих индивидов получены полноценные данные по структуре мтДНК, включающие последовательность ГВС I и статус соответствующих информативных позиций в кодирующей части мтДНК. Были успешно амплифицированы как короткие фрагменты мтДНК (до 150 п.н., в однораундовой ПЦР), так и длинный участок (методом «вложенной» двухраундовой ПЦР). При этом эффективность амплификации последнего для индивида 1 (ребенок) была ниже, чем для индивида 2 (взрослый). Различие в уровне сохранности ДНК в останках ребенка и взрослого особенно ярко проявилось при анализе ядерных локусов. Для индивида 2 была получена полная информация по всем анализируемым системам ядерных маркеров (полиморфный фрагмент гена аме-логенина, полные аллельные профили STR-локусов аутосом и Y-хромосомы). Это свидетельствует о высокой степени сохранности ядерной ДНК в останках взрослого. Для индивида 1 удалось амплифициро-вать только фрагменты ядерной ДНК длиной не более 15 п.н., включая полиморфный фрагмент гена амелогенина и наиболее короткие фрагменты, содержащие некоторые аутосомные STR-локусы (4 из 9) и STR-локусы Y-хромосомы (5 из 17). Тот факт, что низкая результативность амплификации ДНК из останков ребенка была продемонстрирована с использованием разных систем ядерных маркеров, свидетельствует в пользу именно плохой сохранности ДНК в костном веществе и исключает возможно сть влияния на результат эффективности применяемых вариантов ПЦР. Отметим, что низкая степень сохранности ДНК в останках ребенка коррелирует с особенностями макроскопической сохранности костного материала. Таким образом, получено свидетельство варьирования уровня сохранности ДНК в костях индивидов, захороненных в схожих условиях в пределах одного погребального комплекса. Ранее мы уже сталкивались с подобной ситуацией при анализе останков из погребений пазырыкской культуры Горного Алтая [Пилипенко, Трапезов, Полосьмак, 2015]. Эта разница, на наш взгляд, может объясняться как варьирующи- ми в пределах погребального сооружения локальными условиями среды, так и различиями в условиях, в которых останки находились до их захоронения. Именно последний фактор представляется актуальным в случае с Бертеком-56. Уже отмечалось, что отсутствие ряда элементов скелетов при общем правильном анатомическом порядке костей может свидетельствовать о вторичном характере погребений в этом комплексе.
Половая принадлежность и степень родства индивидов . Определение (или подтверждение) пола погребенных и их родства является одной из актуальных задач палеогенетического исследования парных и коллективных погребений. Для обоих индивидов из кургана Бертек-56 был установлен мужской пол. То, что взрослый погребенный – мужчина, не вызывало сомнений у антропологов. В отношении ребенка по косвенным признакам (крупные размеры черепа и степень развития его рельефа) также было сделано заключение о его возможном мужском поле [Чики-шева, 2012, с. 130]. Полученные палеогенетические результаты (наличие двух аллелей гена амелогенина и амплификация STR-локусов, хоть и неполного их набора) позволяют подтвердить это предположение (табл. 1).
Для выяснения степени родства индивидов необходимы данные по мтДНК и полные (или почти полные) профили STR-локусов аутосом и Y-хромосомы. Как было отмечено выше, в силу плохой сохранности ядерной ДНК ребенка были получены лишь частичные профили STR-локусов (табл. 1, 2). Тем не менее даже имеющиеся данные позволяют с высокой вероятностью исключить прямое родство между погребенными. Другими словами, с учетом мужского пола обоих индивидов исключается вариант «отец и сын». Таким образом, исследованный погребальный комплекс служит еще одной иллюстрацией необходимости проведения анализов степени родства индивидов в парных и коллективных захоронениях вместо неподтвержденных объективными данными реконструкций, строящихся на наиболее вероятных, по мнению иссле-
Таблица 1. Результаты анализа аутосомных STR-локусов и полиморфного участка гена амелогенина в образцах ДНК
|
Локус |
Генотип |
Локус |
Генотип |
||
|
Индивид 1, ребенок |
Индивид 2, взрослый |
Индивид 1, ребенок |
Индивид 2, взрослый |
||
|
D3S1358 |
15/17 |
15/15 |
D18S51 |
Нет данных |
22/22 * |
|
vWA |
17/17 * |
18/18 |
D5S818 |
13/13 * |
10/13 |
|
FGA |
Нет данных |
21/23 |
D13S317 |
Нет данных |
10/12 |
|
D8S1179 |
14/14 * |
14/16 |
D7S820 |
То же |
10/12 |
|
D21S11 |
Нет данных |
30/32.2 |
Amelogenin (пол) |
XY (мужской) |
XY (мужской) |
*Существует вероятность отсутствия сигнала от второго аллеля, который не был амплифицирован из-за деградированного состояния ДНК.
Таблица 2. Результаты анализа профиля STR-локусов Y-хромосомы в образцах ДНК
|
Локус |
Генотип |
Локус |
Генотип |
||
|
Индивид 1, ребенок |
Индивид 2, взрослый |
Индивид 1, ребенок |
Индивид 2, взрослый |
||
|
DYS19 |
Нет данных |
13 |
DYS438 |
Нет данных |
12 |
|
DYS385a/b |
То же |
15/21 |
DYS439 |
То же |
11 |
|
DYS389I |
13 |
14 |
DYS448 |
» |
19 |
|
DYS389II |
Нет данных |
30 |
DYS456 |
15 |
16 |
|
DYS390 |
То же |
24 |
DYS458 |
Нет данных |
16 |
|
DYS391 |
» |
9 |
DYS635 |
То же |
22 |
|
DYS392 |
» |
14 |
YGATAH4 |
13 |
10 |
|
DYS393 |
13 |
14 |
Гаплогруппа |
? |
Q |
|
DYS437 |
14 |
14 |
|||
Таблица 3. Результаты анализа структуры образцов митохондриальной ДНК
|
Номер скелета |
Половозрастные характеристики (по данным физической антропологии) |
Гаплотип ГВС I мтДНК |
Статус маркеров в кодирующей части мтДНК |
Гаплогруппа (подгруппа) мтДНК |
Установленный пол останков |
|
1 |
Ребенок, 8–9 лет |
16145A - 16224C-16311C |
12308G |
K (K1a24a) |
Мужской |
|
2 |
Мужчина, 30–35 лет |
16223T-16287T-16298C-16327T |
13262G |
C |
» |
дователя, вариантах генетических связей погребенных. Следует отметить, что с методологической точки зрения для окончательного подтверждения отсутствия прямого родства требуется получение более полных профилей STR-локусов.
Филогенетическая и филогеографическая интерпретация данных по мтДНК и Y-хромосоме . Для обоих индивидов из кургана Бертек-56 была установлена последовательность ГВС I мтДНК (табл. 3). Дополнительно нами был определен статус позиций в кодирующей части мтДНК, маркирующих установленные по ГВС I гаплогруппы мтДНК. Результаты по ГВС I и кодирующей части мтДНК коррелировали для обоих индивидов. Структура гаплотипов ГВС I позволяет однозначно определить филогенетическое положение исследованных вариантов мтДНК.
У взрослого индивида мтДНК характеризуется гаплотипом 16223T-16287T-16298C-16327T и относится к восточно-евразийской гаплогруппе C. Структура гаплотипа не позволяет определить подгруппу, к которой принадлежит данный вариант. У ребенка мтДНК характеризуется гаплотипом 16145A-16224C-16311C и относится к западно-евразийской гаплогруппе K. Присутствие замены в позиции 16145 свидетельствует о том, что данный вариант принадлежит к подгруппе K1a24a.
Выявленные варианты (как и гаплогруппы, к которым они относятся) отличаются в филогеографиче-ском отношении. Гаплогруппа С широко распростра- нена в современных коренных популяциях восточной части Евразии, как в Сибири (для многих популяций которой она наиболее характерна), так и в более южных областях, включая Центральную Азию, в т.ч. Алтай. Гаплотип 16223T-16287T-16298C-16327T является редким и встречается лишь мозаично. По литературным данным о разнообразии мтДНК в современных коренных популяциях Евразии он обнаружен в Средней Азии – в генофонде киргизов [Comas et al., 1998] и на севере Западной Сибири – в генофонде хантов [Губина, Осипова, Виллемс, 2005]. Такое мозаичное распространение не позволяет связать исследуемый вариант гаплогруппы с населением какой-либо конкретной территории. Можно лишь подчеркнуть, что он, безусловно, относится к восточно-евразийскому кластеру мтДНК и, вероятно, маркирует участие локальной группы населения Южной Сибири (или сопредельных регионов). Это подтверждается и неизменным присутствием различных вариантов гаплогруппы C в автохтонном восточно-евразийском компоненте генофонда мтДНК древних популяций сопредельных районов юга Западной и Восточной Сибири (от Прибайкалья до лесостепной зоны Западной Сибири) начиная как минимум с неолита и во все последующие эпохи [Molodin et al., 2012; Трапезов, Пилипенко, Молодин, 2014]. По нашим неопубликованным данным, гаплогруппа C присутствует также в генофонде окуневского населения Минусинской котловины и каракольского населения Горного Алтая эпохи развитой бронзы (хотя у них выявлены другие варианты гаплогруппы C). Из-за недостатка данных о генофонде мтДНК популяций Саяно-Алтая и сопредельных районов Южной Сибири невозможно выявить ассоциации с известными археологам древними группами населения региона. Очевидно, рассматриваемый вариант относится к автохтонному для этой территории генетическому субстрату, а не происходит от генетически контрастных групп населения других регионов Евразии.
Гаплогруппа K, напротив, характерна для генофонда современных коренных популяций западной части Евразии. При этом варианты, идентичные обнаруженному нами в останках ребенка из Бертека-56, в опубликованных данных были выявлены только в единичных случаях у представителей населения Юго-Западной и Центральной Европы – испанцев [Larruga et al., 2001] и французов [Richards et al., 2000]. Родственные, но не идентичные варианты обнаружены у басков [Corte-Real et al., 1996] и немцев [Pfeiffer et al., 1999], а за пределами Европы – у иранцев [Metspalu et al., 2004]. Подгруппа K1a входит в число гаплогрупп преимущественно ближневосточного происхождения, попавших в Европу в неолите в составе групп ранних животноводов и земледельцев. Распространение в более поздние периоды в восточные области Евразии могло происходить как из Восточной Европы и с прилегающих территорий, так и с юга – из ближневосточно-переднеазиатского региона. Безусловно, на основании одного образца нельзя отдать предпочтение какому-либо конкретному направлению проникновения этого варианта на Алтай. Можно лишь констатировать, что наличие линии гаплогруппы К, несомненно, отражает западный вектор генетических связей исследуемой группы населения. При этом проникновение элементов из западных областей Евразии могло происходить как непосредственно в эпоху развитой бронзы, так и в предшествующие. Гаплогруппа K (другой структурный вариант), в частности, выявлена у представителя афанасьевского населения Саяно-Алтая [Тра-пезов, 2014]. Один из ее вариантов (неродственная подгруппа) обнаружен у носителя андроновской культуры Минусинской котловины [Keyser et al., 2009], хотя гаплогруппа K, по нашим данным, не являлась характерным компонентом генофонда популяций ан-дроновской историко-культурной общности.
Таким образом, исследованные варианты мтДНК филогенетически и филогеографически контрастны. Их присутствие в генофонде одной группы древнего населения свидетельствует о его смешанной структуре, сформированной при взаимодействии групп, имеющих местное (или восточное) и западно-евразийское происхождение. Такая структура в целом характерна для населения эпохи развитой бронзы в обширной зоне контакта западно- и восточно-евразийских популяций, в которую помимо Средней Азии и Западной Сибири входит и Алтайская горная страна.
Наличие полного аллельного профиля Y-хромосомы для взрослого индивида (см. табл. 2) позволило с помощью программы-предиктора определить его принадлежность к гаплогруппе Q, которая в наибольшей степени характерна для современного коренного населения Сибири и Центральной Азии. Ее присутствие (с низкой частотой) в отдельных популяциях западной части Евразии, как правило, связывают с влиянием мигрантов с востока – из Центральной Азии. Таким образом, данный структурный вариант Y-хромосомы также маркирует восточно-евразийский компонент генофонда группы, оставившей памятник Бертек-56. Степень исследованности древнего населения в отношении структуры генофонда Y-хромосомы в настоящий момент очень низкая: известны лишь данные по единичным образцам или очень небольшим сериям. Для носителей андроновской культуры гаплогруппа Q, по-видимому, не была характерна (изучена небольшая серия) [Keyser et al., 2009; Allentoft et al., 2015], как и для носителей афанасьевской (исследованы единичные образцы) [Allentoft et al., 2015]. Из имеющихся для Южной Сибири и Центральной Азии фрагментарных сравнительных материалов эпохи бронзы можно отметить лишь присутствие варианта гаплогруппы Q у представителя карасукского населения Минусинской котловины эпохи поздней бронзы [Ibid.]. В еще более позднее время, в середине I тыс. до н.э. эта гаплогруппа была представлена в генофонде ранних кочевников на юге Внутренней Монголии (Китай) [Zhao et al., 2010]. На Алтае она, по-видимому, сохранялась в раннем железном веке (единичные образцы [Allentoft et al., 2015]). Поиск полностью идентичных аллельных профилей 17 STR-локусов Y-хромосомы в базе данных «Y-Chromosome STR Haplotype Reference Database» выявил присутствие таких вариантов в настоящее время у нескольких представителей населения Северо-Восточного Китая и Южной Кореи (хотя следует отметить малочисленность данных по популяциям Северной Евразии в этой базе, что могло исказить картину).
Таким образом, обнаруженные у взрослого погребенного варианты мтДНК и Y-хромосомы маркируют компонент генофонда, автохтонный для юга Сибири или происходящий из прилегающих районов Восточной Евразии, что согласуется с преобладанием в одонтологическом комплексе индивида признаков восточного ствола [Чикишева, 2012, с. 130]. При этом нами не получено генетических свидетельств эффекта индивидуальной метисации (т.е. происхождения индивида от генетически резко контрастных родителей), о чем было высказано предположение на основании антропологических данных. В то же время присутствие в генофонде группы филогенетически и филогеографически контрастных вариантов мтДНК указывает на его смешанную структуру. Очевидно, мозаичность одонтологических и краниометрических признаков, отмеченная антропологами для индивидов из Бертека-56, не является следствием индивидуальной метисации, а отражает механизм формирования популяции в целом в результате смешения генетически (и антропологически) контрастных групп населения. При этом «восточные» элементы, вероятно, могут быть связаны с носителями каракольской и/или окуневской культур Южной Сибири (что согласуется с антропологическими данными), а «западные» пока не удается ассоциировать с конкретными группами древнего населения западного происхождения.
Следует отметить своеобразие в пределах Алтайской горной страны керамики, найденной на памятнике [Молодин и др., 2004]. Интересно, что в регионе морфологически очень близкая посуда была обнаружена при раскопках ритуального комплекса на р. Ку-черле [Молодин, Ефремова, 2001]. При этом культуросодержащий слой эпохи бронзы на памятнике не выявлен [Молодин, Ефремова, 2010]. В контексте данной работы важно отметить, что прямые параллели конструкциям погребальных сооружений и морфологически близкая керамика обнаружены в Акбацком могильнике эпохи бронзы на п-ове Мангышлак [Баландина, Астафьев, 2000]. С этим регионом мог быть связан миграционный импульс, оставивший свой след на плоскогорье Укок. Следует иметь в виду, что погребальный обряд комплекса Бертек-56 отличается от такового как соседствующей на севере каракольской культуры [Кубарев, 1988; Молодин, 2006], который по всем канонам ближе к окуневскому, так и выявленной на юге чемурчекской [Древнейшие европейцы…, 2014]. Вероятно, мы имеем дело с особым этнокультурным образованием, имеющим автохтонную основу и в то же время принесенный с запада своеобразный колорит. Все это выявляется на археологическом и антропологическом материале, а теперь еще и подтверждается данными палеогенетики.
Продолжение реконструкции сложных этногенетических процессов, протекавших на территории Южной Сибири в различные периоды эпохи бронзы и в последующее время, будет в значительной степени зависеть от накопления новых данных по структуре генофонда многочисленных групп древнего населения, как по однородительским (мтДНК, Y-хромосома), так и по другим генетическим маркерам, и их корректной интерпретации с учетом археологического и антропологического контекста исследуемых материалов. Эта задача, в частности, реализуется нашим междисциплинарным коллективом в рамках нескольких научных проектов.
Список литературы Молекулярно-генетический анализ останков людей из погребального комплекса эпохи бронзы Бертек-56 (II тыс. до н.э., Республика Алтай, Россия)
- Баландина Г.В., Астафьева А.Е. Курганы эпохи поздней бронзы с полуострова Мангышлак//Проблемы изучения энеолита и бронзового века Южного Урала. -Орск: Ин-т евразийских исследований, Ин-т степи УрО РАН, 2000. -С. 149-166.
- Губина М. А., Осипова Л.П., Виллемс Р. Анализ материнского генофонда по полиморфизму митохондриальной ДНК в популяциях хантов и коми Шурышкарского района ЯНАО//Коренное население Шурышкарского района Ямало-Ненецкого автономного округа: демографические, генетические и медицинские аспекты. -Новосибирск: Artavenue, 2005. -С. 105-117.
- Древнейшие европейцы в сердце Азии: чемурчекский культурный феномен. -СПб.: Изд-во Санкт-Петербург. музея-института семьи Рерихов, 2014. -Ч. 1: Результаты исследований в Восточном Казахстане, на севере и юге Монгольского Алтая/науч. ред. А.А. Ковалев. -416 с.
- Кубарев В.Д. Древние росписи Каракола. -Новосибирск: Наука, 1988. -174 с.
- Молодин В.И. Основные итоги археологических исследований западносибирского отряда Северо-Азиатской комплексной экспедиции на плоскогорье Укок летом 1992 года//ALTAIKA. -1993. -Вып. 2. -С. 17-20.
